Topographic Factors and Tree Heights of Aged Cryptomeria japonica Plantations in the Boso Peninsula, Japan

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Survey
2.3. Topographic Analysis
2.4. Statistical Method
3. Results
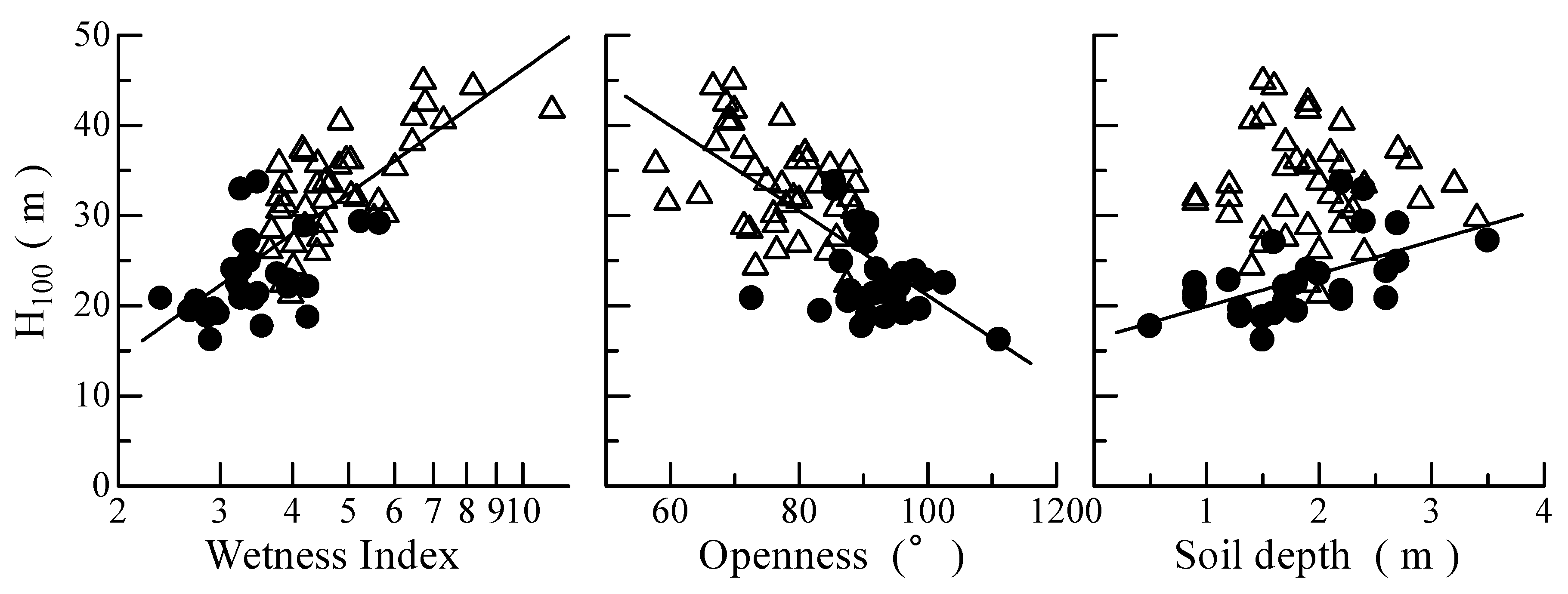
3.1. Topographical Characteristics and Tree Heights
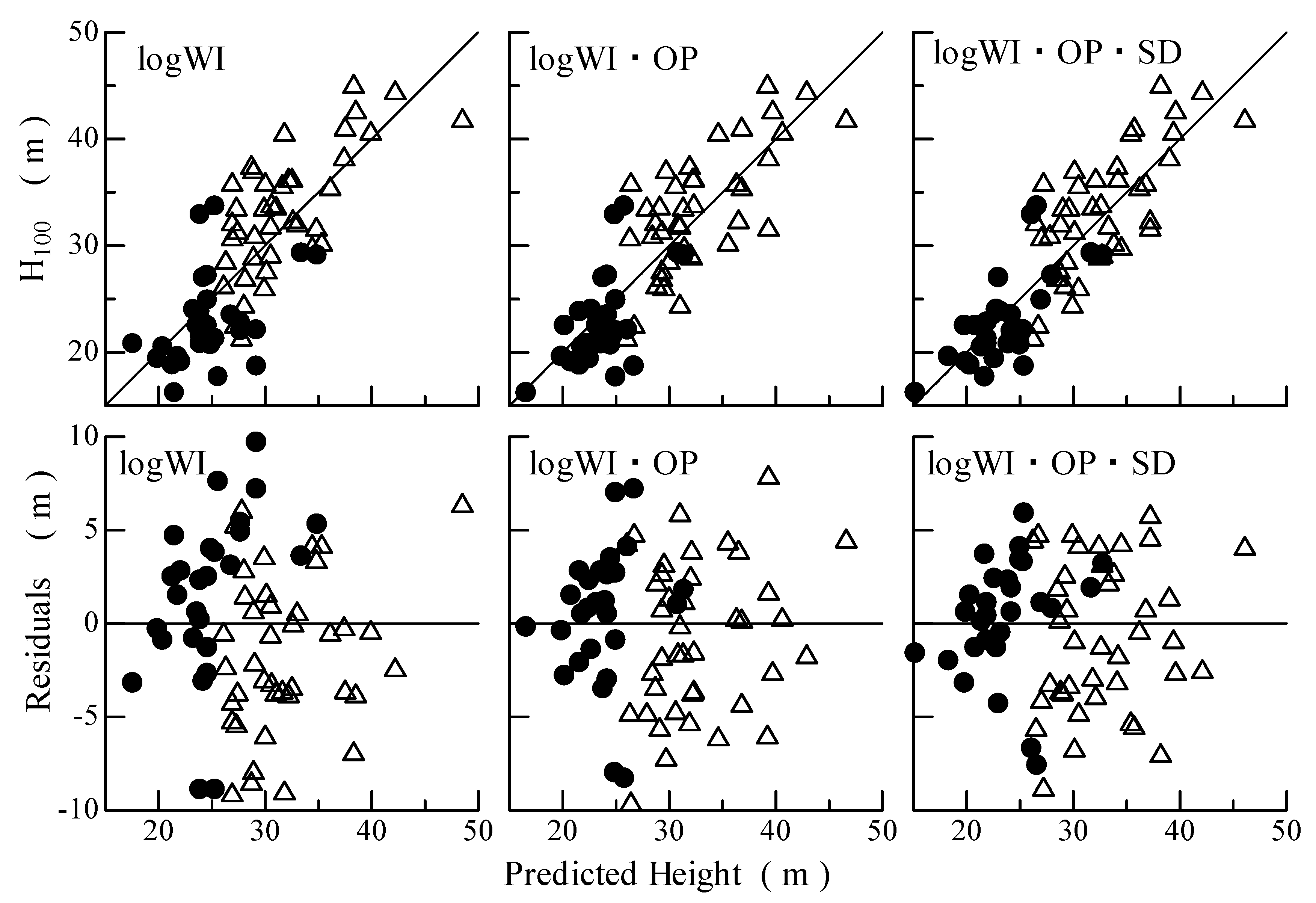
3.2. Prediction of Tree Height from Topographical Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Plot | Age | Elevation | Direction | Slope | Wetness Index | Openness | Soil Depth (m) | Height (m) |
|---|---|---|---|---|---|---|---|---|
| (Year) | (M) | (°) | (°) | (°) | ||||
| R1 | 94 | 320 | 98 | 19 | 3.37 | 93.8 | 0.9 | 22.1 |
| R2 | 99 | 226 | 55 | 21 | 3.26 | 98.1 | 2.6 | 23.7 |
| R3 | 99 | 273 | 247 | 8 | 4.26 | 95.6 | 1.7 | 22.0 |
| R4 | 100 | 321 | 205 | 43 | 2.37 | 72.7 | 0.9 | 20.8 |
| R5 | 100 | 312 | 129 | 11 | 3.94 | 99.6 | 1.2 | 22.8 |
| R6 | 100 | 251 | 21 | 21 | 3.26 | 85.5 | 2.4 | 32.9 |
| R7 | 100 | 305 | 233 | 30 | 2.85 | 90.7 | 1.3 | 18.8 |
| R8 | 100 | 302 | 201 | 21 | 3.26 | 88.0 | 2.2 | 21.6 |
| R9 | 100 | 217 | 161 | 19 | 3.37 | 89.7 | 3.5 | 27.2 |
| R10 | 102 | 304 | 74 | 23 | 3.16 | 92.1 | 1.9 | 24.1 |
| R11 | 103 | 223 | 72 | 17 | 3.49 | 91.7 | 0.9 | 21.5 |
| R12 | 103 | 308 | 59 | 27 | 2.98 | 96.4 | 1.6 | 19.3 |
| R13 | 103 | 316 | 233 | 13 | 3.77 | 96.2 | 2.0 | 23.7 |
| R14 | 105 | 196 | 210 | 20 | 3.31 | 90.4 | 1.6 | 27.3 |
| R15 | 105 | 202 | 265 | 11 | 3.94 | 95.5 | 1.7 | 22.3 |
| R16 | 105 | 205 | 17 | 17 | 3.49 | 85.5 | 2.2 | 34.2 |
| R17 | 105 | 239 | 297 | 18 | 3.43 | 89.7 | 2.2 | 21.0 |
| R18 | 106 | 197 | 279 | 3 | 5.25 | 88.9 | 2.4 | 29.8 |
| R19 | 106 | 210 | 350 | 2 | 5.66 | 90.7 | 2.7 | 29.6 |
| R20 | 107 | 227 | 276 | 16 | 3.55 | 89.8 | 0.5 | 18.0 |
| R21 | 108 | 280 | 259 | 22 | 3.21 | 102.6 | 1.8 | 23.0 |
| R22 | 113 | 340 | 276 | 28 | 2.93 | 98.8 | 1.3 | 20.3 |
| R23 | 114 | 340 | 162 | 29 | 2.89 | 111.1 | 1.5 | 16.8 |
| R24 | 114 | 302 | 263 | 8 | 4.26 | 93.4 | 1.5 | 19.5 |
| R25 | 114 | 323 | 284 | 33 | 2.73 | 87.7 | 1.7 | 21.3 |
| R26 | 115 | 314 | 297 | 35 | 2.66 | 83.3 | 1.8 | 20.2 |
| R27 | 115 | 217 | 87 | 21 | 3.26 | 94.9 | 2.6 | 21.6 |
| R28 | 115 | 154 | 199 | 19 | 3.37 | 86.6 | 2.7 | 25.9 |
| M1 | 93 | 270 | 132 | 42 | 4.20 | 71.4 | 1.9 | 28.3 |
| M2 | 94 | 308 | 132 | 23 | 3.85 | 87.4 | 1.9 | 22.0 |
| M3 | 94 | 292 | 161 | 28 | 4.54 | 80.0 | 2.9 | 31.2 |
| M4 | 94 | 299 | 124 | 32 | 3.87 | 83.0 | 2.4 | 32.9 |
| M5 | 96 | 255 | 49 | 36 | 4.41 | 77.3 | 1.2 | 33.0 |
| M6 | 96 | 232 | 73 | 27 | 6.44 | 67.1 | 1.7 | 37.7 |
| M7 | 99 | 270 | 310 | 18 | 4.81 | 84.8 | 1.9 | 35.4 |
| M8 | 99 | 186 | 39 | 18 | 5.04 | 81.2 | 2.8 | 36.0 |
| M9 | 99 | 233 | 205 | 42 | 4.02 | 79.9 | 1.5 | 26.7 |
| M10 | 99 | 245 | 208 | 16 | 7.29 | 68.8 | 1.4 | 40.4 |
| M11 | 99 | 243 | 298 | 29 | 6.78 | 68.6 | 1.9 | 42.4 |
| M12 | 100 | 239 | 345 | 32 | 4.16 | 71.4 | 2.7 | 37.3 |
| M13 | 100 | 262 | 80 | 31 | 4.20 | 80.9 | 2.1 | 36.9 |
| M14 | 100 | 229 | 108 | 46 | 3.88 | 78.0 | 2.2 | 31.2 |
| M15 | 100 | 250 | 350 | 23 | 4.95 | 79.7 | 1.8 | 36.1 |
| M16 | 100 | 298 | 278 | 44 | 5.63 | 59.5 | 0.9 | 31.5 |
| M17 | 102 | 272 | 118 | 40 | 4.56 | 75.0 | 2.0 | 33.9 |
| M18 | 103 | 276 | 247 | 21 | 6.48 | 77.3 | 1.5 | 41.2 |
| M19 | 103 | 296 | 227 | 34 | 3.79 | 79.1 | 0.9 | 32.2 |
| M20 | 104 | 230 | 97 | 40 | 4.42 | 57.7 | 1.9 | 36.1 |
| M21 | 104 | 198 | 232 | 16 | 6.73 | 69.8 | 1.5 | 45.3 |
| M22 | 104 | 205 | 283 | 34 | 3.79 | 87.8 | 2.2 | 36.1 |
| M23 | 104 | 265 | 47 | 37 | 3.68 | 72.3 | 1.5 | 28.7 |
| M24 | 105 | 206 | 21 | 29 | 4.84 | 69.5 | 2.2 | 40.8 |
| M25 | 105 | 293 | 299 | 30 | 5.05 | 64.5 | 2.1 | 32.7 |
| M26 | 105 | 196 | 185 | 3 | 8.20 | 66.6 | 1.6 | 44.7 |
| M27 | 105 | 193 | 283 | 3 | 11.23 | 69.9 | 1.9 | 42.1 |
| M28 | 105 | 213 | 281 | 24 | 4.21 | 85.8 | 1.7 | 31.2 |
| M29 | 107 | 199 | 336 | 9 | 5.53 | 89.5 | 3.4 | 30.3 |
| M30 | 107 | 238 | 342 | 16 | 5.16 | 87.9 | 1.2 | 32.5 |
| M31 | 107 | 197 | 231 | 34 | 3.79 | 88.5 | 2.3 | 31.2 |
| M32 | 108 | 267 | 188 | 28 | 4.54 | 76.3 | 2.2 | 29.6 |
| M33 | 108 | 255 | 172 | 20 | 4.41 | 84.4 | 2.4 | 26.4 |
| M34 | 109 | 193 | 56 | 38 | 3.65 | 76.5 | 2.0 | 26.7 |
| M35 | 111 | 341 | 324 | 37 | 3.97 | 91.9 | 2.0 | 21.8 |
| M36 | 114 | 183 | 6 | 16 | 4.65 | 88.8 | 3.2 | 34.8 |
| M37 | 114 | 214 | 274 | 36 | 4.01 | 73.2 | 1.4 | 25.2 |
| M38 | 114 | 218 | 352 | 13 | 4.46 | 85.8 | 1.7 | 28.6 |
| M39 | 114 | 295 | 128 | 23 | 5.80 | 76.0 | 1.2 | 31.2 |
| M40 | 115 | 210 | 233 | 25 | 6.01 | 73.1 | 1.7 | 36.7 |
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risk for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Kirilenko, A.P.; Sedjo, R.A. Climate change impacts on forestry. Proc. Natl. Acad. Sci. USA 2007, 104, 19697–19702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skovsgaard, J.P.; Vanclay, J.K. Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry 2008, 81, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Bontemps, J.D.; Bouriaud, O. Predictive approaches to forest site productivity: Recent trends, challenges and future perspectives. Forestry 2014, 87, 109–128. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, M.A.; Dieguez-Aranda, U. Exploring the use of learning techniques for relating the site index of radiata pine stands with climate, soil and physiography. For. Ecol. Manag. 2020, 458, 117803. [Google Scholar] [CrossRef]
- Mitsuda, Y.; Ito, S.; Sakamoto, S. Predicting the site index of sugi plantations from GIS-derived environmental factors in Miyazaki Prefecture. J. For. Res. 2007, 12, 177–186. [Google Scholar] [CrossRef]
- Hunter, I.R.; Gibson, A.R. Predicting Pinus radiata site index from environmental variables. N. Z. J. For. Sci. 1984, 14, 53–64. [Google Scholar]
- Bjelanovic, I.; Comeau, P.G.; White, B. High resolution site index prediction in boreal forests using topographic and wet areas mapping attributes. Forests 2018, 9, 113. [Google Scholar] [CrossRef] [Green Version]
- Socha, J.; Tyminska-Czabanska, L.; Grabska, E.; Orzel, S. Site index models for forest-forming tree species in Poland. Forests 2020, 11, 301. [Google Scholar] [CrossRef] [Green Version]
- Yoder, B.J.; Ryan, M.G.; Waring, R.H.; Schoettle, A.W.; Kaufmann, M.R. Evidence of reduced photosynthetic rates in old trees. For. Sci. 1994, 40, 513–527. [Google Scholar]
- Ryan, M.G.; Yoder, B.J. Hydraulic limits to tree height and tree growth. BioScience 1997, 47, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Ryan, M.G.; Bond, B.J.; Law, B.E.; Hubbard, R.B.; Woodruff, D.V.; Cienciala, E.; Kucera, J. Transpiration and whole-tree conductance in ponderosa pine trees of different heights. Oecologia 2000, 124, 553–560. [Google Scholar] [CrossRef]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Shibata, E. Why do trees decline or dieback after a strong wind? Water status of Hinoki cypress standing after a typhoon. Tree Physiol. 2004, 24, 701–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locatelli, T.; Tarantola, S.; Gardiner, B.; Patenaude, G. Variance-based sensitivity analysis of a wind risk model-Model behavior and lessons for forest modelling. Environ. Model. Softw. 2017, 87, 84–109. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, M.; Mashimo, Y.; Kawabata, K. Estimation method of site index by quantification. Bull. FFPRI. 1965, 176, 1–54, (In Japanese with English summary). [Google Scholar]
- Nakamura, T. Relation between the soil morphological properties and the growing processes of Cryptomeria stands in the University Forest in Tiba Prefecture. Bull. Univ. Tokyo For. 1943, 32, 1–70, (In Japanese with English Summary). [Google Scholar]
- Mashimo, Y. Studies on the physical properties of forest soil and their relation to the growth of sugi (Cryptomeria japonica) and hinoki (Chamaecyparis obtusa). For. Soils Jpn. 1960, 11, 1–182, (In Japanese with English Summary). [Google Scholar]
- Tange, T.; Matsumoto, Y.; Mashimo, Y.; Sakura, T. Course of height growth of Cryptomeria japonica trees planted on a slope. Comparison among the near ridge, the mid-slope and the near bottom in the Tokyo University Forest in Chiba. Bull. Univ. Tokyo For. 1989, 81, 39–51, (In Japanese with English Summary). [Google Scholar]
- Karizumi, N. Illustration of Tree Roots; Seibunndo Sinkosha: Tokyo, Japan, 1979; pp. 592–596. (In Japanese) [Google Scholar]
- Tange, T.; Someya, M. Soil properties of Cryptomeria japonica plantations with different height growth courses at the Experimental Station at Tanashi, The University of Tokyo. Misc. Inf. Univ. Tokyo For. 2013, 53, 1–7. (In Japanese) [Google Scholar]
- Tange, T.; Someya, M.; Norisada, M.; Masumori, M. Photosynthetic limitation of similar-height Cryptomeria japonica trees growing at different rates. Photosynthetica 2013, 51, 158–160. [Google Scholar] [CrossRef]
- Sakata, M. Evaluation of possible causes for the decline of Japanese cedar (Cryptomeria japonica) based on elemental composition and δ13C of needles. Environ. Sci. Technol. 1996, 30, 2376–2381. [Google Scholar] [CrossRef]
- Zushi, K. Spatial distribution of soil carbon and nitrogen storage and forest productivity in a watershed planted to Japanese cedar (Cryptomeria japonica D. Don). J. For. Res. 2006, 11, 351–358. [Google Scholar] [CrossRef]
- Forestry Agency. Annual Report on Forest and Forestry in Japan, Fiscal Year 2018. Available online: https://www.maff.go.jp/e/data/publish/attach/pdf/index-170.pdf (accessed on 10 May 2020).
- The University of Tokyo Forests. Meteorological Data. 2020. Available online: http://www.uf.a.u-tokyo.ac.jp/research_division/data/kishou/index_english.html (accessed on 1 March 2020).
- Maita, E.; Suzuki, M.; Ohta, T. Change in the annual discharge after clear-cutting a 70-year-old mixed plantation of sugi and hinoki in a Tertiary formation area. J. Jpn. For. Soc. 2005, 87, 124–132, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- IIjima, A.; Ikeya, N. Geology of the Tokyo University Forest in Chiba. Misc. Inf. Univ. Tokyo For. 1976, 20, 1–38, (In Japanese with English Summary). [Google Scholar]
- Forest Soil Division. Classification of forest soils in Japan. Bull. FFPRI 1976, 280, 1–28, (In Japanese with English Summary). [Google Scholar]
- Tange, T. Ecophysiological study on the growth of Cryptomeria japonica planted trees. Bull. Univ. Tokyo For. 1995, 93, 1–139, (In Japanese with English Summary). [Google Scholar]
- Shiraishi, N. Study on the growth prediction of even-aged stands. Bull. Univ. Tokyo For. 1986, 75, 199–256, (In Japanese with English Summary). [Google Scholar]
- Suzuki, M.; Shiraishi, N.; Suzuki, M.; Tatsuhara, S.; Yamamoto, H. Studies on the growth of old sugi (Cryptomeria japonica) stand (I): The development process of the old sugi plantations in the Kiyosumi area. Bull. Univ. Tokyo For. 2003, 109, 27–45, (In Japanese with English Summary). [Google Scholar]
- Geospatial Information Authority of Japan. Digital Elevation Model. 2020. Available online: https://fgd.gsi.go.jp/download/ref_dem.html (accessed on 1 March 2020).
- Beven, K.J.; Kirkby, M.J. A physically based, variable contributing area model of basin hydrogy. Hydrol. Sci. 1979, 24, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; McCarty, G.W.; Lang, M.; Ducey, T.; Hunt, P.; Miller, J. Topographic and physicochemical control on soil denitrification in prior converted croplands located on the Delmarva Peninsula, USA. Geoderma 2018, 309, 41–49. [Google Scholar] [CrossRef]
- Yokoyama, R.; Shirasawa, M.; Kikuchi, Y. Representation of topographic feature by openness. J. Jpn. Soc. Photogramm. Remote Sens. 1999, 38, 26–34, (In Japanese with English Summary). [Google Scholar]
- Prime, O.D.A.; Echigo, A.; Yokoyama, R.; Yoshida, T. Supervised landform classification of Northeast Honshu from DEM-derived thematic maps. Geomorphology 2006, 78, 373–386. [Google Scholar] [CrossRef]
- Quinn, P.F.; Beven, K.J. Spatial and temporal predictions of soil moisture dynamics, runoff, variable source areas and evapotranspiration for Plynlimon, Mid-Wales. Hydrol. Process. 1993, 7, 425–448. [Google Scholar] [CrossRef]
- Quinn, P.F.; Beven, K.J.; Lamb, R. The ln(a/tanβ) index: How to calculate and how to use it within the TOPMODEL framework. Hydrol. Process. 1995, 9, 161–182. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Ito, T.; Ueda, M.; Miyauchi, H. Variation of the nutrient content in sugi (Cryptomeria japonica) needles according to tree age, site quality and forest site unit. J. Jpn. For. Soc. 1972, 54, 74–79, (In Japanese with English Summary). [Google Scholar]
- Meredieu, C.; Arrouays, D.; Goulard, M.; Auclair, D. Short range soil variability and its effect on red oak growth (Quercus rubra L.). Soil Sci. 1996, 161, 29–38. [Google Scholar] [CrossRef]
- Zhang, W.; Montgomery, D.R. Digital elevation model grid size, landscape representation and hydrologic simulations. Water Resour. Res. 1994, 30, 1019–1028. [Google Scholar] [CrossRef]
- Socha, J.; Pierzchalski, M.; Baiazy, R.; Clesieiski, M. Modelling top height growth and site index using repeated laser scanning data. For. Ecol. Manag. 2017, 406, 307–317. [Google Scholar] [CrossRef]
- Noordermeer, L.; Gobakken, T.; Naessert, E.; Bollandsas, O.M. Predicting and mapping site index in operational forest inventories using bitemporal airborne laser scanner data. For. Ecol. Manag. 2020, 457, 117768. [Google Scholar] [CrossRef]
- Sasaki, H.; Murata, A.; Hanafusa, M.; Oh’izumi, M.; Kurihara, K. Projection of future climate in a non-hydrostatic regional climate model nested within an atmospheric general circulation model. SOLA 2012, 8, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Kusumoto, S.; Mizuta, R. Future changes in the baiu rain band projected by a 20-km mesh global atmospheric model: Sea surface temperature dependence. SOLA 2008, 4, 85–88. [Google Scholar]
- Japan Meteorology Agency. Archives of Weather Observations. Available online: http://www.data.jma.go.jp/obd/stats/etrn/index.php (accessed on 4 July 2020).


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tange, T.; Ge, F. Topographic Factors and Tree Heights of Aged Cryptomeria japonica Plantations in the Boso Peninsula, Japan. Forests 2020, 11, 771. https://doi.org/10.3390/f11070771
Tange T, Ge F. Topographic Factors and Tree Heights of Aged Cryptomeria japonica Plantations in the Boso Peninsula, Japan. Forests. 2020; 11(7):771. https://doi.org/10.3390/f11070771
Chicago/Turabian StyleTange, Takeshi, and Feng Ge. 2020. "Topographic Factors and Tree Heights of Aged Cryptomeria japonica Plantations in the Boso Peninsula, Japan" Forests 11, no. 7: 771. https://doi.org/10.3390/f11070771