What We Know and What We Do Not Know about Dragon Trees?

 , , and
, , and
Abstract
:1. Introduction–Dragon Trees, Tertiary Relicts in Current Reality
- Some species are threatened and are listed in the IUCN Red List [22]. In fact, of the six species that have been assessed, five are threatened, while the other 11 species have not yet been assessed.
- Most species are present in developing countries with an unstable political situation, and populations are under pressure, as weak governments have insufficient resources to protect them.
2. Evolution, Taxonomy and Distribution
2.1. Current Taxonomical Knowledge and Nomenclature
2.2. Evolution of Arborescent Dracaenas
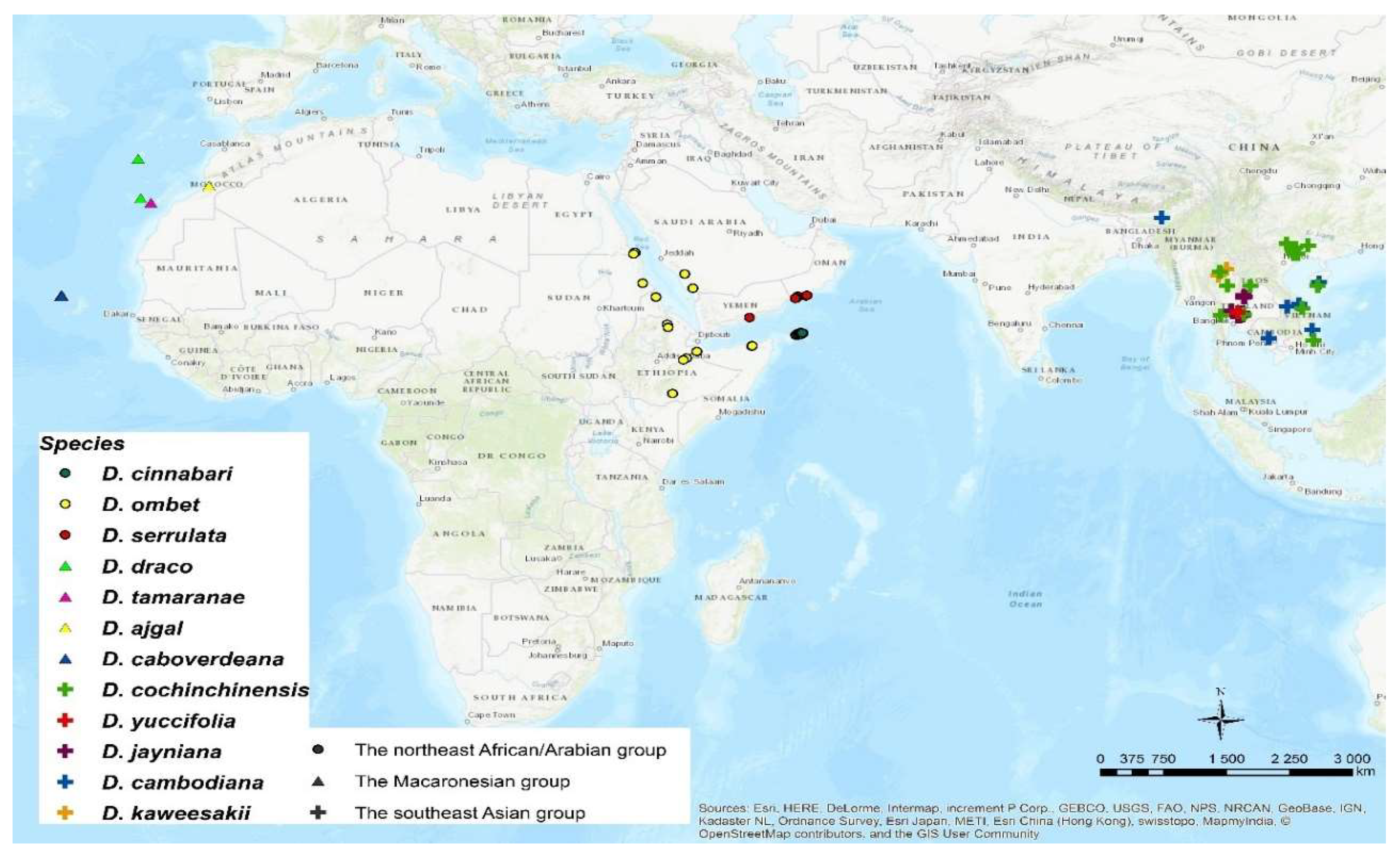
2.3. Geographical Distribution
3. Morphology and Anatomy
3.1. Dracaena Habitus/Shape and its Modifications by Environmental Factors
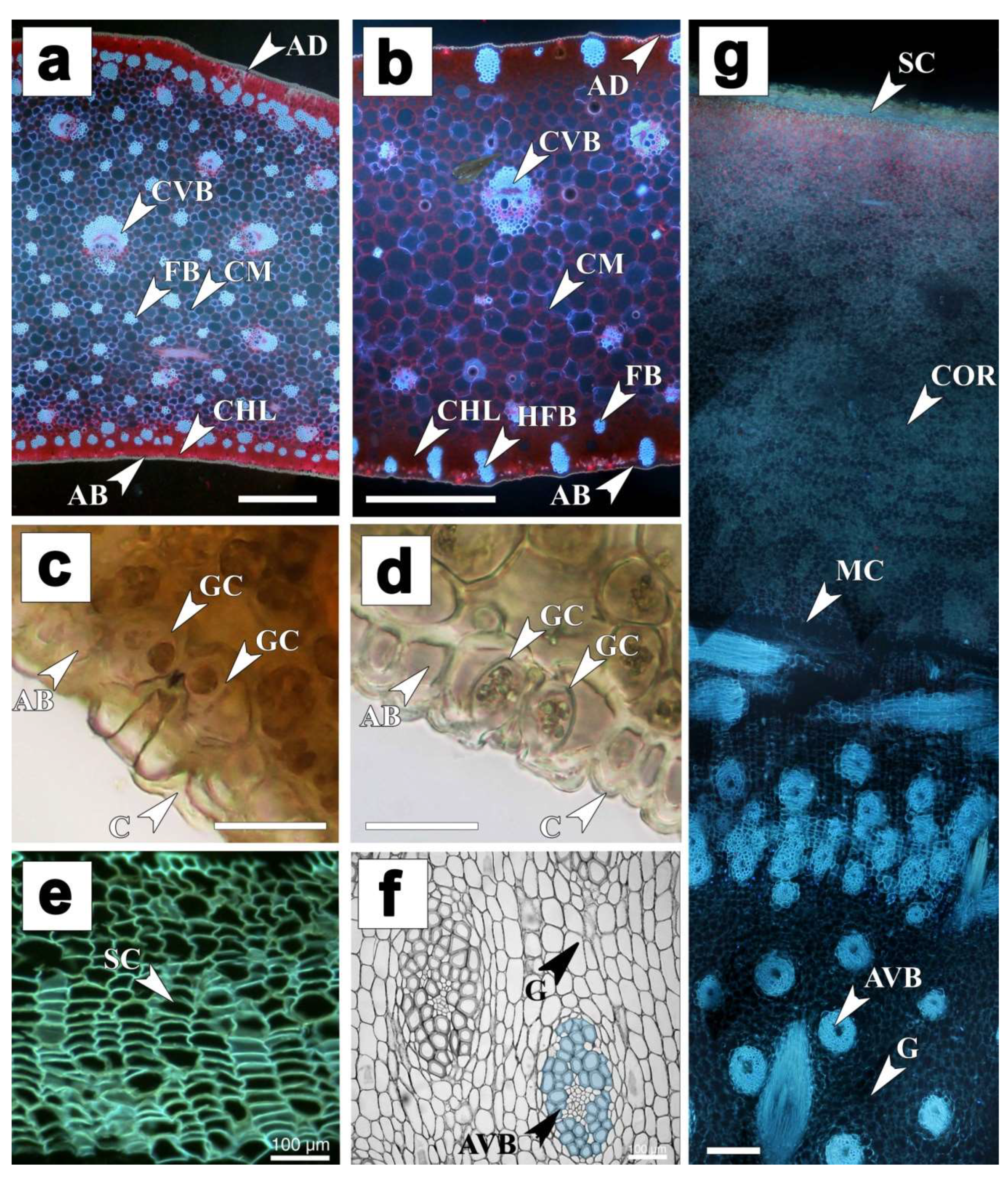
3.2. Leaf Micromorphology and Anatomy—From Mesomorphic to Xeromorphic Leaves
3.3. Secondary Growth in Stem and Roots
3.4. Dracaena Resin Secretion
4. Ontogenetic Cycle and Its Lifespan
5. Water Relations
6. Dracaena Resin and Ethnobotanical Use of Dragon Trees
6.1. Chemical Diversity of Dracaena Resins
6.2. Bioactive Properties of Dracaena Resins
6.3. Historical Records of Dracaena Resins
6.4. The Management and Harvesting of Dracaena cinnabari Resin
6.5. Other Uses of Dracena Species
7. Ecology of Plant Communities with Dragon Trees
8. Populations, Threat, Nature Conservation
8.1. Population Size, Abundance
8.2. Population Structure
8.3. Threats and Conservation Status
8.4. Rescue Programs
9. Future Research Needed
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mabberley, D.J. Mabberley’s Plant-Book, 3rd ed.; Cambridge University Press: New York, NY, USA, 2008; p. 1021. [Google Scholar]
- Bos, J. Dracaena. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; Volume III, pp. 238–241. [Google Scholar]
- Govaerts, R.; Zonneveld, B.J.M.; Zona, S.A.; World checklist of Asparagaceae. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://apps.kew.org/wcsp/ (accessed on 22 December 2019).
- Marrero, A.; Almeida, S.R.; Martín-González, M. A new species of the wild Dragon Tree, Dracaena (Dracaenaceae) from Gran Canaria and its taxonomic and biogeographic Implications. Bot. J. Linn. Soc. 1998, 128, 291–314. [Google Scholar]
- Marrero, A. Dracaena tamaranae, el género dracaena y otros afines: Análisis morfológico para un aproximación filogenética. El Mus. Canar. 2000, 55, 301–334. [Google Scholar]
- Bos, J.J.; Teketay, D. Dracaenaceae. In Flora of Ethiopia and Eritrea, 1st ed.; Edwards, S., Ed.; Uppsala University: Uppsala, Sweden, 1997; Volume 5, pp. 76–84. [Google Scholar]
- Marrero, A.; Almeida, S.R. A new subspecies, Dracaena draco (L.) L. subsp. caboverdeana Marrero Rodr. & R. Almeida (Dracaenaceae) from Cape Verde Island. Int. J. Geobot. Res. 2012, 2, 35–40. [Google Scholar] [CrossRef]
- Benabid, A.; Cuzin, F. Populations de dragonnier (Dracaena draco L. subsp. aigal Benabid et Cuzin) au Maroc: Valeurs taxinomique, biogéographique et phytosociologique. C. R. Acad. Sci. Paris, Sciences de la vie 1997, 320, 267–277. [Google Scholar]
- Wilkin, P.; Suksathan, P.; Keeratikiat, K.; Van Welzen, P.; Wiland-Szymanska, J. A new threatened endemic species from central and northeastern Thailand, Dracaena jayniana (Asparagaceae: Tribe Nolinoideae). Kew. Bull. 2012, 67, 697–705. [Google Scholar] [CrossRef]
- Wilkin, P.; Suksathan, P.; Keeratikiat, K.; Van Welzen, P.; Wiland-Szymanska, J. A new species from Thailand and Burma, Dracaena kaweesakii Wilkin & Suksathan (Asparagaceae subfamily Nolinoideae). PhytoKeys 2013, 26, 101–112. [Google Scholar] [CrossRef]
- Zheng, D.J.; Xie, L.S.; Zhu, J.H.; Zhang, Z.L. Low genetic diversity and local adaptive divergence of Dracaena cambodiana (Liliaceae) populations associated with historical population bottlenecks and natural selection: An endangered long-lived tree endemic to Hainan Island, China. Plant Biol. 2012, 14, 828–838. [Google Scholar] [CrossRef]
- Zona, S.; Álvarez De Zayas, A.; Orellana, R.; Oviedo, R.; Jestrow, B.; Francisco-Ortega, J. Dracaena L. (Asparagaceae) in the New World: Its history and Botany. Vieraea 2014, 42, 219–240. [Google Scholar]
- Denk, T.; Güner, H.T.; Grimm, G.W. From mesic to arid: Leaf epidermal features suggest preadaptation in Miocene dragon trees (Dracaena). Rev. Paleobotany Palynol. 2014, 200, 211–228. [Google Scholar] [CrossRef]
- Adolt, R.; Pavliš, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees-Struct. Funct. 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Bayley Balfour, I. Botany of Socotra, 1st ed.; Royal Society of Edinburgh: Edinburgh, UK, 1888; p. 446. [Google Scholar]
- Almeida Pérez, R.S. Censo, distribución, habitat y estado de conservation de Dracaena tamaranae A.Marrero, R.S.González-Martín. Gran Canaria, Islas Canarias. Bot. Macarónesica 2003, 24, 39–56. [Google Scholar]
- Lavranos, J.J. A new, arborescent subspecies of Dracaena from Saudi Arabia. Cactus Succul. J. 2017, 89, 148–152. [Google Scholar] [CrossRef]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saed, A.; Alfo, M.; Bruno, F. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]
- Van Damme, K.; Banfield, L. Past and present human impacts on the biodiversity of Socotra Island (Yemen): Implications for future conservation. Zool. Middle East 2011, 3, 31–88. [Google Scholar] [CrossRef]
- Kamel, M.; Ghazaly, U.M.; Callmander, M.W. Conservation status of the Endangered Nubian dragon tree Dracaena ombet in Gebel Elba National Park, Egypt. Oryx 2015, 49, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Čermák, Z.; Pavliš, J. Dragon’s blood tree-Threatened by overmaturity, not by extinction: Dynamics of a Dracaena cinnabari woodland in the mountains of Soqotra. Biol. Conserv. 2009, 142, 772–778. [Google Scholar] [CrossRef]
- IUCN red list of threatened species version 2017.2. Available online: http://www.iucnredlist.org (accessed on 25 October 2017).
- Brown, G.; Mies, B.A. Vegetation Ecology of Socotra. Plant and Vegetation 7, 1st ed.; Springer: Dordrecht, Germany, 2012; p. 379. [Google Scholar]
- Habrová, H. Geobiocoenological differentiation as a tool for sustainable land-use of Soqotra Island (Republic of Yemen). Ekológia Bratisl. 2004, 23, 47–57. [Google Scholar]
- Hubálková, I. Prediction of Dragon’s Blood Tree (Dracaena cinnabari Balf.) stand sample density on Soqotra Island. J. Landsc. Ecol. 2011, 4, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Pavliš, J. Dynamic response of woody vegetation on fencing protection in semi-arid areas; Case study: Pilot exclosure on the Firmihin Plateau, Socotra island. Saudi J. Biol. Sci. 2017, 24, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.G.; Morris, M.; Diccon, A.; Atkinson, R. Ethnoflora of the Soqotra Archipelago, 1st ed.; Royal Botanic Garden: Edinburgh, UK, 2004; p. 759. [Google Scholar]
- Rejžek, M.; Svátek, M.; Šebesta, J.; Adolt, R.; Maděra, P.; Matula, R. Loss of a single tree species will lead to an overall decline in plant diversity: Effect of Dracaena cinnabari Balf. f. on the vegetation of Socotra Island. Biol. Conserv. 2016, 196, 165–172. [Google Scholar] [CrossRef]
- García, C.; Vasconcelos, R. The beauty and the beast: Endemic mutualistic interactions promote community-based conservation on Socotra Island (Yemen). J. Nature Conserv. 2017, 35, 20–23. [Google Scholar] [CrossRef]
- González-Castro, A.; Pérez-Pérez, D.; Romero, J.; Nogales, M. Unraveling the Seed Dispersal System of an Insular “Ghost” Dragon Tree (Dracaena draco) in the Wild. Front. Ecol. Evol. 2019, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, A.; Eltahir, E.A.B. Forest on the edge: Seasonal cloud forest in Oman creates its own ecological niche. Geophys. Res. Lett. 2006, 33, L11401. [Google Scholar] [CrossRef]
- Scholte, P.; Al-Okaishi, A.; Suleyman, A.S. When conservation precedes development: A case study of the opening up of the Socotra archipelago, Yemen. Oryx 2011, 45, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.-L.; Morden, C.W. Phylogenetic Relationships among Dracaenoid Genera (Asparagaceae: Nolinoideae) Inferred from Chloroplast DNA Loci. Syst. Bot. 2014, 39, 90–104. [Google Scholar] [CrossRef]
- Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.-L.; Morden, C.W. Phylogenetics of the plant genera Dracaena and Pleomele (Aparagaceae). Bot. Orient. J. Plant Sci. 2010, 7, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Kuntze, O. Revisio generum plantarum: vascularium omnium atque cellularium multarum secundum leges nomenclaturae internationales cum enumeratione plantarum exoticarum in itinere mundi collectarum, 2nd ed.; A. Felix: Leipzig, Germany, 1891; p. 374. [Google Scholar]
- Bos, J.J. Dracaena in West Africa. Meded. Landbouwhogesch. Wagening. 1984, 84, 1–126. [Google Scholar]
- Brown, N.E. Notes on the genera Cordyline, Dracaena, Pleomele, Sansevieria, and Taetsia. Bull. Misc. Inf. (Royal Bot. Gard. Kew.) 1914, 8, 273–279. [Google Scholar] [CrossRef]
- Wiland-Szymańska, J.; Klimko, M. Differentiation of leaf anatomy of the genera Dracaena L. and Sansevieria Thunb. (Dracaenaceae). In Abstracts XVII International Botanical Congress; International Association of Botanical and Mycological Societies: Vienna, Austria, 16 July 2005; p. 328. [Google Scholar]
- Jankalski, S. The Sansevieria inflorescence and new sections proposed. Sansevieria 2009, 19, 8–10. [Google Scholar]
- Mansfeld, P. Die Systematik der Gattung Sansevieria (Asparagaceae)–ein aktueller Stand. Sansevieria Online 2015, 3, 20–29. [Google Scholar]
- Takawira-Nyenya, R.; Mucina, L.; Cardinal-Mcteague, W.; Thiele, K. Sansevieria (Asparagaceae, Nolinoideae) is a herbaceous clade within Dracaena: Inference from non-coding plastid and nuclear DNA sequence data. Phytotaxa 2018, 376, 254–276. [Google Scholar] [CrossRef]
- Klimko, M.; Nowińska, R.; Wilkin, P.; Wiland-Szymańska, J. Comparative leaf micromorphology and anatomy of the dragon tree group of Dracaena (Asparagaceae) and their taxonomic implications. Plant Syst. Evol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Klimko, M.; Nowińska, R.; Jura-Morawiec, J.; Wiland-Szymańska, J.; Wilkin, P. Pollen morphology of selected species of the genera Chrysodracon and Dracaena (Asparagaceae, subfamily Nolinoideae) and its systematic implications. Plant Syst. Evol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Rodríguez, C. A Dracaenites Saporta fossil in the Pliocene of Gran Canaria, Canary Islands. Vieraea 2013, 41, 385–388. [Google Scholar]
- Baldwin, A.; Webb, R. The Genus Sansevieria: An introduction to molecular (DNA) analysis and preliminary insights to intrageneric relationships. Sansevieria 2016, 34, 14–26. [Google Scholar]
- Zhang, Z.; Zhang, Y.; Song, M.; Guan, Y.; Ma, X. Species identification of Dracaena using the complete chloroplast genome as a super-barcode. Front. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Zhua, Z.-X.; Mub, W.-X.; Wanga, J.-H.; Zhanga, J.-R.; Zhaoa, K.-K.; Friedmanc, C.R.; Wanga, H.-F. Complete plastome sequence of Dracaena cambodiana (Asparagaceae): A species considered “Vulnerable” in Southeast Asia. Mitochondrial DNA Part B Resour. 2018, 3, 620–621. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.-L.; Zhang, L.; Dayanandan, S.; Nagaraju, S.; Liu, D.-M.; Li, Q.-M. Tertiary origin and pleistocene diversification of dragon blood tree (Dracaena cambodiana-Asparagaceae) populations in the Asian tropical forests. PLoS ONE 2013, 8, e60102. [Google Scholar] [CrossRef] [Green Version]
- Rivas Martinez, S. Ensayo geobotánico global sobre la Macaronesia. In Homenaje al Prof Dr. Wolfredo Wildproel de la Torre, 1st ed.; Beltrán Tejera, E., Afonso-Carrillo, J., García Gallo, A., Rodríguez Delgado, O., Eds.; Instituto de Estudios Canarios: La Laguna (Santa Cruz de Tenerife), Spain, 2009; pp. 256–296. [Google Scholar]
- Almeida Pérez, R.S. Sobre la presencia de Dracaena draco (L.) L. En Gran Canaria (Islas Canarias): Aportación corológica, estado actual y significación biogeográfica. Bot. Macarónesica 2003, 24, 17–38. [Google Scholar]
- Turland, N.J. Agavaceae. In Flora of Madeira; Press, J.R., Short, M.J., Eds.; HMSO, Natural History Museum: London, UK, 1995; pp. 391–392. [Google Scholar]
- Santos, A. Vegetación y flora de La Palma; Editorial Interinsular Canaria: Santa Cruz de Tenerife, Spain, 1983. [Google Scholar]
- Rivas-Martínez, S.; Wildpret, W.; Del Arco, M.; Rodríguez, O.; Pérez de Paz, P.L.; Garcia-Gallo, A.; Acebes, J.R.; Díaz, T.E.; Fernández-González, F. Las comunidades vegetales de la isla de Tenerife (islas Canarias). Itinera Geobotánica 1993, 7, 169–374. [Google Scholar]
- Král, K.; Pavliš, J. The first detailed land cover map of Socotra Island by Landsat /ETM+ data. Int. J. Remote Sens. 2006, 27, 3239–3250. [Google Scholar] [CrossRef]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable land use management needed to conserve the dragon´s blood tree of Socotra Island, a vulnerable endemic umbrella species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef] [Green Version]
- Collenette, S. An Illustrated Guide to the Flowers of Saudi Arabia; Scorpion Publishing: London, UK, 1985; p. 266. [Google Scholar]
- Miller, A.G.; Cope, T.A. Flora of the Arabian Peninsula and Socotra; Edinburgh University Press: Edinburgh, UK, 1996; Volume 1, p. 438. [Google Scholar]
- Ghazali, U. The globally endangered Dracaena ombet monitoring and assessment project in Gabel Elba protected area, Egypt. Final report, conservation leadership programme. 2008. Available online: http://www.conservationleadershipprogramme.org/ (accessed on 5 December 2013).
- Zahran, M.A.; Willis, A.J. The Vegetation of Egypt; Springer: Berlin/Heidelberg, Germany, 2009; p. 437. [Google Scholar]
- Friis, I. Forests and Forest Trees of Northeast Tropical Africa; Their natural habitats and distribution patterns in Ethiopia, Djibouti and Somalia; HMSO: Middlesex, UK, 1992; p. 396. [Google Scholar]
- El Azzouni, M. Conserving Dracaena Ombet, Egypt’s Dragon Tree. PlantTalk 2003, 3, 38–39. [Google Scholar]
- Usama, M. Dracaena Ombet Status Survey in Erkowit Mountains in East Sudan; Project Report; Dragon Tree Land Project: Mount Erkowit, Sudan, 2015; p. 21. [Google Scholar]
- Baker, J.G. Dracaena schizantha Baker sp. nov. In The Journal of Botany, New Series; Trimen, H., Ed.; Ranken & Co.: London, UK, 1877; Volume 6, p. 71. [Google Scholar]
- Thulin, M. Dracaenaceae. In Flora of Somalia; Thulin, M., Ed.; Royal Botanic Gardens Kew: London, UK, 1995; Volume 4, pp. 27–29. [Google Scholar]
- Chen, X.; Turland, N.J. Dracaena Vandelli ex Linnaeus. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press, and Missouri Botanical Garden Press: Beijing, China; Louis, MO, USA, 2000; Volume 24, pp. 73–263. [Google Scholar]
- Sarma, J.; Barbhuiya, H.A.; Dey, S. First report of a strict Dragon Tree species (Dracaena cambodiana: Asparagaceae) from India. J. Bot. Res. Inst. of Texas, 2019, 13, 241–247. [Google Scholar]
- Wang, X.H.; Zhang, C.H.; Yang, L.L.; Gomes-Laranjo, J. Production of dragon’s blood in Dracaena cochinchinensis plants by inoculation of Fusarium proliferatum. Plant Sci. 2011, 180, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.Y.; Yi, T.; Sze-To, C.M.; Zhu, L.; Peng, W.L.; Zhang, Y.Z.; Zhao, Z.Z.; Chen, H.B. A Systematic Review of the Botanical, Phytochemical and Pharmacological Profile of Dracaena cochinchinensis, a Plant Source of the Ethnomedicine “Dragon’s Blood”. Molecules 2014, 19, 10650–10669. [Google Scholar] [CrossRef] [Green Version]
- Beyhl, F.E. Two different growth forms of Dracaena draco L. (Monocotyledones: Liliales: Agavaceae). Bol. Mus. Munic. Funchal. 1995, 4, 91–95. [Google Scholar]
- Cheadle, V.I. Secondary growth by means of a thickening ring in certain monocotyledons. Bot. Gaz. 1937, 98, 535–555. [Google Scholar] [CrossRef]
- Carlquist, S. Monocot Xylem Revisited: New Information, New Paradigms. Bot. Rev. 2012, 78, 87–153. [Google Scholar] [CrossRef]
- Hubálková, I.; Houška, J.; Kubíček, J.; Mazal, P.; Pavliš, J.; Pohořalý, J.; Vačkářová, G.; Duchoslav, M. Preliminary anatomical study on secondary thickening parts of endemic Dracaena cinnabari Bal.fil. from the Soqotra island. Wood Res. 2017, 62, 67–77. [Google Scholar]
- Krawczyszyn, J.; Krawczyszyn, T. Massive aerial roots growth and form of Dracaena draco. Trees-Struct. Funct. 2014, 28, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Beyhl, F.E. Dragon-trees (Dracaena draco L.) with abberant growth forms (Monocotyledons: Liliflorae: Agavaceae). Arquipélago. Life Mar. Sci. 2001, 2, 101–103. [Google Scholar]
- Krawczyszyn, J.; Krawczyszyn, T. Photomorphogenesis in Dracaena draco. Trees-Struct. Funct. 2016, 30, 647–664. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.H.; Tomlinson, P.B. The vascular system in the axis of Dracaena fragrans (Agavaceae) 1. Distribution and development of primary strands. J. Arnold Arbor. 1969, 51, 370–383. [Google Scholar]
- Haushahn, T.; Speck, T.; Masselter, T. Branching morphology of decapitated arborescent monocotyledons with secondary growth. Am. J. Bot. 2014, 101, 754–763. [Google Scholar] [CrossRef]
- Byström, K. Dracaena draco L. in the Cape Verde Islands. Acta Horti.-Gotobg. 1960, 23, 179–214. [Google Scholar]
- Lyons, G. In search of dragons or: The plant that roared. Cactus Succul. J. 1974, 46, 267–282. [Google Scholar]
- Beyhl, F.E.; Mies, B.A. Die Wuchsformen des Drachenbaums (Dracaena cinnabari Balf. Fil.) auf der Insel Soqotra-growth forms of the Dragon tree (Dracaena cinnabari Balf. Fil.) of Soqotra island. Acta Biol. Benrodis 2007, 14, 55–66. [Google Scholar]
- Jura-Morawiec, J. Rhythmic growth and age estimation of aerial roots in Dracaena draco (Asparagaceae). Trees 2019, 33, 1513–1518. [Google Scholar] [CrossRef] [Green Version]
- WCSP World Checklist of Selected Plant Families. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://wcsp.science.kew.org/ (accessed on 29 November 2019).
- Nadezhdina, N.; Plichta, R.; Nadezhdin, V.; Gebauer, R.; Jupa, R.; Habrová, H.; Maděra, P. A comparative structural and functional study of leaf traits and sap flow in Dracaena cinnabari and Dracaena draco seedlings. Funct. Plant Biol. 2015, 42, 1092–1105. [Google Scholar] [CrossRef]
- Rudall, P.; Conran, J.G.; Chase, M.W. Systematics of Ruscaceae/Convallariaceae: A combined morphological and molecular investigation. Bot. J. Linn. Soc. 2000, 134, 73–92. [Google Scholar] [CrossRef]
- Jupa, R.; Plichta, R.; Paschová, Z.; Nadezhda, N.; Gebauer, R. Mechanisms underlying the long-term survival of the monocot Dracaena marginata under drought conditions. Tree Physiol. 2017, 37, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimko, M.; Wiland-Szymańska, J. Scanning electron microscopic studies of leaf surface in taxa of genus Dracaena L. (Dracaenaceae). Bot.–Steciana 2008, 12, 117–127. [Google Scholar]
- Rudall, P. Lateral meristems and stem thickening growth in monocotyledons. Bot. Rev. 1991, 57, 150–163. [Google Scholar] [CrossRef]
- Rudall, P.J. New records of secondary thickening in monocotyledons. Iawa J. 1995, 16, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Jura-Morawiec, J.; Tulik, M.; Iqbal, M. Lateral Meristems Responsible for Secondary Growth of the Monocotyledons: A Survey of the State of the Art. Bot. Rev. 2015, 81, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, P.B.; Zimmermann, M.H. Vascular anatomy of monocotyledons with secondary growth—An introduction. J. Arnold Arbor. 1969, 50, 159–179. [Google Scholar]
- Evert, R. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development, 3rd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2006; p. 624. [Google Scholar]
- Jura-Morawiec, J. Formation of amphivasal vascular bundles in Dracaena draco stem in relation to rate of cambial activity. Trees-Struct. Funct. 2015, 29, 1493–1499. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, M.H.; Tomlinson, P.B. The vascular system in the axis of Dracaena fragrans (Agavaceae) 2. Distribution and development of secondary tissue. J. Arnold Arbor. 1970, 51, 478–491. [Google Scholar]
- Jura-Morawiec, J. Atypical origin, structure and arrangement of secondary tracheary elements in the stem of the monocotyledonous dragon tree, Dracaena draco. Planta 2017, 245, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.H.; Brebner, G. On the secondary tissues in certain monocotyledons. Ann. Bot. 1893, 7, 22–62. [Google Scholar] [CrossRef]
- Waterhouse, J.T. The phylogenetic significance of Dracaena—Type growth. Proc. Linn. Soc. New South Wales 1987, 109, 129–138. [Google Scholar]
- De Bary, A. Comparative Anatomy of the Vegetative Organs of the Phanerogams and Ferns, 1st. ed.; Claredon Press: Oxford, UK, 1884. [Google Scholar]
- Jura-Morawiec, J.; Wiland-Szymanska, J. A novel insight into the structure of amphivasal secondary bundles on the example of Dracaena draco L. stem. Trees-Struct. Funct. 2014, 28, 871–877. [Google Scholar] [CrossRef]
- Zimmermann, M.H.; Brown, C.L. Trees Structure and Function; Springer: New York, NY, USA, 1971; p. 336. [Google Scholar]
- Spicer, R. Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport. J. Exp. Bot. 2014, 65, 1829–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.H.; Zhang, C.H.; Yang, X.H.; Lou, J.D.; Cao, Q.; Gomes Laranjo, J. Enhanced dragon’s blood prodution in Dracaena cochinchinesis by elicitation of Fusarium oxysporum strands. J. Med. Plants Res. 2010, 4, 2633–2640. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.L.; Wang, C.L.; Guo, S.X.; Xiao, P.G.; Wang, M.L. Stimulation of dragon’s blood accumulation in Dracaena cambodiana via fungal inoculation. Fitoterapia 2013, 87, 31–36. [Google Scholar] [CrossRef]
- Wang, X.H.; Zhang, W.Y.; Gomes-Laranjo, J. Screen of micro-organisms for inducing the production of dragon’s blood by leaf Dracaena cochinchinensis. Lett. Appl. Microbiol. 2010, 51, 504–510. [Google Scholar] [CrossRef]
- Ding, X.; Mei, W.; Huang, S.; Wang, H.; Zhu, J.; Hu, W.; Ding, Z.; Tie, W.; Peng, S.; Dai, H. Genome survey sequencing for the characterization of genetic background of Dracaena cambodiana and its defense response during dragon’s blood formation. PLoS ONE 2018, 13, e0209258. [Google Scholar] [CrossRef] [Green Version]
- Jura-Morawiec, J.; Tulik, M. Morpho-anatomical basis of dragon´s blood secret in Dracaena draco stem. Flora 2015, 213, 1–5. [Google Scholar] [CrossRef]
- Gupta, D.; Bleakley, B.; Gupta, R.K. Dragon’s blood: Botany, chemistry and therapeutic uses. J. Ethnopharmacol. 2008, 115, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H. Flavones formed from xylem stem of Dracaena cochinchinensis by co-culture of fungus strain and bacteria strain. Nat. Prod. Res. Dev. 2007, 19, 11–15. [Google Scholar]
- Jura-Morawiec, J.; Tulik, M. Dragon’s blood secretion and its ecological significance. Chemoecology 2016, 26, 101–105. [Google Scholar] [CrossRef] [Green Version]
- González, A.G.; Hernández, J.C.; Leôn, F.; Padrôn, J.I.; Estévez, F.; Quintana, J.; Bermejo, J. Steroidal saponins from the bark of Dracaena draco and their cytoctoxic activity. J. Nat. Prod. 2003, 66, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Habrová, H.; Maděra, P. Ecology of Dragon’s blood tree (Dracaena cinnabari) communities on the Socotra Island. In Evaluation of State and Development of Forest Geobiocoenosis; Faculty of Forestry and Wood, Technology, Polehla, P., Eds.; Mendel University: Brno, Czech Republic, 2004; pp. 120–126. [Google Scholar]
- Adolt, R.; Habrová, H.; Maděra, P. Crown age estimation of a monocotyledonous tree species Dracaena cinnabari using logistic regression. Trees-Struct. Funct. 2012, 26, 1287–1298. [Google Scholar] [CrossRef]
- Maděra, P.; Habrová, H.; Šenfeldr, M.; Kholová, I.; Lvončík, S.; Ehrenbergerová, L.; Roth, M.; Nadezhdina, N.; Němec, P.; Rosenthal, J.; et al. Growth dynamics of endemic Dracaena cinnabari Balf. f. of Socotra Island suggest essential elements for a conservation strategy. Biológia 2018. [Google Scholar] [CrossRef]
- Hubálková, I.; Maděra, P.; Volařík, D. Growth dynamics of Dracaena cinnabari under controlled conditions as the most effective way to protect endangered species. Saudi. J. Biol. Sci. 2017, 24, 1445–1452. [Google Scholar] [CrossRef] [Green Version]
- Symon, D.E. The growth of Dracaena draco-dragon’s blood tree. J. Arnold Arbor. 1974, 55, 51–58. [Google Scholar]
- Mägdefrau, K. Das Alter der Drachenbäume auf Tenerife. Flora 1975, 164, 347–357. [Google Scholar] [CrossRef]
- Pütter, A. Altersbestimmung an Drachenbäumen von Tenerife. Sitz. Der Heidelb. Akad. Der Wissenschäften. Math.-Nat. Kl. 1925, 12, 12–18. [Google Scholar]
- Humboldt, A.v. Relation historique du Voyage aux Régions équinoctiales du Nouveau Continent; F. Schoell: Paris, France, 1814. [Google Scholar]
- Christ, H. Eine Frühlingsfahrt zu den Canarischen Inseln; H. Georg’s Verlag: Basel, Switzerland, 1886. [Google Scholar]
- Schenck, H. Beitrage zur Kenntnis der Vegetation der Canarischen Inseln; Wissensch. Ergebn. d. dtsch. Tiefsee-Expedition "Valdivia", 2/1; Verlag G. Fischer,: Jena, Germany, 1907; pp. 225–406. [Google Scholar]
- Nadezhdina, N.; Čermák, J.; Nadezhdin, V. Heat field deformation method for sap flow measurements. In Measuring Sap Flow in Intact Plants, 1st ed.; Čermák, J., Nadezhdina, N., Eds.; Mendel University: Brno, Czech Republic, 1998; pp. 72–92. [Google Scholar]
- Nadezhdina, N.; Tributsch, H.; Čermák, J. Infra-red images of heat field around a linear heater and sap flow in stems of lime trees under natural and experimental conditions. Ann. For. Sci. 2004, 61, 203–213. [Google Scholar] [CrossRef]
- Nadezhdina, N. Revisiting the Heat Field Deformation (HFD) method for measuring sap flow. Iforest–Biogeosci. For. 2018, 11, 118–130. [Google Scholar] [CrossRef] [Green Version]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Long-term in situ sap flow monitoring in a mature Dracaena cinnabari tree on Socotra. Biologia 2019. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Nadezhdin, V. Are Dracaena nebulophytes able to drink atmospheric water? Environ. Exp. Bot. 2017, 139, 57–66. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Gebauer, R.; Nadezhdin, V.; Plichta, R. Water transport secrets of the dragon’s blood trees revealed through sap flow measurements following partial stem incision. Flora 2019, 250, 44–51. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Al-Okaishi, A.; Madera, P. Sap flow measurements in a Socotra dragon’s blood tree (Dracaena cinnabari) in its area of origin. Trop. Plant Biol. 2018, 11, 107–118. [Google Scholar] [CrossRef]
- Nadezhdina, N.; Nadezhdin, V.; Gebauer, R.; Čermák, J.; David, J.S.; David, T.S.; Jimenez, M.S.; Morales, D. Redistribution of water within the aboveground part of trees. Acta Hortic. 2012, 951, 241–249. [Google Scholar] [CrossRef]
- Sun, J.; Liu, J.N.; Fan, B.; Chen, X.N.; Pang, D.R.; Zheng, J.; Zhang, Q.; Zhao, Y.F.; Xiao, W.; Tu, P.F.; et al. Phenolic constituents, pharmacological activities, quality control, and metabolism of Dracaena species: A review. J. Ethnopharmacol. 2019, 224, 112138. [Google Scholar] [CrossRef]
- Edward, H.G.M.; Oliveira, L.F.C.; Quye, A. Raman spectroscopy of coloured resins used in antiquity: dragon’s blood and related substances. Spectrochim. Acta Part A 2001, 57, 2831–2842. [Google Scholar] [CrossRef]
- Vaníčková, L.; Pompeiano, A.; Maděra, P.; Massad, TJ.; Vahalík, P. Terpenoid profiles of resin in genus Dracaena are species specific. Phytochemistry 2020, 170, 112197. [Google Scholar] [CrossRef]
- González, A.G.; León, F.; Hernández, J.C.; Padrón, J.I.; Sánchez-Pinto, L.; Barrera, J.B. Flavans of dragon’s blood from Dracaena draco and Dracaena tamaranae. Biochem. Syst. Ecol. 2004, 32, 179–184. [Google Scholar] [CrossRef]
- Yi, T.; Chen, H.B.; Zhao, Z.Z.; Yu, Z.L.; Jiang, Z.H. Comparison of the chemical profiles and anti-platelet aggregation effects of two “Dragon’s Blood” drugs used in traditional Chinese medicine. J. Ethnopharmacol. 2011, 133, 796–802. [Google Scholar] [CrossRef] [Green Version]
- Israa, A.I.; Haider, M.H.; Imad, H.H. Bioactivities, characterization, and therapeutic uses of Dracaena cinnabari. Int. J. Pharm. Qual. Assur. 2018, 9, 11–14. [Google Scholar] [CrossRef]
- Masaoud, M.; Schmidt, J.; Adam, G. Sterols and triterpenoids from Dracaena cinnabari. Phytochemistry 1995, 38, 795–796. [Google Scholar] [CrossRef]
- Zheng, Q.A.; Li, H.Z.; Zhang, Y.J.; Yang, C.R. Dracaenogenins A and B, new spirostanols from the red resin of Dracaena cochinchinensis. Steroids 2006, 71, 160–164. [Google Scholar] [CrossRef]
- Zheng, Q.A.; Zhang, Y.J.; Yang, C.R. A new meta-homoisoflavane from the fresh stems of Dracaena cochinchinensis. J. Asian Nat. Prod. Res. 2006, 8, 571–577. [Google Scholar] [CrossRef]
- Hernández, J.C.; León, F.; Quintana, J.; Estévez, F.; Bermejo, J. Icogenin, a new cytotoxic steroidal saponin isolated from Dracaena draco. Bioorganic Med. Chem. 2004, 12, 4423–4429. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Ide Atsushi Kameyama, A.; Yokosuka, A.; Sashida, Y. Steroidal saponins from the aerial parts of Dracaena draco and their cytostatic activity on HL-60 cells. Phytochemistry 1999, 50, 805–813. [Google Scholar] [CrossRef]
- Zheng, Q.A.; Zhang, Y.J.; Li, H.Z.; Yang, C.R. Flavonoids from Dragon’s Blood of Dracaena cochinchinensis. Helv. Chim. Acta 2004, 87, 1267–1271. [Google Scholar] [CrossRef]
- Zheng, Q.A.; Zhang, Y.J.; Li, H.Z.; Yang, C.R. Steroidal saponins from fresh stems of Dracaena cochinchinensis. Steroids 2004, 69, 111–119. [Google Scholar] [CrossRef]
- Gurib-Fakim, A.; Demarne, F. Volatile constituents of Dracaena reflexa Lam. var. angustifolia Baker. J. Essent. Oil Res. 1994, 6, 651–652. [Google Scholar] [CrossRef]
- Santos, R.P.; Mendes, L.S.; Silva, B.M.; de Pinho, P.G.; Valentao, P.; Andrade, P.B.; Pereira, J.A.; Carvalho, M. Phytochemical profiles and inhibitory effect on free radical-induced human erythrocyte damage of Dracaena draco leaf: A potential novel antioxidant agent. Food Chem. 2011, 124, 927–934. [Google Scholar] [CrossRef]
- Teng, Z.; Zhang, M.; Meng, S.; Dai, R.; Meng, W.; Deng, Y.; Huang, L. A comparative study on volatile metabolites profile of Dracaena cochinchinensis (Lour.) S.C. Chen xylem with and without resin using GC-MS. Biomed. Chromatogr. 2015, 29, 1744–1749. [Google Scholar] [CrossRef] [PubMed]
- Masaoud, M.; Himmelreich, H.; Ripperger, H.; Adam, G. New bioflavonoids from Dracaena cinnabari. Planta Med. 1995, 61, 341–344. [Google Scholar] [CrossRef]
- Masaoud, M.; Ripperger, H.; Himmelreich, U.; Adam, G. Cinnabarone, a biflavonoid from dragon’s blood of Dracaena cinnabari. Phytochemistry 1995, 38, 751–753. [Google Scholar] [CrossRef]
- Lu, W.J.; Wang, X.; Chen, J.; Lu, Y.; Wu, N.; Kang, W.; Zheng, Q. Studies on the chemical constituents of chloroform extract of Dracaena cochinchinensis. Acta Pharm. Sin. 1998, 33, 755–758. [Google Scholar]
- Melo, M.J.; Sousa, M.; Parola, A.J.; de Melo, J.S.; Catarino, F.; Marcalo, J.; Pina, F. Identification of 7,4′ -Dihydroxy-5-methoxyflavylium in “Dragon’s Blood”: To be or not to be an anthocyanin. Chemistry 2006, 13, 1417–1422. [Google Scholar] [CrossRef] [Green Version]
- Veselá, D.; Marek, R.; Ubik, K.; Lunerová, K.; Sklenář, V.; Suchý, V. Dracophane, a metacyclophane derivative from the resin of Dracaena cinnabari Balf. Phytochemistry 2002, 61, 967–970. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, J.L.; Yang, C.R. Cochinchinenin-a new chalcone dimer from the Chinese Dragon’s blood. Acta Pharm. Sin. 2001, 36, 200–204. [Google Scholar]
- Zhou, Z.H.; Wang, J.L.; Yang, C.R. Chemical constituents of Sanguis Draxonis made in China. Chin. Tradit. Herb. Drugs 2001, 32, 484–486. [Google Scholar]
- Fischer, H.; Machen, T.E.; Widdicombe, J.H.; Carlson, T.J.; King, S.R.; Chow, J.W.; Illek, B. A novel extract SB-300 from the stem bark latex of Croton lechleri inhibits CFTR-mediated chloride secretion in human colonic epithelial cells. J. Ethnopharmacol. 2004, 93, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Gupta, R.K. Bioprotective properties of Dragon’s blood resin: In vitro evaluation of antioxidant activity and antimicrobial activity. Bmc Complementary Altern. Med. 2011, 11, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco-Topete, R.; Sierra-Madero, J.; Cano-Dominguez, C.; Kershenovich, J.; Ortiz- Pedroza, G.; Vazquez-Valls, E.; Garcia-Cosio, C.; Soria-Cordoba, A.; Armendariz, A.M.; Teran-Toledo, X. Safety and efficacy of Virend® for topical treatment of genital and anal herpes simplex lesions in patients with AIDS. Antiviriral Res. 1997, 35, 91–103. [Google Scholar] [CrossRef]
- Wang, H.; Liu, J.; Wu, J.; Mei, W.L. Flavonoids from Dracaena cambodiana. Chem. Nat. Compd. 2011, 47, 624–626. [Google Scholar] [CrossRef]
- Chen, Y.L.; Chen, S.; Liu, X.M. Experimental study on analgesic effect and block of nerve conduction induced by dragon’s blood. Lishizhen Med. Mater. Med. Res. 2010, 21, 2446–2447. [Google Scholar] [CrossRef]
- Machala, M.; Kubinova, R.; Horavova, P.; Suchy, V. Chemoprotective potentials of Homoisoflavonoids and Chalcones of Dracaena cinnabari: Modulations of Drug-metabolizing Ensymes and Antioxidant Activity. Phytother. Res. 2001, 15, 114–118. [Google Scholar] [CrossRef]
- Al-Awthan, Y.S.; Zarga, M.A.; Abdalla, S. Flavonoids content of Dracaena cinnabari resin and effects of the aqueous extract on isolated smooth muscle preparations, perfused heart, blood pressure and diuresis in the rat. Jordan J. Pharm. Sci. 2010, 3, 8–17. [Google Scholar]
- Eichholz, D.E.; Jones, W.H.S.; Rackham, H. Pliny: Natural History; with an English Translation in Ten Volumes; Jones, W.H.S., Rackham, H., Eds.; Harvard University Press: Cambridge, MA, USA, 1961; pp. 115–116. [Google Scholar]
- Casson, L. The Periplus of the Erythraean Sea; Princeton University Press: Princeton, NJ, USA, 1989; p. 169. [Google Scholar]
- Strauch, I. Foreign Sailors on Socotra: The Inscriptions and Drawings from the Cave HOQ.; Hempen: Bremen, Germany, 2012. [Google Scholar]
- Lev, E.; Amar, Z. Practical materia medica of the medieval eastern Mediterranean according to the Cairo Genizah; Brill: Leiden, Germany, 2008; p. 400. [Google Scholar]
- Al-Mujāwir, I. A Traveller in Thirteenth-century Arabia, Ibn al-Mujāwir’s Tārīkh al-Mustabir; Translated from Oscar Löfgren’s Arabic text and edited with revisions and annotations by G. Rex Smith; The Hakluyt Society: London, UK, 2008; p. 2264. [Google Scholar]
- Hirth, F.; Rockhill, W. Chau Ju-Kua, his work on the Chinese and Arab Trade in the Twelfth and Thirteenth Centuries, entitled ‘Chu-fan-chï’. St.; Imperial Academy of Sciences: Petersburg, Russia, 1911; pp. 130–132. [Google Scholar]
- Yāqūt, B.; Abd, A.R. Jacut’s Geographisches Wörterbuch (Kitāb mu’jam al-buldān); Wüstenfeld, F., Brockhaus, F.A., Eds.; Deutsche Morgenlandische Gesellschaf: Leipzig, Germany, 1866; p. 6237. [Google Scholar]
- Roe, T. The Embassy of Sir Thomas Roe to India, 1615–1619. (2 volumes); Foster, W., Ed.; The Hakluyt Society: London, UK, 1967; pp. 29–37. [Google Scholar]
- Foster, W. The English Factories in India 1665–1667. (12 vols.); Clarendon Press: Oxford, UK, 1970; pp. 1906–1927. [Google Scholar]
- Hunter, F.M.; Sealey, C.W.H. An Account of the Arab Tribes in the Vicinity of Aden; Darf Publishers: London, UK, 1986; pp. 366–367. [Google Scholar]
- Brown, G.H.H. Social and Economic Conditions and Possible Development of Socotra. Unpublished report. 1966; 29. [Google Scholar]
- Wellsted, J.R. Memoir on the Island of Socotra. J. R. Geogr. Soc. 1835, 5, 129–229. [Google Scholar] [CrossRef]
- Bent, T. Southern Arabia; Smith, Elder and Co.: London, UK, 1900; pp. 351–352. [Google Scholar]
- Botting, D. The Oxford University Expedition to Socotra. Geogr. J. 1958, 124, 200–207. [Google Scholar] [CrossRef]
- Shinnie, P.L. Socotra. Antiquity 1960, 34/134, 100–110. [Google Scholar] [CrossRef]
- Doe., D.B. Socotra: An Archaeological Reconnaissance in 1967; Coconut Grove: Miami, Florida, USA, 1970. [Google Scholar]
- Doe, D.B. Socotra Island of Tranquillity; Immel: London, UK, 1992. [Google Scholar]
- Naumkin, V.V.; Sedov, A.V. Monuments of Socotra. Topoi 1993, 70, 569–623. [Google Scholar] [CrossRef]
- Weeks, L.; Morris, M.; McCall, B.; Al-Zubairy, K. A recent archaeological survey on Soqotra. Report on the Preliminary Expedition Season,5 January–2 February 2001. Arab. Archaeol. Epigr. 2002, 13, 95–125. [Google Scholar] [CrossRef]
- Jansen van Rensburg, J.; Hopper, K. Incense and imagery: Mapping agricultural and water management systems on the island of Socotra, Yemen. Proc. Semin. Arab. Stud. 2017, 47, 129–138. [Google Scholar]
- Al-Fatimi, M. Ethnobotanical survey of Dracaena cinnabari and investigation of the pharmacognostical properties, antifungal and antioxidant activity of its resin. Plants 2018, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Černý, V.; Pereira, L.; Kujanová, M.; Vašíková, A.; Hájek, M.; Morris, M.; Mulligan, J.C. Out of Arabia—The Settlement of Island Soqotra as Revealed by Mitochondrial and Y Chromosome Genetic Diversity. Am. J. Phys. Anthropol. 2009, 138, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Elnoby, S.K.; Abdelraouf, A. Moustafa; Samira, R. Mansour. Impact of climate change on the endangered Nubian dragon tree (Dracaena ombet) in the South Eastern of Egypt. Catrina 2017, 16, 23–28. [Google Scholar] [CrossRef]
- 182. Abrha Tesfay Mehari. Ethnobotanical study of Dess’a forest, north-eastern escarpment of Ethiopia, with emphasis on use and management of forest resources by the local people. Master’s Thesis, Addis Ababa University, Addis Ababa, Ethiopia, 2008.
- Miller, A.G.; Morris, M. Plants of Dhofar, the Southern Region of Oman: Traditional, Economic and Medicinal uses, 1st ed.; Office of the Adviser for Conservation of the Environment, Diwan of Royal Court Sultanate of Oman Publishing: Sultanate of Oman, 1988; p. 16. [Google Scholar]
- Mies, B. On the comparison of the flora and vegetation of the island groups of Socotra and Macaronesia. Bol. Do Mus. Munic. Do Funchal (História Nat.) 1995, 4, 455–471. [Google Scholar]
- Kim, S.-C.; McGowen, M.R.; Lubinsky, P.; Barber, J.C.; Mort, M.E.; Santos-Guerra, A. Timing and tempo of early and successive adaptive radiations in Macaronesia. PLoS ONE 2008, 3, e2139. [Google Scholar] [CrossRef]
- Cropper, T. The weather and climate of Macaronesia: Past, present and future. Weather 2013, 68, 300–307. [Google Scholar] [CrossRef]
- Marzol, V.M.; Sánchez, J.L.; Yanes, A. Meteorological patterns and fog water collection in Morocco and the Canary Islands. Erdkunde 2011, 65, 291–303. [Google Scholar] [CrossRef]
- Médail, F.; Quézel, P. The phytogeographical signifikance of S. W. Morocco compared to the Canary Islands. Plant Ecol. 1999, 140, 221–244. [Google Scholar] [CrossRef]
- Beyhl, F.E. The Emta tree (Euphorbia arbuscula Balf. fil.), a succulent tree of dracoid habitus from the island of Soqotra (Dicotyledones: Tricococcae: Euphorbiaceae). In Proceedings of the First International Symposium on Soqotra Island: Present and Future; Dumont, H.J., Ed.; United Nations Publications: New York, NY, USA, 1996; pp. 115–123. [Google Scholar]
- Costa, J.C.; Neto, C.; Aguiar, C.; Capelo, J.; Espírito Santo, M.D.; Honrado, J.; Pinto-Gomes, C.; Monteiro-Henriques, T.; Sequeira, M.; Lousã, M. Vascular plant communities in Portugal (Continental, Azores and Madeira). Glob. Geobot. 2012, 2, 1–180. [Google Scholar] [CrossRef]
- Olehowski, C.; Naumann, S.; Fischer, D.; Siegmund, A. Geo-ecological spatial pattern analysis of the island of Fogo (Cape Verde). Glob. Planet. Chang. 2008, 64, 188–197. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Lousã, M.; Costa, J.C.; Duarte, M.C. Geobotanical survey of Cabo Verde Islands (West Africa). Int. J. Geobot. Res. 2017, 7, 1–103. [Google Scholar] [CrossRef]
- Kassas, M. The mist oasis of Erkwit, Sudan. J. Ecol. 1954, 44, 180–194. [Google Scholar] [CrossRef]
- Kürschner, H.; Hein, P.; Kilian, N.; Hubaishan, M.A. Diversity and zonation of the forests and woodlands of the mountains of northern Socotra, Yemen. Englera 2006, 28, 11–55. [Google Scholar] [CrossRef]
- De Sanctis, M.; Adeeb, A.; Farcomeni, A.; Patriarca, C.H.; Saed, A.; Attorre, F. Classification and distribution patterns of plant communities on Socotra Island, Yemen. Appl. Veg. Sci. 2013, 16, 148–165. [Google Scholar] [CrossRef] [Green Version]
- Habrová, H.; Buček, A. Overview of biotope types of Socotra Island. J. Lansdscape Ecol. 2013, 6, 60–83. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M.; Abdel-Khalik, K.N. Floristic Diversity and Phytogeography of the Gebel Elba National Park, South-East Egypt. Turk. J. Bot. 2006, 30, 121–136. [Google Scholar]
- Aynekulu, E.; Aerts, R.; Moonen, P.; Denich, M.; Gebrehiwot, K.; Vagen, T.G.; Mekuria, W.; Boehmer, H.J. Altitudinal variation and conservation priorities of vegetation along the Great Rift Valley escarpment, northern Ethiopia. Biodivers. Conserv. 2012, 21, 2691–2707. [Google Scholar] [CrossRef]
- Mwachala, G. Systematics and Ecology of Dracaena L. (Ruscaceae) in Central, East and Southern Africa. Ph.D. Thesis, University of Koblenz-Landau, Mainz, Germany, 2005. [Google Scholar]
- Van Breugel, P.; Friis, I.; Demissew, S. The transitional semi-evergreen bushland in Ethiopia: Characterization and mapping of its distribution using predictive modelling. Appl. Veg. Sci. 2016, 19, 355–367. [Google Scholar] [CrossRef]
- Friis, I.; Gilbert, M.G.; van Breugel, P.; Weber, O.; Demissew, S. Kalanchoe hypseloleuce (Crassulaceae), a new species from eastern Ethiopia, with notes on its habitat. Kew Bull. 2017, 72, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H. Forest vegetation of Xishuangbanna, south China. For. Stud. China 2006, 8, 1–58. [Google Scholar] [CrossRef]
- Zheng, D.J.; Li, H.W.; Yun, Y.; Zhang, L.; Zhang, Z.L. Investigation on the habitat and natural regeneration capacity of Dracaena cambodiana. J. Trop. Subtrop. Bot. 2010, 18, 627–632. [Google Scholar]
- Geekiyanage, N.; Goodale, U.M.; Cao, K.; Kitajima, K. Plant ecology of tropical and subtropical karst ecosystems. Biotropica 2019, 1–15. [Google Scholar] [CrossRef]
- Almeida Pérez, R.S. Dracaena tamaranae Marrero Rodr., R.S. Almeida & M. González-Martín. In Atlas y Libro Rojo de la Flora Vascular Amenazada de España, 2nd ed.; Bañares, A., Blanca, G., Güemes, J., Moreno, J.C., Ortiz, S., Eds.; Publicaciones de O.A.P.N.: Madrid, Spain, 2004; pp. 226–227. [Google Scholar]
- Almeida Pérez, R.S. Dracaena draco (L.) L. In Atlas y Libro Rojo de la Flora Vascular Amenazada de España, 2nd. ed.; Bañares, A., Blanca, G., Güemes, J., Moreno, J.C., Ortiz, S., Eds.; Publicaciones de O.A.P.N.: Madrid, Spain, 2004; pp. 680–681. [Google Scholar]
- Adolt, R.; Maděra, P.; Abraham, J.; Čupa, P.; Svrátek, M.; Matula, R.; Šebesta, J.; Čermák, M.; Volařík, D.; Koutecký, T.; et al. Field survey of Dracaena cinnabari populations in Firmihin, Socotra island: Methodology and preliminary results. J. Landsc. Ecol. 2013, 6, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Vetaas, O.R. Spatial and temporal vegetation changes along a moisture gradient in Northeastern Sudan. Biotropica 1993, 25, 164–175. [Google Scholar] [CrossRef]
- Van Rampellbergh, M.; Fleitmann, D.; Verheydeno, S.; Cheng, H.; Edwards, L.; De Geest, P.; De Vleeschouwer, D.; Burns, J.S.; Matter, A.; Claeys, P.; et al. Mid- to late holocene Indian ocean monsoon variability recorded in four speleothems from Socotra island, Yemen. Quat. Sci. Rev. 2013, 65, 129–142. [Google Scholar] [CrossRef]
- Marrero, A.; Almeida, R.S.; Roca, A. Diáspora y rescate: Acciones para la conservación del Drago de Gran Canaria, Dracaena tamaranae; Poster, V., Ed.; Congreso de Biología de la Conservación de Plantas: Balearic Islands, Spain, 2011. [Google Scholar]
- Chen, X.; Zhang, Z.; Yang, L.; Yun, Y.; Chen, J.; Zheng, D. Effects of light intensity on phenotypic plasticity and survival strategy of Dracaena cambodiana seedlings. J. Trop. Subtrop. Bot. 2018, 26, 150–156. [Google Scholar] [CrossRef]
- Moreno, J.C. (Ed.) Lista Roja 2008 de la flora vascular española, 1st ed.; Dirección General de Medio Natural y Política Forestal: Madrid, Español, 2008; p. 86. [Google Scholar]
- Galus, A.; Bouket, A.C.H.; Belbahri, L. In vitro propagation and acclimatization of Dragon Tree (Dracaena draco). Horticulturae 2019, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Al Hosni, A.; Oliver, I.; Al Jabri, Y.; Al Saidi, A.; Al Rawahi, A.; Al Hinai, H. Ex situ conservation of Dracaena serrulata in Dhofar province, southern Oman. Acta Hortic. 2018, 1190. [Google Scholar] [CrossRef]
- Schweiger, A.H.; Irl, S.D.H.; Svenning, J.C.H.; Higgins, S.I. Dynamic management needs for long-lived, sporadically recruiting plant species in human-dominated landscapes. Plants People Planet 2020, 1–15. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Description |
|---|---|
| Seedling | This stage is characterized by height increase being realized through leaf elongation. The completion of leaf elongation defines the end of this stage. In natural condition this stage lasts minimally five years, probably more. |
| Early juvenile stage | The number of leaves increases, and height growth is realized by addition of new leaves to the rosette. The appearance of stem formation defines the end of this stage. this stage typically lasts 5 to 10 years, but can be as short as 3 years and as long as 15 years |
| Medium juvenile stage | The onset of this stage is defined by the first appearance of the stem, and height increase is realized by stem growth. The end of this stage is defined when the stem height reaches 1.3 m. The stem diameter strongly depends on the number of leaves, thus the future stem diameter of adult trees is determined by the vitality of the plant in the early and medium juvenile stages. The end of this stage occurs when the tree is about 100 years. |
| Late juvenile stage | The stem height is more than 1.3 m and its end is defined by the onset of first flowering. This stage lasts between 100 and 150 or more years. |
| Species | Height [m] |
|---|---|
| Dracaena draco subsp. ajgal | 20 [4] |
| Dracaena draco subsp. draco | 12 [4], 21 [115], 22 [116] |
| Dracaena draco subsp. caboverdeana | 8 [7] |
| Dracaena tamaranae | 10 [4] |
| Dracaena cinnabari | 10 [4], 12 [111] |
| Dracaena ombet subsp. schizantha | 9 [4] |
| Dracaena ombet subsp. ombet | 8 [4] |
| Dracaena serrulata | 8 [4] |
| Dracaena americana | 10-12 (18) [12] |
| Dracaena jayniana | 5-8 [9] |
| Dracaena kaweesakii | 3-6 (12) [10] |
| Dracaena cambodiana | 3-4 (10) [11] |
| Species | Taxonomy | Evolution | Distribution | Anatomy, morphology | Ontogenetic cycle | Water relations | Resin, ethnobotany | Ecology | Populations | Threat, conservation |
|---|---|---|---|---|---|---|---|---|---|---|
| Dracaena spp. | 1, 2, 3, 33, 34, 35, 36, 37, 38, 42 | 13, 33, 35, 42, 45, 46, 97 | 85 | 4, 39, 40, 41, 42, 43, 44, 71, 72, 77, 78, 84, 86, 87, 88, 89, 90, 91, 94, 96, 97 | 86 | 97 | ||||
| D.cinnabari | 15, 27 | 24, 27, 55, 56 | 4, 43, 44, 73, 81, 83, 87 | 14, 18, 111, 112, 113, 114 | 23, 83, 124, 125, 126, 127 | 27, 107, 129, 130, 131, 134, 135, 145, 146, 148, 149, 153, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 176, 178, 179 | 4, 23, 24, 28, 29, 111, 184, 194, 195, 196 | 14, 18, 21, 25, 207 | 14, 18, 19, 21, 25, 26, 32, 113, 207, 215 | |
| D.serrulata | 17 | 4, 17, 57, 58 | 4, 43, 44 | 4 | 131, 183 | 4, 17, 31 | 17 | 17, 214 | ||
| D.ombet subsp. ombet | 6 | 4, 6, 20, 59, 60, 61, 62, 63 | 4, 43, 44 | 4 | 131, 182 | 4, 20, 193, 197, 198 | 20, 63, 181 | 20, 63, 181, 208 | ||
| D.ombet subsp. schizantha | 6 | 3, 6, 61, 64, 65 | 4, 43, 44, 87 | 4 | 129 | 4, 61 | ||||
| D. ellenbeckiana | 6 | 6 | 44, 87 | 61, 199, 200, 201 | ||||||
| D.draco subsp. draco | 51, 52, 53, 54 | 4, 43, 44, 70, 74, 75, 76, 82, 83, 87, 93, 95, 99, 106 | 4, 76, 115, 116, 117, 118, 119, 120 | 83, 109, 125, 126, 128 | 107, 110, 129, 131, 132, 138, 139, 148 | 4, 30, 106, 184, 185, 188, 190 | 206, 212, 213 | |||
| D.draco subsp. ajgal | 8 | 8 | 131, 143 | 4, 8, 188 | ||||||
| D.draco subsp. caboverdeana | 7 | 7, 50 | 7, 79 | 7, 191, 192 | ||||||
| D.tamaranae | 4, 5 | 5 | 4, 16 | 4, 43 | 132 | 4, 16 | 16 | 16, 205, 210, 212 | ||
| D.cochinchinensis | 47 | 68, 69 | 68, 102 | 69, 102, 107, 108, 129, 133, 136, 137, 140, 141, 144, 147, 150, 151 | 202 | |||||
| D.cambodiana | 47, 48, 49, 105 | 11, 49, 66, 67 | 44, 87, 103, 105 | 10, 11 | 103, 105, 129, 155 | 11, 49, 203 | 11 | 11, 211 | ||
| D. kaweesakii | 10 | 10 | 43 | 10 | ||||||
| D. jayniana | 9 | 9 | 43 | 9 | ||||||
| D. yuccifolia | 10 | |||||||||
| D.americana | 12 | 12 | 87 | 12 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maděra, P.; Forrest, A.; Hanáček, P.; Vahalík, P.; Gebauer, R.; Plichta, R.; Jupa, R.; Van Rensburg, J.J.; Morris, M.; Nadezhdina, N.; et al. What We Know and What We Do Not Know about Dragon Trees? Forests 2020, 11, 236. https://doi.org/10.3390/f11020236
Maděra P, Forrest A, Hanáček P, Vahalík P, Gebauer R, Plichta R, Jupa R, Van Rensburg JJ, Morris M, Nadezhdina N, et al. What We Know and What We Do Not Know about Dragon Trees? Forests. 2020; 11(2):236. https://doi.org/10.3390/f11020236
Chicago/Turabian StyleMaděra, Petr, Alan Forrest, Pavel Hanáček, Petr Vahalík, Roman Gebauer, Roman Plichta, Radek Jupa, Julian Jansen Van Rensburg, Miranda Morris, Nadezhda Nadezhdina, and et al. 2020. "What We Know and What We Do Not Know about Dragon Trees?" Forests 11, no. 2: 236. https://doi.org/10.3390/f11020236