Aboveground Carbon Content and Storage in Mature Scots Pine Stands of Different Densities



Abstract
:1. Introduction
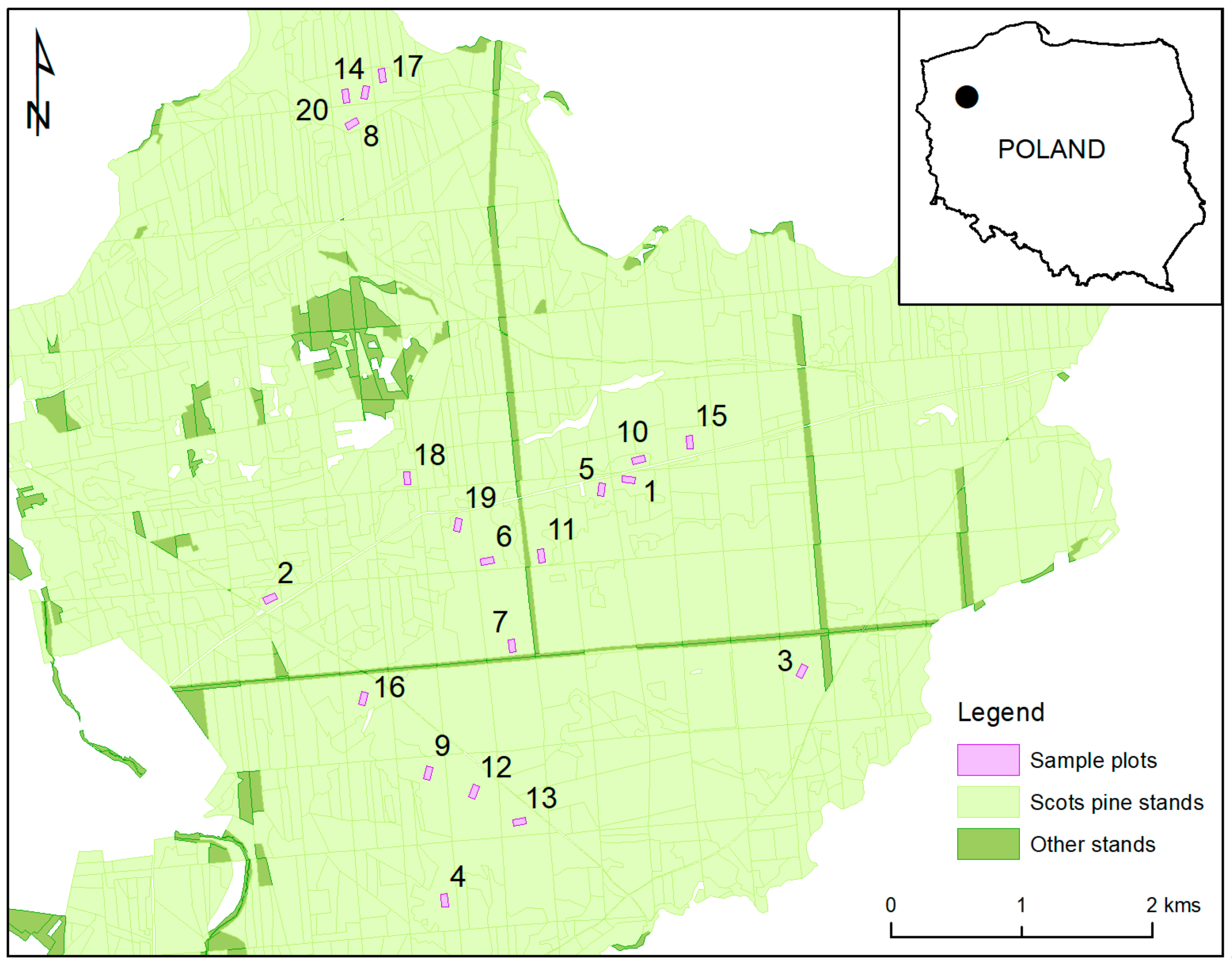
2. Materials and Methods
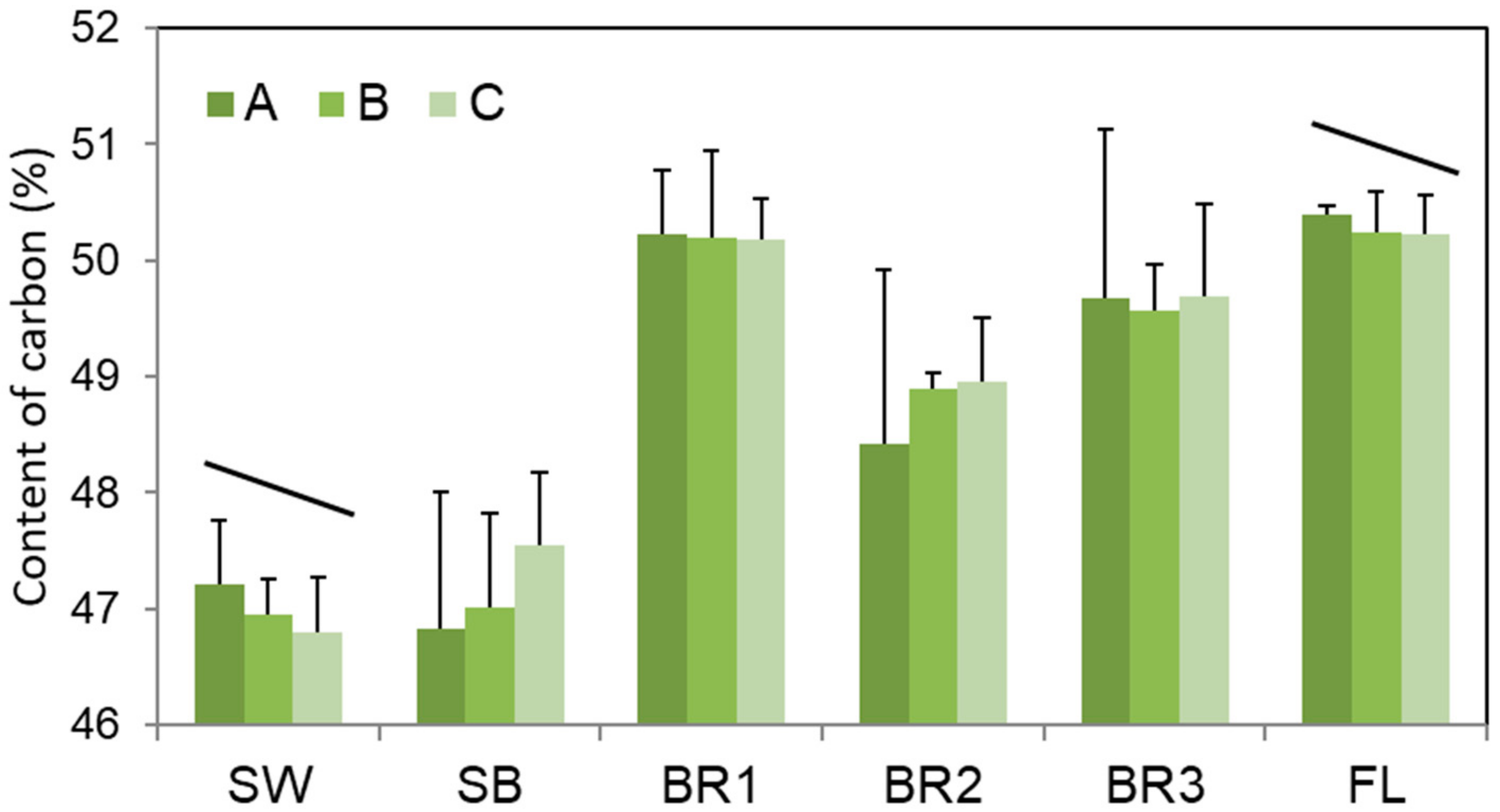
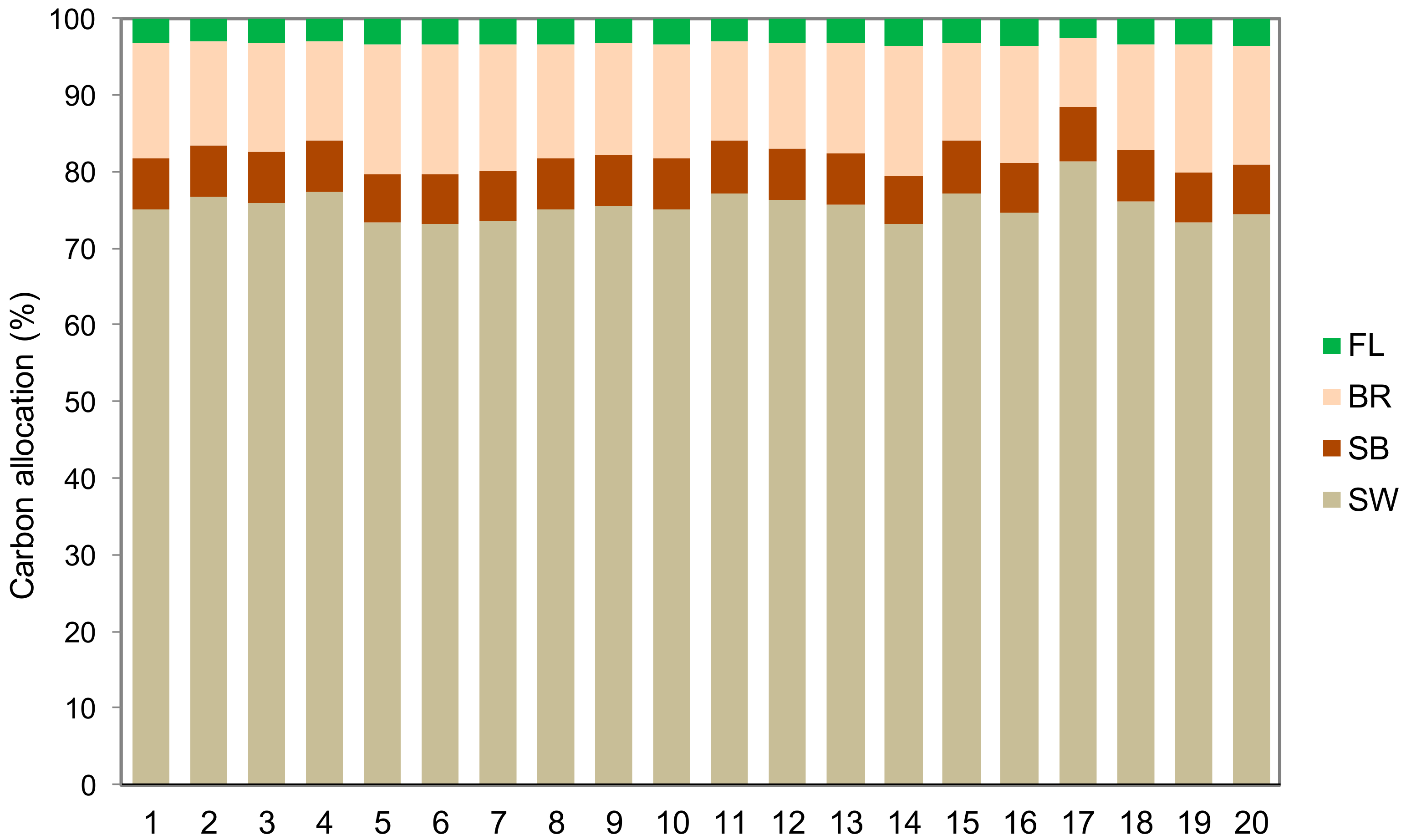
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Oberthür, S.; Ott, H.E. The Kyoto Protocol: International Climate Policy for the 21st Century; Springer Science & Business Media: Berlin, Germany, 1999; p. 360. [Google Scholar]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Lamlom, S.H.; Savidge, R.A. A reassessment of carbon content in wood: Variation within and between 41 North American species. Biomass Bioenergy 2003, 25, 381–388. [Google Scholar] [CrossRef]
- Malmsheimer, R.W.; Bowyer, J.L.; Fried, J.S.; Gee, E.; Izlar, R.L.; Miner, R.A.; Munn, I.A.; Oneil, E.; Stewart, W.C. Managing forests because carbon matters: Integrating energy, products, and land management policy. J. For. 2011, 109, 7–51. [Google Scholar] [CrossRef]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Sedjo, R.A. Forests: A tool to moderate global warming. Environ. Sci. Policy Sustain. Dev. 1989, 31, 14–20. [Google Scholar] [CrossRef]
- Dewar, R.C.; Cannell, M.G. Carbon sequestration in the trees, products and soils of forest plantations: An analysis using UK examples. Tree Physiol. 1992, 11, 49–71. [Google Scholar] [CrossRef]
- Hollinger, D.Y.; Maclaren, J.P.; Beets, P.N.; Turland, J. Carbon sequestration by New Zealand’s Plantation forests. N. Z. J. For. Sci. 1993, 23, 194–208. [Google Scholar]
- Laiho, R.; Laine, J. Tree stand biomass and carbon content in an age sequence of drained pine mires in southern Finland. For. Ecol. Manag. 1997, 93, 161–169. [Google Scholar] [CrossRef]
- Bert, D.; Danjon, F. Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Thomas, S.C.; Malczewski, G. Wood carbon content of tree species in Eastern China: Interspecific variability and the importance of the volatile fraction. J. Environ. Manag. 2007, 85, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.C.; Martin, A.R. Carbon content of tree tissues: A synthesis. Forests 2012, 3, 332–352. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.R.; Doraisami, M.; Thomas, S.C. Global patterns in wood carbon concentration across the world’s trees and forests. Nat. Geosci. 2018, 11, 915–920. [Google Scholar] [CrossRef]
- Tahvonen, O.; Pihlainen, S.; Niinimäki, S. On the economics of optimal timber production in boreal Scots pine stands. Can. J. For. Res. 2013, 43, 719–730. [Google Scholar] [CrossRef]
- Janssens, I.A.; Sampson, D.A.; Cermak, J.; Meiresonne, L.; Riguzzi, F.; Overloop, S.; Ceulemans, R. Above- and belowground phytomass and carbon storage in a Belgian Scots pine stand. Ann. For. Sci. 1999, 56, 81–90. [Google Scholar] [CrossRef]
- GUS. Główny Urząd Statystyczny. Leśnictwo; Informacje i Opracowania Statystyczne: Warszawa, Poland, 2017. [Google Scholar]
- Valinger, E.; Elfving, B.; Mörling, T. Twelve-year growth response of Scots pine to thinning and nitrogen fertilisation. For. Ecol. Manag. 2000, 134, 45–53. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and long-term changes in increment and stem form of Scots pine trees. For. Ecol. Manag. 2004, 203, 21–34. [Google Scholar] [CrossRef]
- Giuggiola, A.; Bugmann, H.; Zingg, A.; Dobbertin, M.; Rigling, A. Reduction of stand density increases drought resistance in xeric Scots pine forests. For. Ecol. Manag. 2013, 310, 827–835. [Google Scholar] [CrossRef]
- Karu, H.; Szava-Kovats, R.; Pensa, M.; Kull, O. Carbon sequestration in a chronosequence of Scots pine stands in a reclaimed opencast oil shale mine. Can. J. For. Res. 2009, 39, 1507–1517. [Google Scholar] [CrossRef]
- Vanninen, P.; Mäkelä, A. Carbon budget for Scots pine trees: Effects of size, competition and site fertility on growth allocation and production. Tree Physiol. 2005, 25, 17–30. [Google Scholar] [CrossRef]
- Armolaitis, K.; Varnagirytė-Kabašinskienė, I.; Stupak, I.; Kukkola, M.; Mikšys, V.; Wojcik, J. Carbon and nutrients of Scots pine stands on sandy soils in Lithuania in relation to bioenergy sustainability. Biomass Bioenergy 2013, 54, 250–259. [Google Scholar] [CrossRef]
- Skonieczna, J.; Małek, S.; Polowy, K.; Węgiel, A. Element content of Scots pine (Pinus sylvestris L.) stands of different densities. Drewno 2014, 57, 77–87. [Google Scholar] [CrossRef]
- Bembenek, M.; Giefing, D.E.; Jelonek, T.; Karaszewski, Z.; Kruszyk, R.; Tomczak, A.; Woszczyk, M.; Mederski, P.S. Carbon content in juvenile and mature wood of Scots pine (Pinus sylvestris L.). Balt. For. 2015, 21, 279–284. [Google Scholar]
- Tolunay, D. Carbon concentrations of tree components, forest floor and understorey in young Pinus sylvestris stands in north-western Turkey. Scand. J. For. Res. 2009, 24, 394–402. [Google Scholar] [CrossRef]
- Durkaya, A.; Durkaya, B.; Makineci, E.; Orhan, I. Aboveground biomass and carbon storage relationship of Turkish pines. Fresenius Environ. Bull. 2015, 24, 3573–3583. [Google Scholar]
- Lee, J.; Tolunay, D.; Makineci, E.; Çömez, A.; Son, Y.M.; Kim, R.; Son, Y. Estimating the age-dependent changes in carbon stocks of Scots pine (Pinus sylvestris L.) stands in Turkey. Ann. For. Sci. 2016, 73, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Erkan, N.; Güner, S.T. Determination of carbon concentration of tree components for Scotch pine forests in Turkmen Mountain (Eskisehir, Kutahya) Region. Forestist 2018, 68, 87–92. [Google Scholar] [CrossRef]
- Mjöfors, K.; Strömgren, M.; Nohrstedt, H.-Ö.; Johansson, M.-B.; Gärdenäs, A.I. Indications that site preparation increases forest ecosystem carbon stocks in the long term. Scand. J. For. Res. 2017, 32, 717–725. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Ruiz-Peinado, R.; Modrego, P.; Alonso, R.; Montero, G. Forest thinning impact on carbon stock and soil condition in Southern European populations of P. sylvestris L. For. Ecol. Manag. 2015, 357, 259–267. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; Montero, G.; del Río, M. Carbon stocks in a Scots pine afforestation under different thinning intensities management. Mitig. Adapt. Strateg. Glob. Chang. 2016, 21, 1059–1072. [Google Scholar] [CrossRef]
- Pohjola, J.; Valsta, L. Carbon credits and management of Scots pine and Norway spruce stands in Finland. For. Policy Econ. 2007, 9, 789–798. [Google Scholar] [CrossRef]
- Goetz, R.U.; Hritonenko, N.; Mur, R.J.; Xabadia, A.; Yatsenko, Y. Forest management and carbon sequestration in size-structured forests: The case of Pinus sylvestris in Spain. For. Sci. 2010, 56, 242–256. [Google Scholar] [CrossRef]
- Goetz, R.U.; Hritonenko, N.; Mur, R.; Xabadia, À.; Yatsenko, Y. Forest management for timber and carbon sequestration in the presence of climate change: The case of Pinus sylvestris. Ecol. Econ. 2013, 88, 86–96. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Díaz-Pinés, E.; Barbeito, I.; Sánchez-González, M.; Montes, F.; Rubio, A.; Cañellas, I. Temporal carbon dynamics over the rotation period of two alternative management systems in Mediterranean mountain Scots pine forests. For. Ecol. Manag. 2015, 348, 186–195. [Google Scholar] [CrossRef]
- Węgiel, A.; Małek, S.; Bielinis, E.; Grebner, D.L.; Polowy, K.; Skonieczna, J. Determination of elements removal in different harvesting scenarios of Scots pine (Pinus sylvestris L.) stands. Scand. J. For. Res. 2018, 33, 261–270. [Google Scholar] [CrossRef]
- Węgiel, A.; Bembenek, M.; Łacka, A.; Mederski, P.S. Relationship between stand density and value of timber assortments: A case study for Scots pine stands in north-western Poland. N. Z. J. For. Sci. 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Picard, N.; Saint-André, L.; Henry, M. Manual for Building Tree Volume and Biomass Allometric Equations: From Field Measurement to Prediction; FAO: Rome, Italy; CIRAD: Montpellier, France, 2012; p. 215. [Google Scholar]
- Bruchwald, A. New empirical formula for determination of volume of Scots pine stands. Folia For. Pol. 1996, 38, 5–10. [Google Scholar]
- Kojola, S.; Ahtikoski, A.; Hökkä, H.; Penttilä, T. Profitability of alternative management regimes in Scots pine stands on drained peatlands. Eur. J. For. Res. 2012, 131, 413–426. [Google Scholar] [CrossRef]
- Curtis, R.O.; Marshall, D.D.; Bell, J.F. LOGS: A pioneering example of silvicultural research in coast Douglas-fir. J. For. 1997, 95, 19–25. [Google Scholar]
- Kuliešis, A.; Saladis, J.; Kuliešis, A.A. Development and productivity of young Scots pine stands by regulating density. Balt. For. 2010, 16, 235–246. [Google Scholar]
- Nilsson, U.; Agestam, E.; Ekö, P.-M.; Elfving, B.; Fahlvik, N.; Johansson, U.; Karlsson, K.; Lundmark, T.; Wallentin, C. Thinning of Scots pine and Norway spruce monocultures in Sweden. Stud. For. Suec. 2010, 219, 1–46. [Google Scholar]
- Zhang, J.; Oliver, W.W.; Ritchie, M.W. Effect of stand densities on stand dynamics in white fir (Abies concolor) forests in northeast California, USA. For. Ecol. Manag. 2007, 244, 50–59. [Google Scholar] [CrossRef]
- Del Río, M.; Calama, R.; Cañellas, I.; Roig, S.; Montero, G. Thinning intensity and growth response in SW-European Scots pine stands. Ann. For. Sci. 2008, 65. [Google Scholar] [CrossRef] [Green Version]
- Gizachew, B.; Brunner, A. Density-growth relationships in thinned and unthinned Norway spruce and Scots pine stands in Norway. Scand. J. For. Res. 2011, 26, 543–554. [Google Scholar] [CrossRef]
- Routa, J.; Kellomäki, S.; Strandman, H. Effects of forest management on total biomass production and CO2 emissions from use of energy biomass of Norway spruce and Scots pine. Bioenergy Res. 2012, 5, 733–747. [Google Scholar] [CrossRef]
- Egnell, G.; Ulvcrona, K.A. Stand productivity following whole-tree harvesting in early thinning of Scots pine stands in Sweden. For. Ecol. Manag. 2015, 340, 40–45. [Google Scholar] [CrossRef]
- Moulinier, J.; Brais, S.; Harvey, B.D.; Koubaa, A. Response of boreal Jack pine (Pinus banksiana Lamb.) stands to a gradient of commercial thinning intensities, with and without N fertilization. Forests 2015, 6, 2678–2702. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Zhao, S.Q.; Liu, S.; Oeding, J. A meta-analysis on the impacts of partial cutting on forest structure and carbon storage. Biogeosciences 2013, 10, 3691–3703. [Google Scholar] [CrossRef] [Green Version]
- Güner, Ş.T.; Makineci, E. Determination of annual organic carbon sequestration in soil and forest floor of Scots pine forests on the Türkmen Mountain (Eskişehir, Kütahya). J. Fac. For. Istanb. Univ. 2017, 67, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Alegria, C. Simulation of silvicultural scenarios and economic efficiency for maritime pine (Pinus pinaster Aiton) wood-oriented management in centre inland of Portugal. For. Syst. 2011, 20, 361–378. [Google Scholar] [CrossRef] [Green Version]
- Litton, C.M.; Ryan, M.G.; Knight, D.H. Effects of tree density and stand age on carbon allocation patterns in postfire lodgepole pine. Ecol. Appl. 2004, 14, 460–475. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Stand Number | Age (Years) | Stand Density (tree·ha−1) | Mean DBH ± SD (cm) | Mean Height ± SD (m) | Basal Area (m2·ha−1) | Volume (m3·ha−1) |
|---|---|---|---|---|---|---|
| 1 * | 82 | 476 | 28.2 ± 4.5 | 22.9 ± 1.7 | 30.5 | 325 |
| 2 | 87 | 518 | 27.5 ± 4.9 | 23.6 ± 2.0 | 31.8 | 350 |
| 3 | 82 | 534 | 25.9 ± 4.3 | 22.4 ± 1.5 | 28.4 | 298 |
| 4 | 82 | 564 | 28.0 ± 4.5 | 24.5 ± 1.4 | 35.5 | 401 |
| 5 * | 82 | 594 | 25.7 ± 4.7 | 20.8 ± 1.4 | 31.5 | 306 |
| 6 | 82 | 596 | 27.1 ± 5.2 | 21.6 ± 1.4 | 35.7 | 358 |
| 7 | 82 | 618 | 24.6 ± 4.0 | 20.4 ± 1.5 | 30.2 | 288 |
| 8 | 82 | 632 | 24.0 ± 4.7 | 21.1 ± 1.4 | 29.6 | 293 |
| 9 | 82 | 632 | 25.6 ± 4.7 | 22.0 ± 1.4 | 33.6 | 347 |
| 10 | 82 | 634 | 24.6 ± 4.6 | 21.4 ± 1.6 | 31.0 | 311 |
| 11 | 82 | 640 | 26.0 ± 4.7 | 23.4 ± 1.7 | 35.1 | 385 |
| 12 | 82 | 644 | 26.4 ± 4.4 | 23.0 ± 1.4 | 35.9 | 384 |
| 13 | 82 | 648 | 25.9 ± 5.2 | 22.3 ± 1.7 | 35.4 | 373 |
| 14 * | 82 | 674 | 23.6 ± 4.3 | 19.6 ± 1.5 | 30.1 | 280 |
| 15 | 82 | 682 | 24.7 ± 4.9 | 22.8 ± 1.7 | 34.0 | 363 |
| 16 | 82 | 708 | 21.8 ± 4.2 | 19.5 ± 2.2 | 27.3 | 250 |
| 17 | 82 | 720 | 22.3 ± 4.3 | 24.7 ± 3.8 | 29.1 | 345 |
| 18 | 82 | 756 | 21.7 ± 4.4 | 20.5 ± 2.0 | 28.9 | 281 |
| 19 * | 82 | 758 | 23.9 ± 5.3 | 20.1 ± 2.0 | 35.7 | 340 |
| 20 * | 82 | 836 | 21.8 ± 4.0 | 19.3 ± 1.7 | 31.5 | 291 |
| Tree Part | Equation | Equation Coefficients | R2 | RSE | ||
|---|---|---|---|---|---|---|
| a | b | c | ||||
| Stem (wood + bark) | a · (D2 · H2)b | 0.012996 | 0.761234 | 0.9357 | 22.187 | |
| Thick branches | a · Db · Hc | 0.048094 | 3.333163 | −1.53455 | 0.7281 | 8.9224 |
| Thin branches | a · (D2 · H2)b | 0.001054 | 0.879284 | 0.6744 | 1.5290 | |
| Dead branches | a · Db · Hc | 0.023518 | 3.569452 | 1.87147 | 0.7457 | 3.3543 |
| Needles | a · (D)b | 0.011092 | 2.012406 | 0.7499 | 1.9823 | |
| Tree Part | Aboveground Biomass (Mg·ha−1) | Carbon Content (%) | |||||
|---|---|---|---|---|---|---|---|
| Mean | SD | Proportion | Mean | SD | Min | Max | |
| Stem wood | 116.0 | 16.2 | 76.3% | 47.0 | 0.5 | 46.3 | 48.1 |
| Stem bark | 10.1 | 1.4 | 6.6% | 47.1 | 1.0 | 45.8 | 49.1 |
| Thick branches | 13.3 | 1.9 | 8.7% | 48.8 | 1.0 | 45.5 | 50.0 |
| Thin branches | 3.0 | 0.3 | 2.0% | 50.2 | 0.6 | 49.2 | 51.3 |
| Dead branches | 5.0 | 0.8 | 3.3% | 49.6 | 1.0 | 48.4 | 52.5 |
| Needles | 4.7 | 0.4 | 3.1% | 50.3 | 0.3 | 49.7 | 50.9 |
| Total | 152.1 | 100% | |||||
| Stand Number | Stand Density (tree·ha−1) | Above Ground Biomass (Mg·ha−1) | Carbon Stock (Mg·ha−1) | ||||
|---|---|---|---|---|---|---|---|
| SW | SB | BR | FL | Total | |||
| 1 | 476 | 147.6 | 52.5 | 4.6 | 10.6 | 2.3 | 69.9 |
| 2 | 518 | 160.2 | 58.1 | 5.1 | 10.3 | 2.3 | 75.8 |
| 3 | 534 | 137.3 | 49.4 | 4.3 | 9.2 | 2.1 | 65.1 |
| 4 | 564 | 183.7 | 67.1 | 5.9 | 11.3 | 2.6 | 86.9 |
| 5 | 594 | 140.7 | 48.9 | 4.3 | 11.2 | 2.3 | 66.8 |
| 6 | 596 | 162.9 | 56.5 | 5.0 | 13.1 | 2.6 | 77.2 |
| 7 | 618 | 133.1 | 46.6 | 4.1 | 10.4 | 2.2 | 63.2 |
| 8 | 632 | 135.8 | 48.4 | 4.2 | 9.6 | 2.2 | 64.5 |
| 9 | 632 | 159.7 | 57.1 | 5.0 | 11.1 | 2.5 | 75.7 |
| 10 | 634 | 143.4 | 51.1 | 4.5 | 10.2 | 2.3 | 68.0 |
| 11 | 640 | 177.6 | 65.0 | 5.7 | 10.9 | 2.6 | 84.1 |
| 12 | 644 | 176.2 | 63.7 | 5.6 | 11.6 | 2.7 | 83.5 |
| 13 | 648 | 171.1 | 61.4 | 5.4 | 11.7 | 2.6 | 81.1 |
| 14 | 674 | 130.0 | 45.2 | 4.0 | 10.5 | 2.2 | 61.8 |
| 15 | 682 | 169.0 | 61.8 | 5.4 | 10.3 | 2.5 | 80.1 |
| 16 | 708 | 118.0 | 41.9 | 3.7 | 8.6 | 2.0 | 56.1 |
| 17 | 720 | 168.4 | 64.9 | 5.7 | 7.1 | 2.2 | 79.8 |
| 18 | 756 | 132.5 | 48.0 | 4.2 | 8.7 | 2.1 | 63.0 |
| 19 | 758 | 157.3 | 54.8 | 4.8 | 12.5 | 2.6 | 74.7 |
| 20 | 836 | 137.1 | 48.5 | 4.2 | 10.1 | 2.3 | 65.2 |
| Parameter | Mean DBH | Mean Height | Basal Area | Stand Volume | Above Ground Biomass | Stand Density |
|---|---|---|---|---|---|---|
| C stock in stem | 0.486 * | 0.850 *** | 0.713 *** | 0.961 *** | 0.986 *** | −0.160 |
| C stock in branches | 0.599 *** | −0.053 | 0.827 *** | 0.501 * | 0.413 | −0.223 |
| C stock in needles | 0.534 * | 0.321 | 0.999 *** | 0.846 *** | 0.809 *** | −0.045 |
| C stock in aboveground biomass | 0.011 | 0.789 *** | 0.815 *** | 0.991 *** | 1.000 *** | −0.184 |
| Percentage of C stock in stem | −0.019 | 0.790 *** | −0.026 | 0.447 * | 0.534 * | 0.005 |
| Percentage of C stock in branches | 0.061 | −0.762 *** | 0.051 | −0.416 | −0.505 * | −0.041 |
| Percentage of C stock in needles | −0.334 | −0.956 *** | −0.186 | −0.666 *** | −0.721 *** | 0.292 |
| Stand density | −0.860 *** | −0.508 * | −0.044 | −0.269 | −0.190 | − |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Węgiel, A.; Polowy, K. Aboveground Carbon Content and Storage in Mature Scots Pine Stands of Different Densities. Forests 2020, 11, 240. https://doi.org/10.3390/f11020240
Węgiel A, Polowy K. Aboveground Carbon Content and Storage in Mature Scots Pine Stands of Different Densities. Forests. 2020; 11(2):240. https://doi.org/10.3390/f11020240
Chicago/Turabian StyleWęgiel, Andrzej, and Krzysztof Polowy. 2020. "Aboveground Carbon Content and Storage in Mature Scots Pine Stands of Different Densities" Forests 11, no. 2: 240. https://doi.org/10.3390/f11020240