First Age-Estimation Model for Dracaena ombet and Dracaena draco subsp. caboverdeana

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
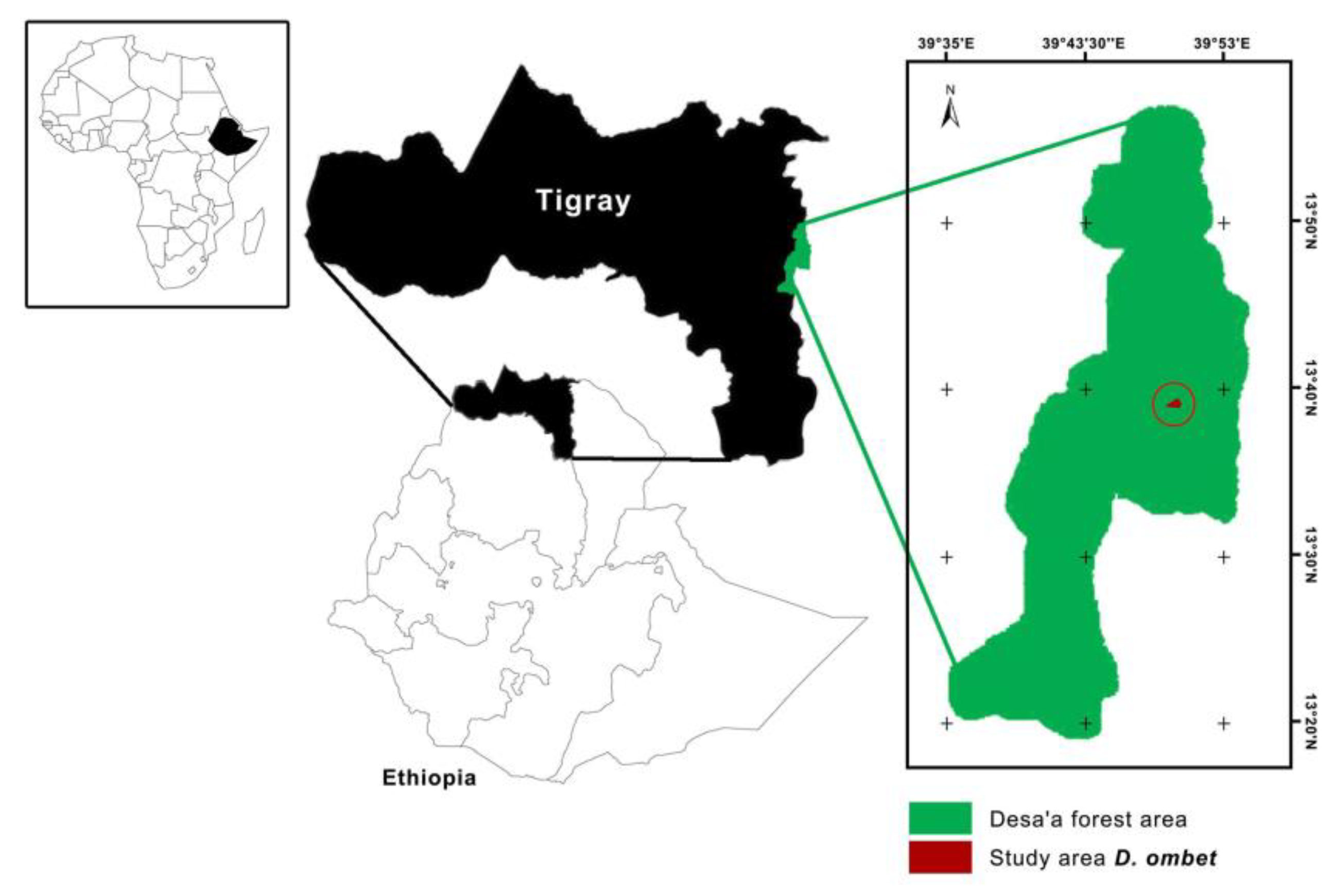
2.1. Study Area
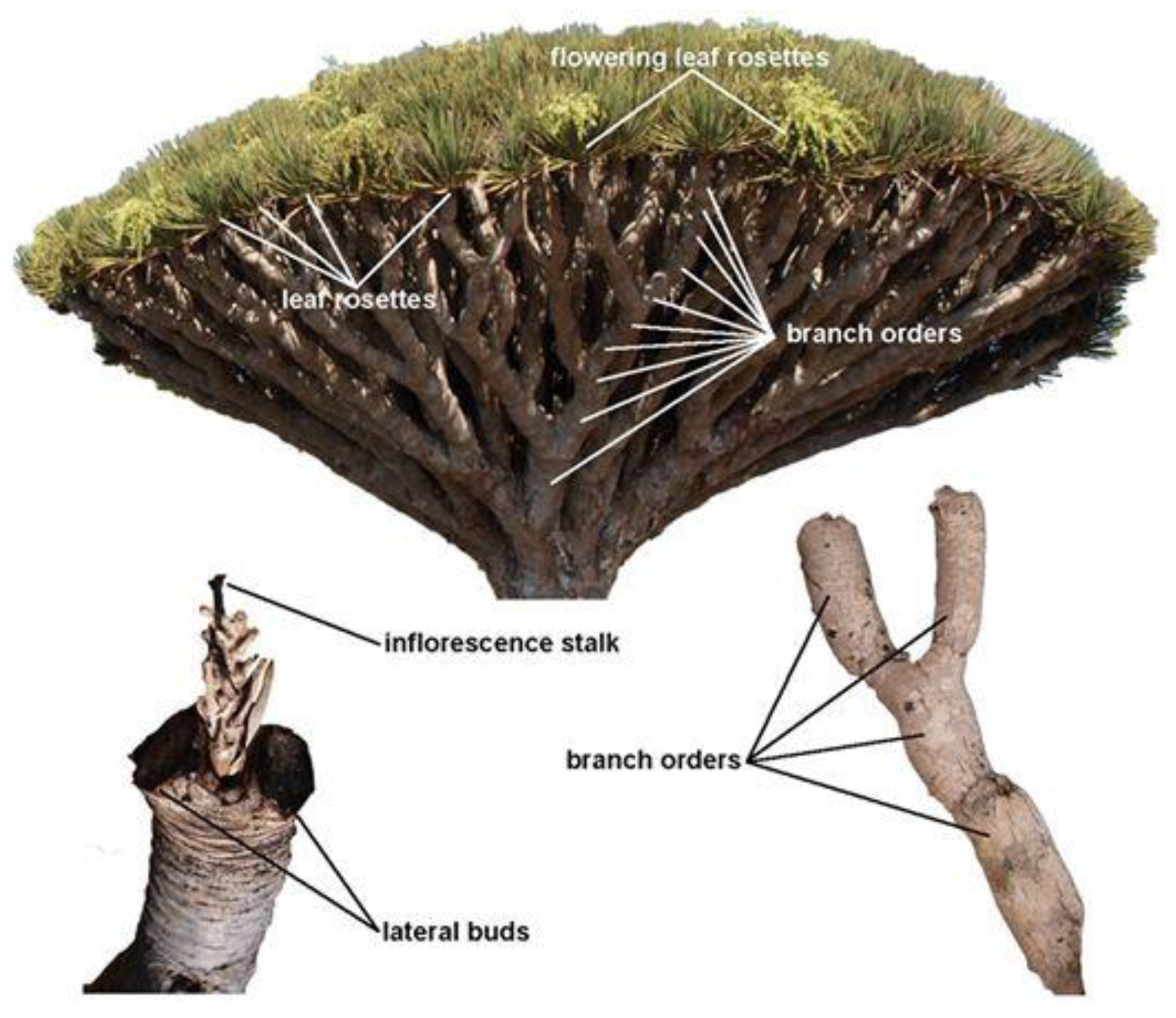
2.2. Data Ccollection
2.3. Crown Age Calculation
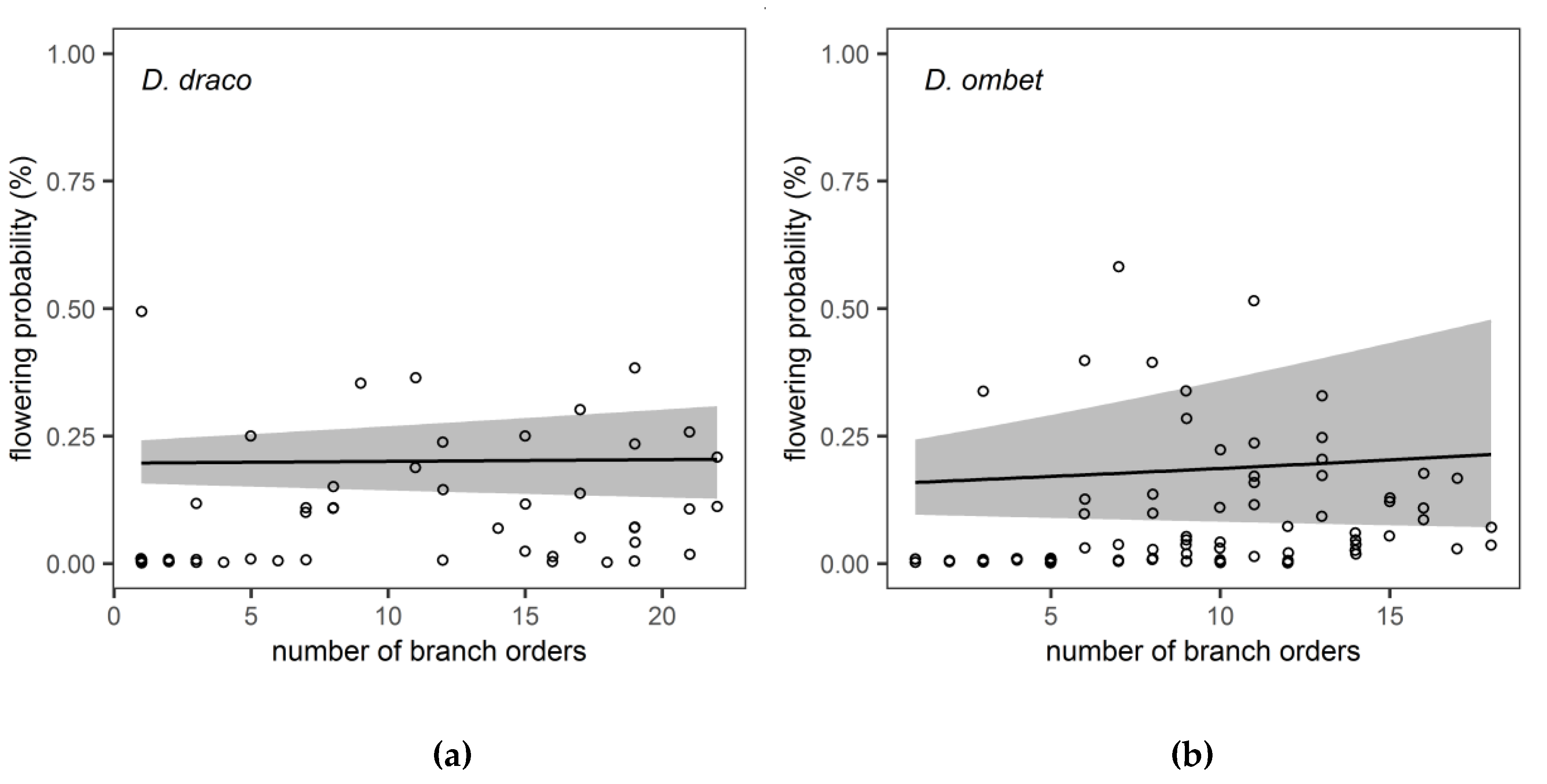
3. Results
4. Discussion
4.1. Comparisons with Age Estimations of Other Dragon Tree Species
4.2. Effects of Environmental Conditions
4.3. Suggestions for Future Research
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.-L.; Morden, C.W. Phylogenetic relationships among dracaenoid genera (Asparagaceae: Nolinoideae) inferred from chloroplast DNA loci. Syst. Bot. 2014, 39, 90–104. [Google Scholar] [CrossRef]
- Bos, J. Dracaena. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 3, pp. 238–241. [Google Scholar]
- Klimko, M.; Nowińska, R.; Wilkin, P.; Wiland-Szymańska, J. Comparative leaf micromorphology and anatomy of the dragon tree group of Dracaena (Asparagaceae) and their taxonomic implications. Plant Syst. Evol. 2018, 304, 1041–1055. [Google Scholar] [CrossRef] [Green Version]
- Adolt, R.; Pavliš, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari Balf. f. of Socotra Island suggest essential elements for a conservation strategy. Biológia 2018. [Google Scholar] [CrossRef]
- Hubálková, I.; Maděra, P.; Volařík, D. Growth dynamics of Dracaena cinnabari under controlled conditions as the most effective way to protect endangered. Saudi J. Biol. Sci. 2017, 24, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Marrero, A.; Almeida, R.S.; Gonzalez-Martin, M. A new species of the wild dragon tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Habrová, H.; Maděra, P. Ecology of Dragon’s blood tree (Dracaena cinnabari) communities on the Socotra Island. In Proceedings of the Conference Evaluation of State and Development of Forest Geobiocoenosis, Brno, Czech Republic, 15–16 October 2004; Polehla, P., Ed.; Faculty of Forestry and Wood Technology Mendel University: Brno, Czech Republic, 2004; pp. 120–126. [Google Scholar]
- Kamel, M.; Ghazaly, U.M.; Callmander, M.W. Conservation status of the endangered Nubian dragon tree dracaena ombet in Gebel Elba national park, Egypt. Oryx 2015, 49, 704–709. [Google Scholar] [CrossRef] [Green Version]
- WCSP. World Checklist of Selected Plant Families. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://wcsp.science.kew.org/ (accessed on 29 November 2019).
- Elnoby, S.K.; Moustafa, A.A.; Mansour, S.R. Impact of climate change on the endangered Nubian dragon tree (Dracaena ombet) in the South Eastern of Egypt. Catrina 2017, 16, 23–28. [Google Scholar]
- Aynekulu, E.; Aerts, R.; Moonen, P.; Denich, M.; Gebrehiwot, K.; Vagen, T.G.; Wolde, M.; Boehmer, H.J. Altitudinal variation and conservation priorities of Vegetation along the great rift valley escarpment, Northern Ethiopia. Biodivers. Conserv. 2012, 21, 2691–2707. [Google Scholar] [CrossRef]
- El Azzouni, M. Conserving Dracaena ombet Egypt’s dragon tree. Plant Talk 2003, 34, 38–39. [Google Scholar]
- Nadezhdina, N.; Plichta, R.; Nadezhdin, V.; Gebauer, R.; Jupa, R.; Habrová, H.; Maděra, P. A comparative structural and functional study of leaf traits and sap flow in Dracaena cinnabari and Dracaena draco seedlings. Funct. Plant Biol. 2015, 42, 1092–1105. [Google Scholar] [CrossRef]
- Marrero, A.; Almeida, S.R. A new subspecies, Dracaena draco (L.) L. subsp. caboverdeana Marrero Rodr. & R. Almeida (Dracaenaceae) from Cape Verde Island. Int. J. Geobot. Res. 2012, 2, 35–40. [Google Scholar] [CrossRef]
- Marerro, A. Dragos. Rincones Atlántico 2010, 6/7, 130–140. [Google Scholar]
- Hubálková, I. Prediction of dragon’s blood tree (Dracaena cinnabari Balf.) stand sample density on Soqotra Island. J. Landsc. Ecol. 2011, 4, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.; Mies, B.A. Vegetation Ecology of Socotra. Plant and Vegetation 7, 1st ed.; Springer: Heidelberg, Germany, 2012; p. 379. [Google Scholar]
- Zheng, D.J.; Li, H.W.; Yun, Y.; Zhang, L.; Zhang, Z.L. Investigation on the habitat and natural regeneration capacity of Dracaena cambodiana. J. Trop. Subtrop. Bot. 2010, 18, 627–632. [Google Scholar]
- Habrová, H.; Pavliš, J. Dynamic response of woody vegetation on fencing protection in semi-arid areas; Case study: Pilot exclosure on the Firmihin Plateau, Socotra island. Saudi J. Biol. Sci. 2017, 24, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Čermák, Z.; Pavliš, J. Dragon’s blood tree—Threatened by overmaturity, not by extinction: Dynamics of a Dracaena cinnabari woodland in the mountains of Soqotra. Biol. Conserv. 2009, 142, 772–778. [Google Scholar] [CrossRef]
- Robiansyah, I.; Hajar, A.S. Predicting current and future distribution of endangered tree Dracaena ombet Kotschy and Peyr. Under climate change. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 87, 225–232. [Google Scholar] [CrossRef]
- Ghazali, U. Effect of climatic change on the endangered Dracaena ombet tree in the Gabel Elba protected area in Egypt. Mt. Forum. Bull. 2008, 8, 11–12. [Google Scholar]
- Ghazali, U. The Globally Endangered Dracaena Ombet Monitoring and Assessment Project in Gabel Elba Protected Area, Egypt. Final Report, Conservation Leadership Programme. 2008. Available online: http://www.conservationleadershipprogramme.org/ (accessed on 5 December 2013).
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable land use management needed to conserve the dragon´s blood tree of Socotra Island, a vulnerable endemic umbrella species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef] [Green Version]
- IUCN Red List of Threatened Species Version 2017.2. Available online: htttp://www.iucnredlist.org (accessed on 25 December 2019).
- Pütter, A. Altersbestimmung an Drachenbäumen von Tenerife. Sitz. Heidelb. Akad. Wissenschäften. Math.Nat. Kl. 1925, 12, 12–18. [Google Scholar]
- Adolt, R.; Habrová, H.; Maděra, P. Crown age estimation of a monocotyledonous tree species Dracaena cinnabari using logistic regression. Trees Struct. Funct. 2012, 26, 1287–1298. [Google Scholar] [CrossRef]
- Gebreegziabher, Z. Dessa’a Protected Area: An Assessment of Human Impact, Evolutionary Pattern and Options for Sustainable Management; Research Report; Tigray Region Bureau of Agriculture and Natural Resources: Mekelle, Ethiopia, 1999; p. 35. [Google Scholar]
- Alemayehu, W.; Demel, T.; Powell, N. Church forests in North Gondar Administrative Zone, northern Ethiopia. For. Trees Livelihoods 2005, 15, 349–374. [Google Scholar] [CrossRef]
- Asfawossen, A. The rock-hewn churches of Tigrai, northern Ethiopia: A geological perspective. Geoarcheology 2002, 17, 649–663. [Google Scholar] [CrossRef]
- Friis, I. Forests and Forest Trees of Northeast Tropical Africa: Their Natural Habitats and Distribution Patterns in Ethiopia, Djibouti and Somalia; HMSO: Middlesex, UK, 1992; p. 396.
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- White, F. The Vegetation of Africa, 1st ed.; UNESCO: Paris, France, 1983; p. 365. [Google Scholar]
- Duarte, M.C.; Romeiras, M.M. Cape Verde Islands. In Encyclopedia of Islands, 1st ed.; Gillespie, R.G., Clague, D.A., Eds.; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 2009; pp. 143–148. [Google Scholar]
- Duarte, M.C.; Romeiras, M.M.; Santos-Guerra, A.; Nepi, C.; Francisco-Ortega, J.; Joseph, D. Hooker’s 1839 Cabo Verde collections. Curtis’ Bot. Mag. 2017, 34, 146–168. [Google Scholar]
- Langworthy, M.; Finan, T.J. Waiting for Rain: Agriculture and Ecological Imbalance in Cape Verde, 1st ed.; Lynne Rienner Publishers: Boulder, CO, USA, 1997; p. 230. [Google Scholar]
- Castilla-Beltrán, A.; de Nascimento, L.; Fernández-Palacios, J.M.; Fonville, T.; Whittaker, R.J.; Edwards, M.; Nogué, S. Late Holocene environmental change and the anthropisation of the highlands of Santo Antão Island, Cabo Verde. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 524, 101–117. [Google Scholar] [CrossRef]
- Bosa, M.S. Water institutions and management in Cape Verde. Water 2015, 7, 2641–2655. [Google Scholar] [CrossRef] [Green Version]
- Agresti, A. Categorial Data Analysis; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2002. [Google Scholar]
- Pekár, S.; Brabec, M. Modern Analysis of Biological Data. 2. Linear Models with Correlation in R; Masaryk University Press: Brno, Czech Republic, 2012; p. 256. [Google Scholar]
- Humboldt, A.V. Relation Historique Du Voyage Aux Régions Équinoctiales Du Nouveau Continent; F. Schoell: Paris, France, 1814; Nachdruck Stuttgart 1970. [Google Scholar]
- Christ, H. Eine Frühlingsfahrt Zu Den Canarischen Inseln; H. Georg’s Verlag: Basel, Switzerland, 1886. [Google Scholar]
- Schenck, H. Beitrage Zur Kenntnis Der Vegetation Der Canarischen Inseln; Wissenschaftliche Ergebnisse der Deutschen Tiefsee-Expedition auf dem Dampfer “Valdivia”; Fischer: Jena, Germany, 1907; Volume 2/1, pp. 225–406. [Google Scholar]
- Mägdefrau, K. Das alter der drachenbäume auf Tenerife. Flora 1975, 164, 347–357. [Google Scholar] [CrossRef]
- Zheng, D.J.; Xie, L.S.; Zhu, J.H.; Zhang, Z.L. Low genetic diversity and local adaptive divergence of Dracaena cambodiana (Liliaceae) populations associated with historical population bottlenecks and natural selection: An endangered long-lived tree endemic to Hainan Island, China. Plant Biol. 2012, 14, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Usama, M. Dracaena Ombet Status Survey in Erkowit Mountains in East Sudan. Dragon Tree Land Project Report. 2015, p. 21. Available online: https://www.rufford.org/files/11512-1%20April%20update.pdf (accessed on 5 January 2020).
- Krawczyszyn, J.; Krawczyszyn, T. Photomorphogenesis in Dracaena draco. Trees Struct. Funct. 2016, 30, 647–664. [Google Scholar] [CrossRef] [Green Version]
- Byström, K. Dracaena draco L. in the Cape Verde Islands. Acta Horti Gotobg. 1960, 23, 179–214. [Google Scholar]
- Symon, D.E. The growth of Dracaena draco—Dragon’s blood tree. J. Arnold Arbor. 1974, 55, 51–58. [Google Scholar]
- Scholte, P.; De Geest, P. The climate of Socotra Island (Yemen). A first-time assessment of the timing of the monsoon wind reversal and its influence on precipitation and vegetation patterns. J. Arid Environ. 2010, 74, 1507–1515. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Eltahir, E.A.B. Using a horizontal precipitation model to investigate the role of turbulent cloud deposition in survival of a seasonal cloud forest in Dhofar. J. Geophys. Res. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, A.; Eltahir, E.A.B. Forest on the edge: Seasonal cloud forest in Oman creates its own ecological niche. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Abdul-Wahab, S.A.; Al-Hinai, H.; Al-Najar, K.A.; Al-Kalbani, M.S. Fog water harvesting: Quality of fog water collected for domestic and agricultural use. Environ. Eng. Sci. 2007, 24, 446. [Google Scholar] [CrossRef]
- Marzol, V.M.; Sánchez, J.L.; Yanes, A. Meteorological patterns and fog water collection in Morocco and the Canary Islands. Erdkunde 2011, 65, 291–303. [Google Scholar] [CrossRef]
- Martorell, C.; Ezcurra, E. Rosette scrub occurrence and fog availability in arid mountains of Mexico. J. Veg. Sci. 2002, 13, 651–662. [Google Scholar] [CrossRef]
- Denk, T.; Güner, H.T.; Grimm, G.W. From mesic to arid: Leaf epidermal features suggest preadaptation in Miocene dragon trees (Dracaena). Rev. Paleobotany Palynol. 2014, 200, 211–228. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Long-term in situ sap flow monitoring in a mature Dracaena cinnabari tree on Socotra. Biologia 2019. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Nadezhdin, V. Are Dracaena nebulophytes able to drink atmospheric water? Environ. Exp. Bot. 2017, 139, 57–66. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Sap flow measurements in a Socotra dragon’s blood tree (Dracaena cinnabari) in its area of origin. Trop. Plant Biol. 2018, 11, 107–118. [Google Scholar] [CrossRef]
- Adolt, R.; Maděra, P.; Abraham, J.; Čupa, P.; Svátek, M.; Matula, R.; Šebesta, J.; Čermák, M.; Volařík, D.; Koutecký, T.; et al. Field survey of Dracaena cinnabari populations in Firmihin, Socotra island: Methodology and preliminary results. J. Landsc. Ecol. 2013, 6, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Wilkin, P.; Suksathan, P.; Keeratikiat, K.; Van Welzen, P.; Wiland-Szymanska, J. A new threatened endemic species from central and northeastern Thailand, Dracaena jayniana (Asparagaceae: Tribe Nolinoideae). Kew Bull. 2012, 67, 697–705. [Google Scholar] [CrossRef]
- Wilkin, P.; Suksathan, P.; Keeratikiat, K.; Van Welzen, P.; Wiland-Szymanska, J. A new species from Thailand and Burma, Dracaena kaweesakii Wilkin & Suksathan (Asparagaceae subfamily Nolinoideae). PhytoKeys 2013, 26, 101–112. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Annual Temperature (°C) | Max Temperature of Warmest Month (°C) | Min Temperature of Coldest Month (°C) | Mean Annual Precipitation (mm) | Precipitation of Wettest Month (mm) | Precipitation of Driest Month (mm) | |
|---|---|---|---|---|---|---|
| Do | 21.1 | 31.6 | 10.3 | 599 | 146 | 21 |
| Dd | 17.4 | 22.1 | 12.9 | 314.7 | 99.5 | 0 |
| Number of Branch Order | Number of Trees D. ombet | Number of Trees D. caboverdeana |
|---|---|---|
| 1 | 7 | 24 |
| 2 | 7 | 8 |
| 3 | 6 | 7 |
| 4 | 6 | 1 |
| 5 | 8 | 4 |
| 6 | 7 | 1 |
| 7 | 6 | 4 |
| 8 | 8 | 3 |
| 9 | 10 | 1 |
| 10 | 7 | 0 |
| 11 | 6 | 3 |
| 12 | 7 | 3 |
| 13 | 6 | 0 |
| 14 | 6 | 1 |
| 15 | 4 | 4 |
| 16 | 3 | 2 |
| 17 | 3 | 3 |
| 18 | 2 | 1 |
| 19 | 0 | 6 |
| 20 | 0 | 0 |
| 21 | 0 | 3 |
| 22 | 0 | 3 |
| Total number | 109 | 82 |
| Coefficients | Coeff. Estimate | t Value | p-Value | |
|---|---|---|---|---|
| D. draco | Intercept (ß0) | −1.405075 | −6.191 | ˂0.001 |
| Branch orders (ß1) | 0.002147 | 0.162 | 0.872 | |
| D. ombet | Intercept (ß0) | −1.68578 | −6.961 | ˂0.001 |
| Branch orders (ß1) | 0.02132 | 1.068 | 0.288 |
| Number of Branch Order | Age of D. ombet | Age of D. draco subsp. caboverdeana |
|---|---|---|
| 1 | 5.2 | 4.9 |
| 2 | 10.5 | 9.9 |
| 3 | 15.7 | 14.8 |
| 4 | 20.9 | 19.7 |
| 5 | 26.2 | 24.7 |
| 6 | 31.4 | 29.6 |
| 7 | 36.6 | 34.6 |
| 8 | 41.9 | 39.5 |
| 9 | 47.1 | 44.4 |
| 10 | 52.3 | 49.3 |
| 11 | 57.5 | 54.3 |
| 12 | 62.8 | 59.2 |
| 13 | 68.0 | 64.2 |
| 14 | 73.2 | 69.1 |
| 15 | 78.5 | 74.1 |
| 16 | 83.7 | 79.0 |
| 17 | 88.9 | 83.9 |
| 18 | 94.2 | 88.9 |
| 19 | - | 93.8 |
| 20 | - | 98.7 |
| 21 | - | 103.7 |
| 22 | - | 108.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lengálová, K.; Kalivodová, H.; Habrová, H.; Maděra, P.; Tesfamariam, B.; Šenfeldr, M. First Age-Estimation Model for Dracaena ombet and Dracaena draco subsp. caboverdeana. Forests 2020, 11, 264. https://doi.org/10.3390/f11030264
Lengálová K, Kalivodová H, Habrová H, Maděra P, Tesfamariam B, Šenfeldr M. First Age-Estimation Model for Dracaena ombet and Dracaena draco subsp. caboverdeana. Forests. 2020; 11(3):264. https://doi.org/10.3390/f11030264
Chicago/Turabian StyleLengálová, Klára, Hana Kalivodová, Hana Habrová, Petr Maděra, Berihu Tesfamariam, and Martin Šenfeldr. 2020. "First Age-Estimation Model for Dracaena ombet and Dracaena draco subsp. caboverdeana" Forests 11, no. 3: 264. https://doi.org/10.3390/f11030264