Flowering Phenology and Characteristics of Pollen Aeroparticles of Quercus Species in Korea

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
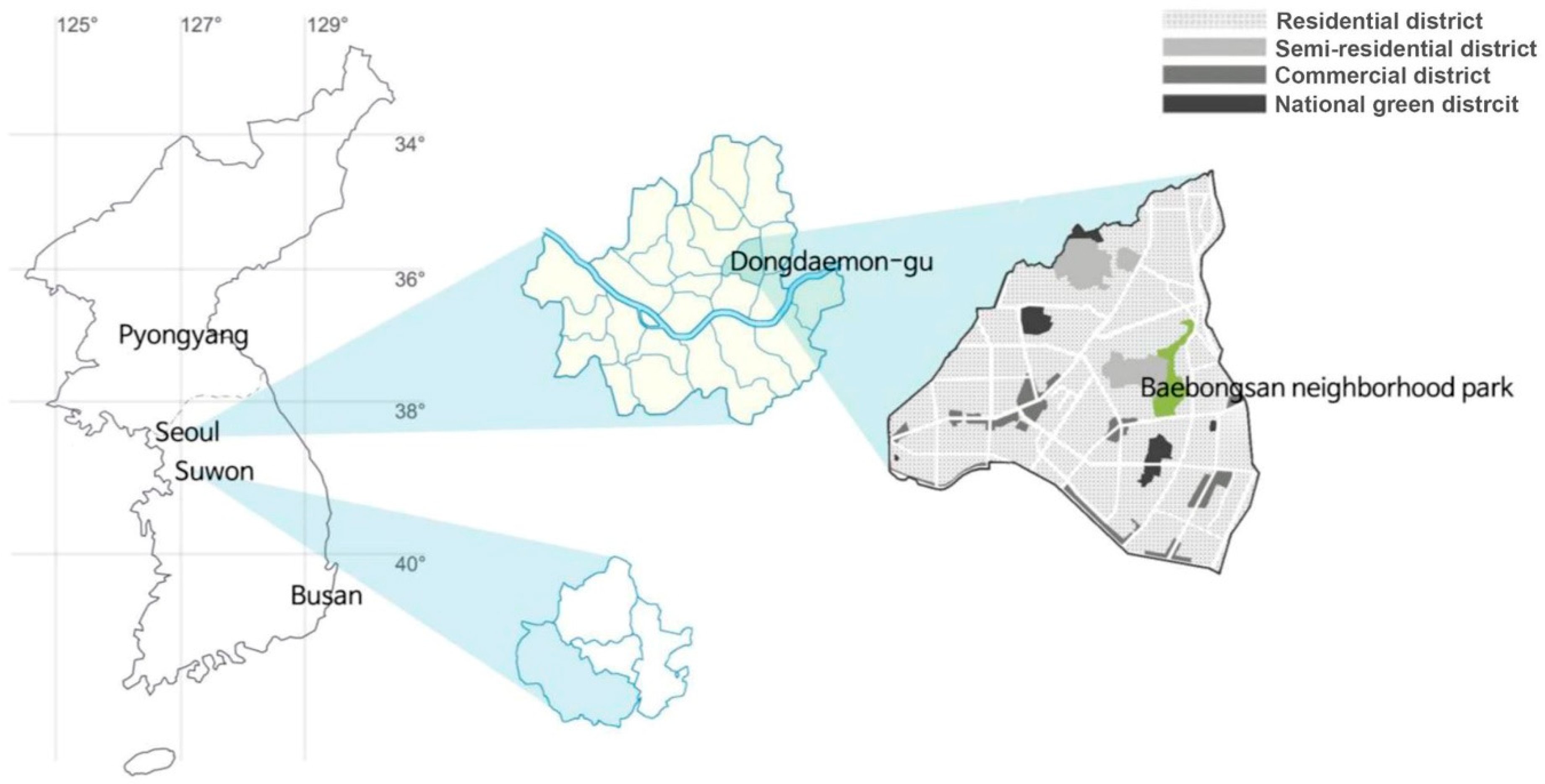
2.1. Study Site and Plant Material
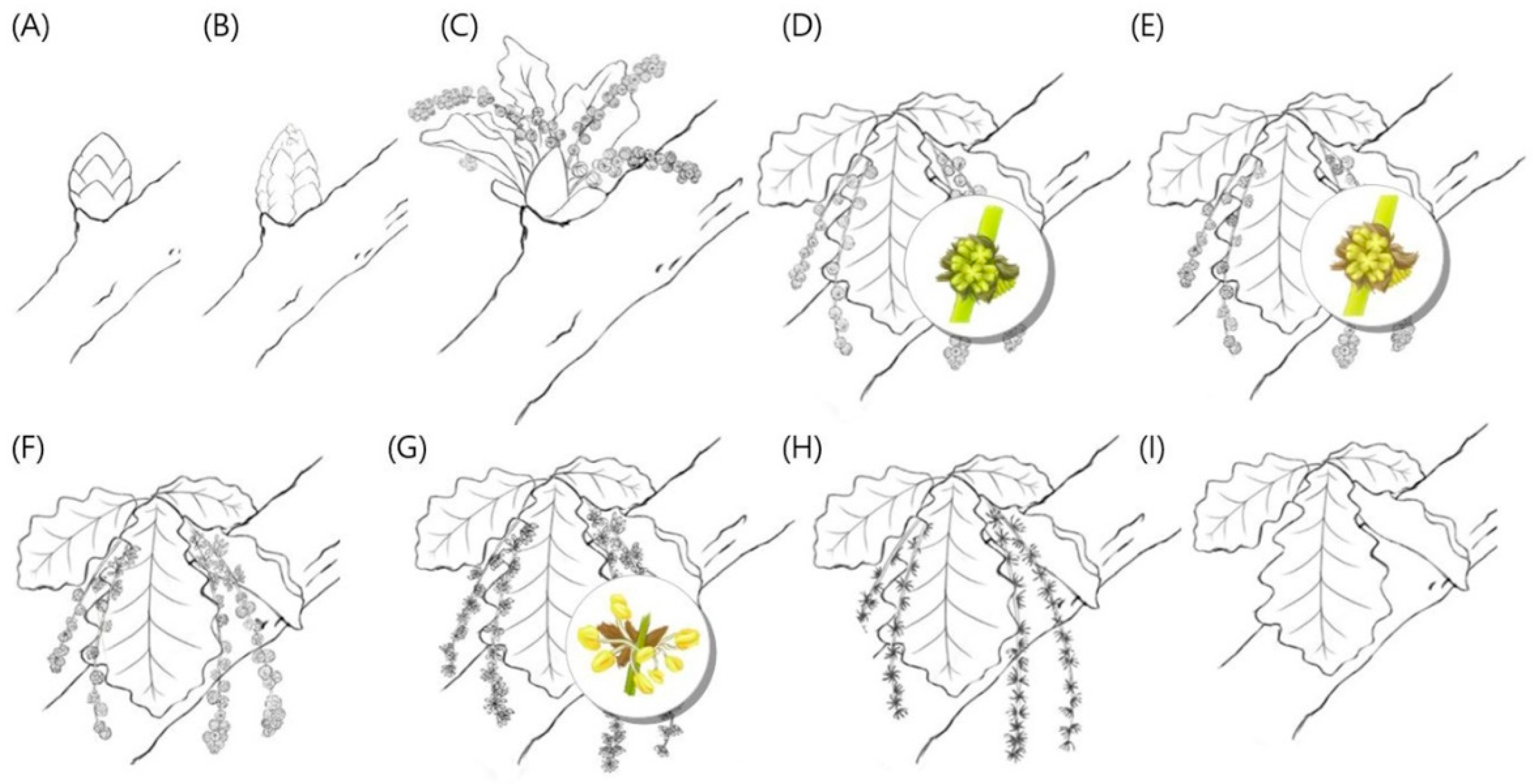
2.2. Flowering Phenology
2.3. Airborne Pollen
2.4. Size of Pollen Grains
2.5. Observation of Pollen Exine Structure
2.6. Soluble Protein Content
2.7. Statistical Analysis
3. Results
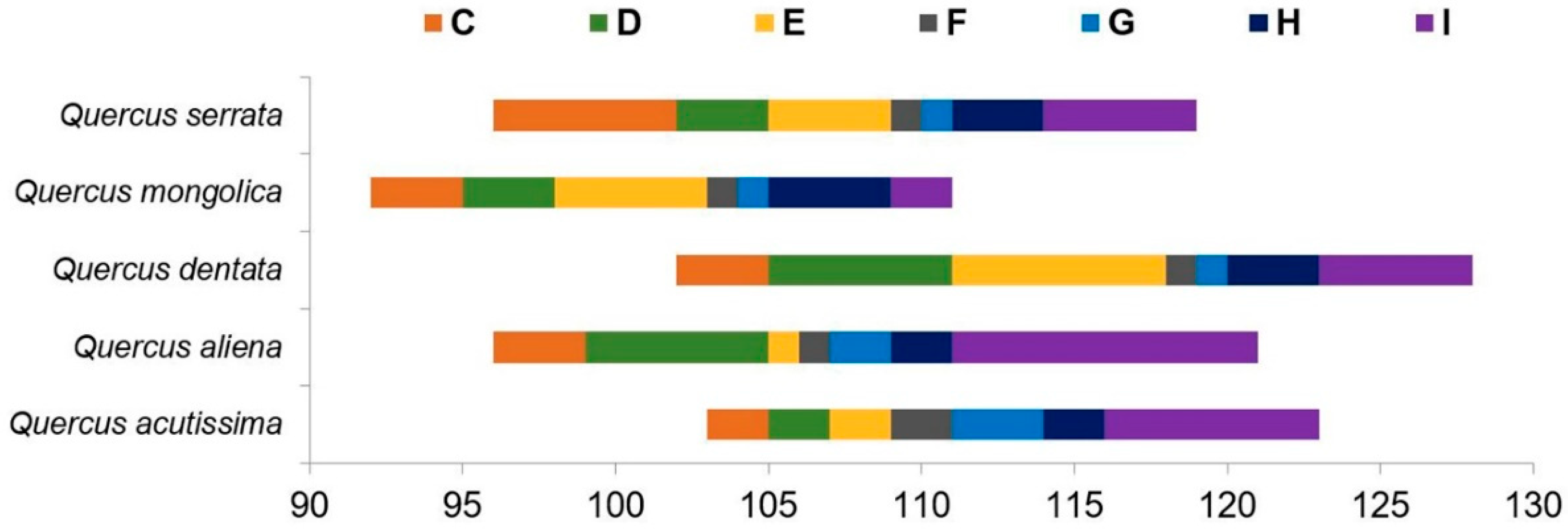
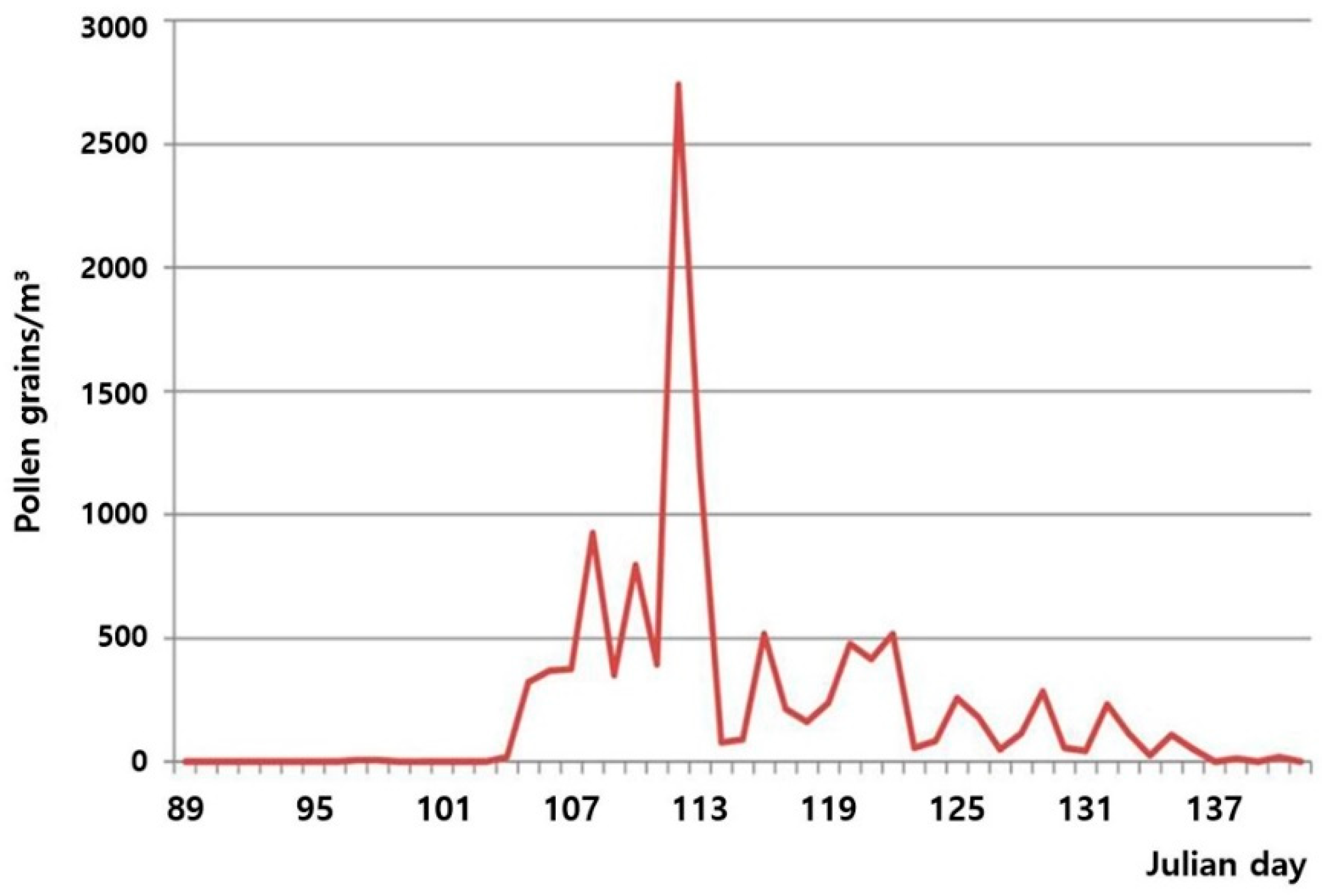
3.1. Flowering Phenology
3.2. Size of Pollen Grains
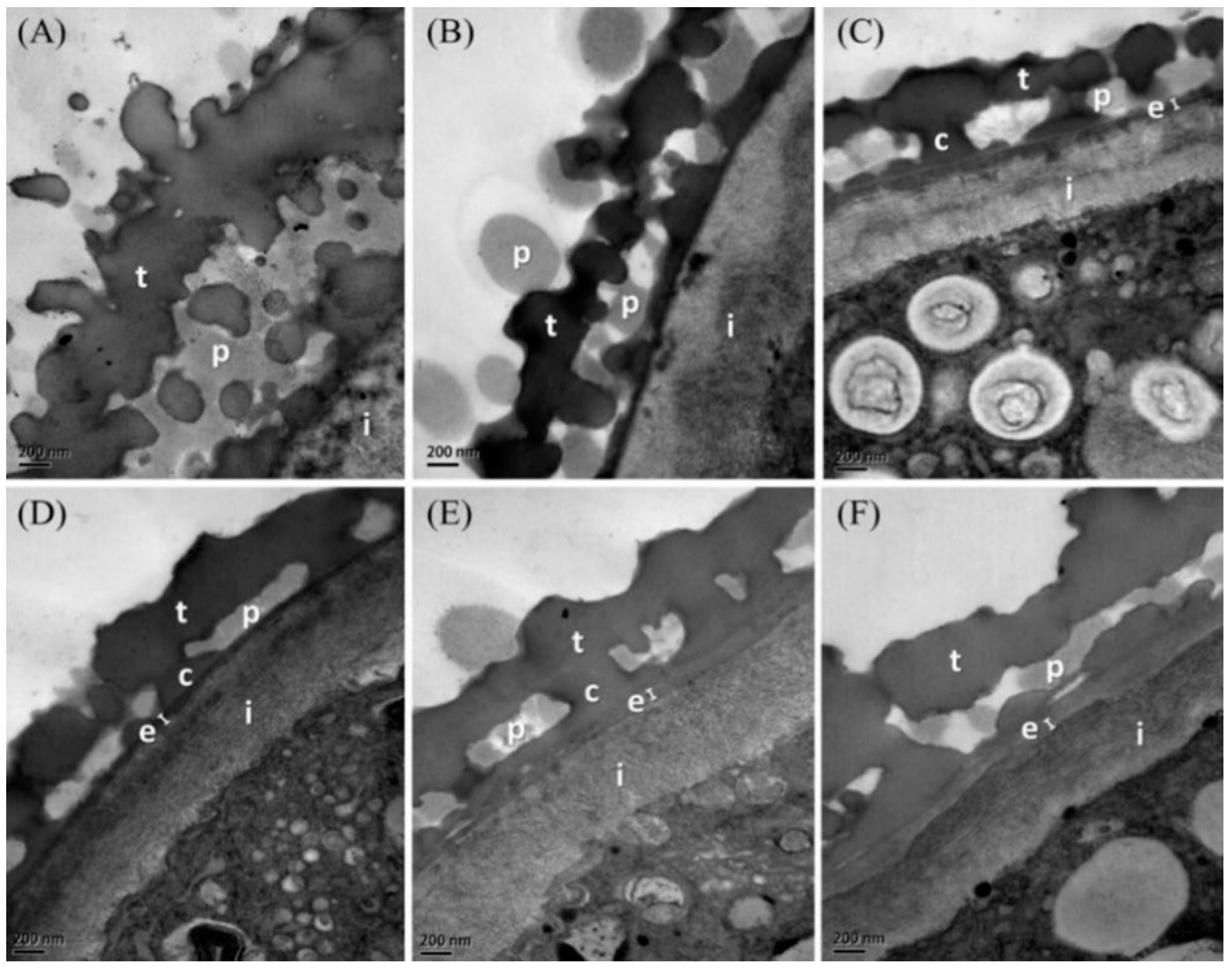
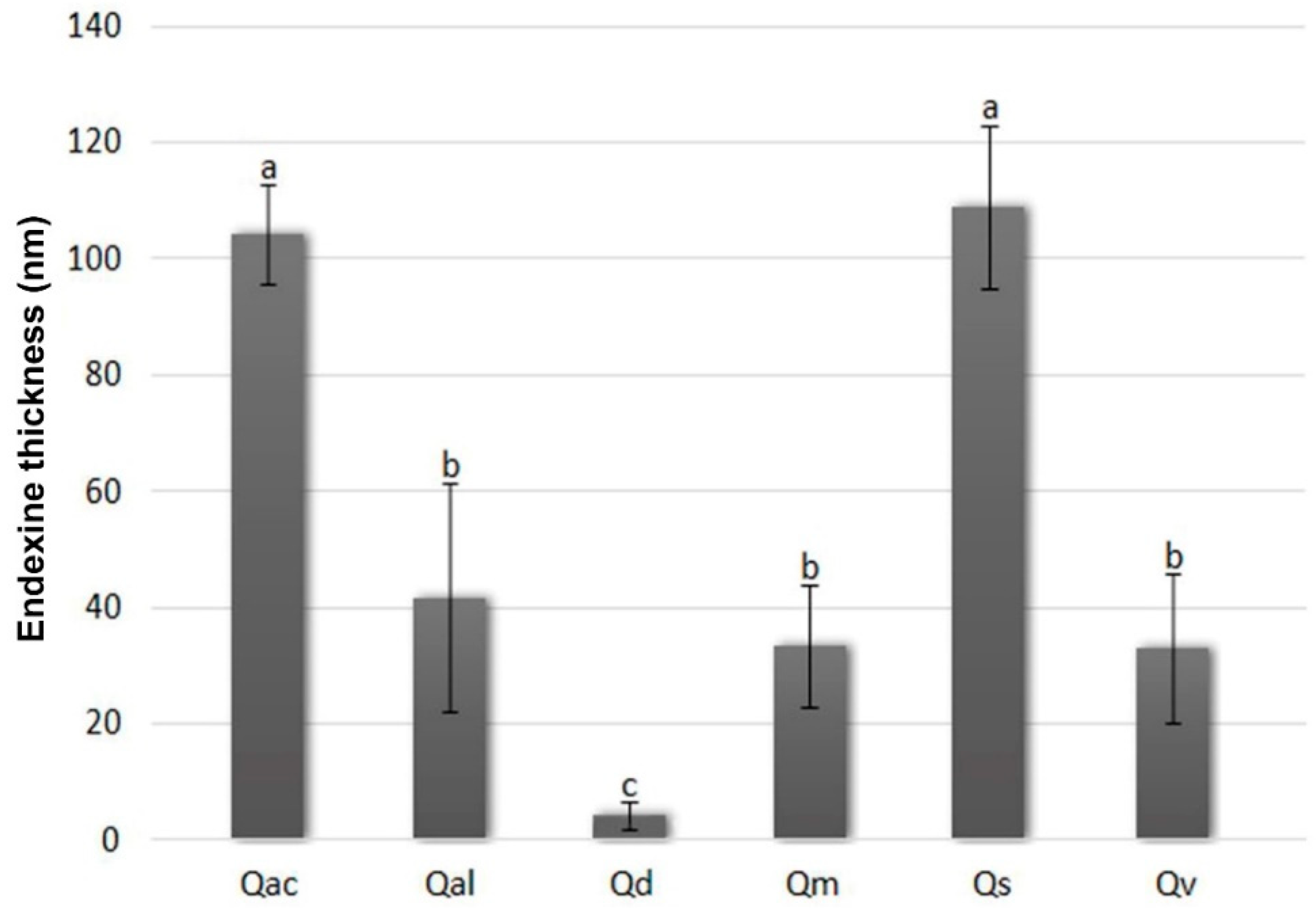
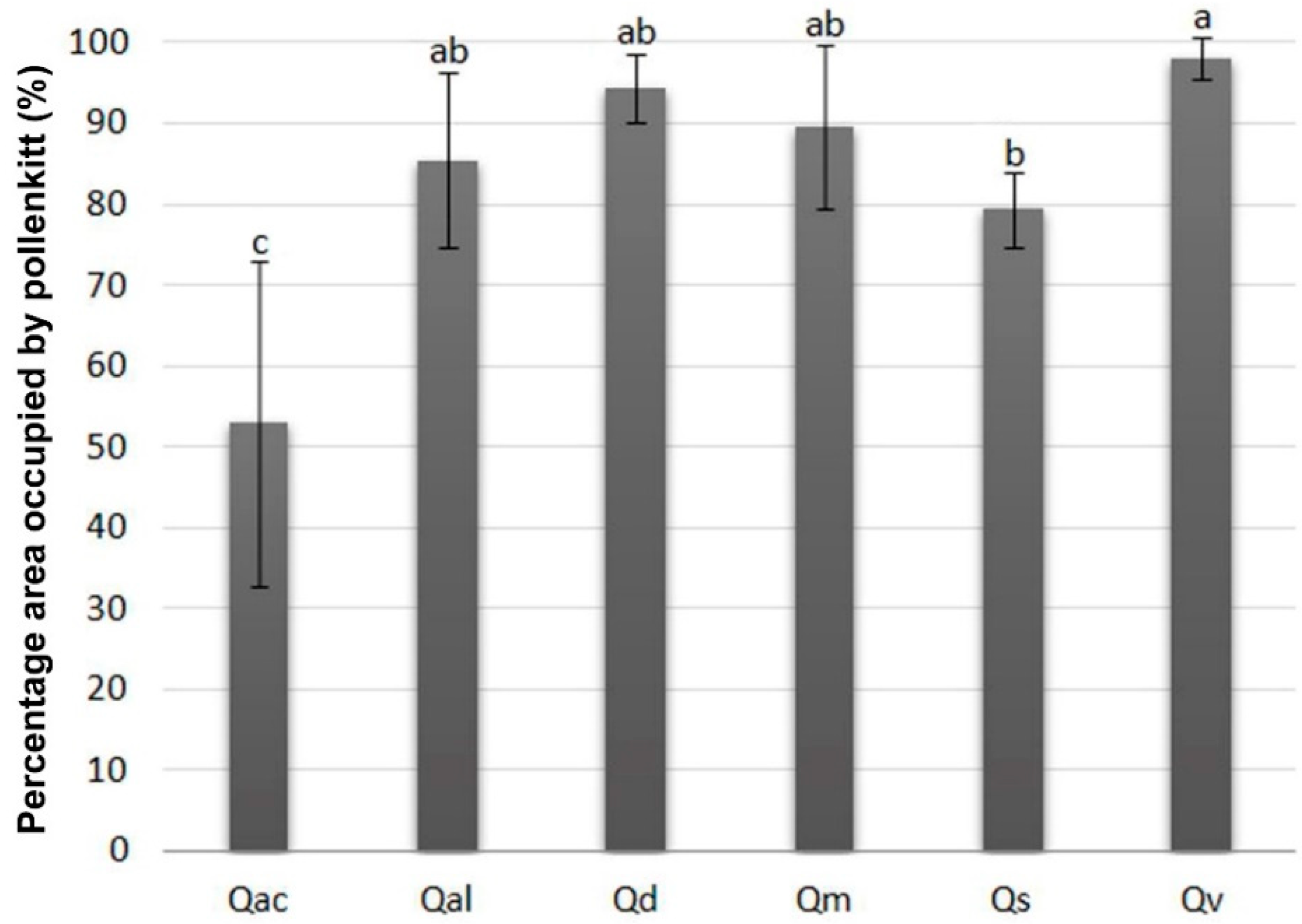
3.3. Specificity of Pollen Endexine Thickness
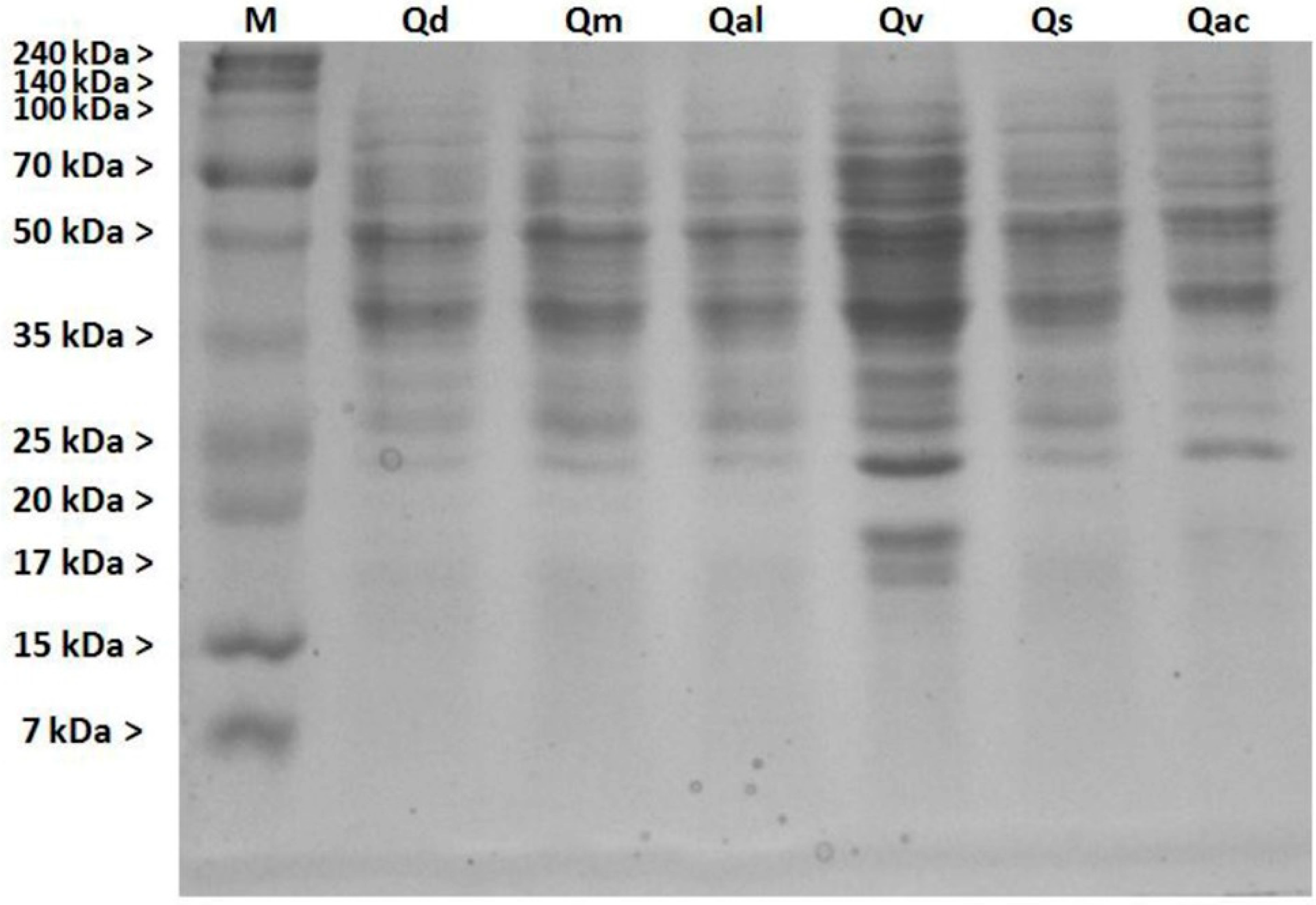
3.4. Soluble Protein Contents of Pollen
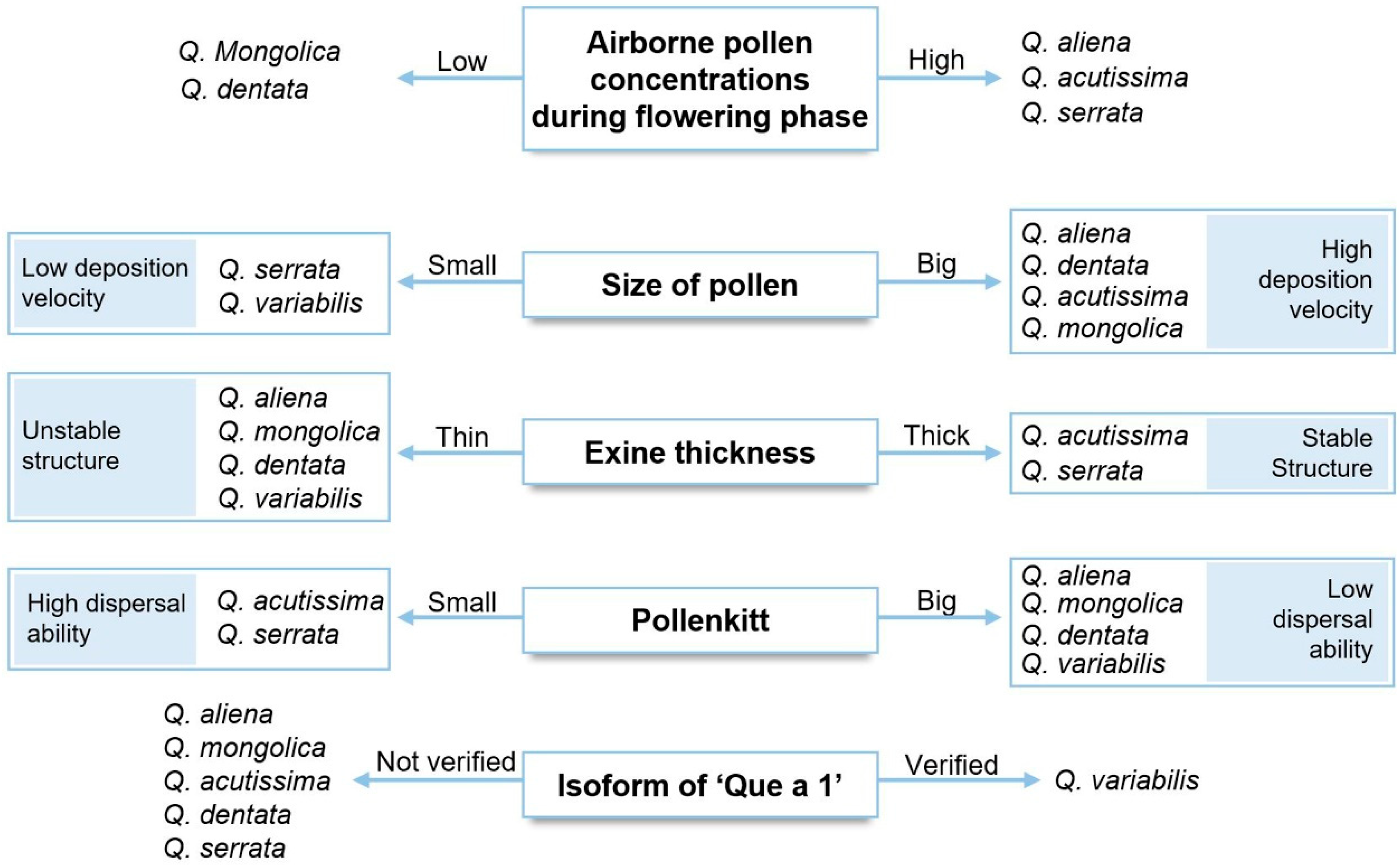
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Korea Forest Service. 2019 Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korea, 2019; p. 164. [Google Scholar]
- Pilar, C.D.; Gabriel, M.M. Phenological pattern of fifteen Mediterranean phanaerophytes from shape Quercus ilex communities of NE-Spain. Plant Ecol. 1998, 139, 103–112. [Google Scholar] [CrossRef]
- Boavida, L.C.; Silva, J.P.; Feijó, J.A. Sexual reproduction in the cork oak (Quercus suber L.). II. Crossing intra-and interspecific barriers. Sex. Plant Reprod. 2001, 14, 143–152. [Google Scholar] [CrossRef]
- Rocheta, M.; Sobral, R.; Magalhães, J.; Amorim, M.I.; Ribeiro, T.; Pinheiro, M.; Conceição Egas, C.; Leonor Morais-Cecílio, L.; Costa, M.M. Comparative transcriptomic analysis of male and female flowers of monoecious Quercus suber. Front. Plant Sci. 2014, 5, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.G. Ecological Management of Forests; Seoul National University Press: Seoul, Korea, 2012; pp. 179–193. [Google Scholar]
- Lee, C.S.; Cho, H.J.; Yi, H. Stand dynamics of introduced black locust (Robinia pseudoacacia L.) plantation under different disturbance regimes in Korea. For. Ecol. Manag. 2004, 189, 281–293. [Google Scholar] [CrossRef]
- Kim, H.S.; Lee, S.M.; Song, H.K. Actual vegetation distribution status and ecological succession in the Deogyusan National Park. Korean J. Environ. Ecol. 2011, 5, 37–46. [Google Scholar]
- Park, H.J.; Lim, H.S.; Park, K.H.; Lee, J.H.; Park, J.W.; Hong, C.S. Changes in allergen sensitization over the last 30 years in Korea respiratory allergic patients: A single-center. Allergy Asthma Immunol. Res. 2014, 6, 434–443. [Google Scholar] [CrossRef]
- Edlund, A.F.; Swanson, R.; Preuss, D. Pollen and stigma structure and function: The role of diversity in pollination. Plant Cell 2004, 16, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Diethart, B.; Sam, S.; Weber, M. Walls of allergenic pollen: Special reference to the endexine. Grana 2007, 46, 164–175. [Google Scholar] [CrossRef]
- Culley, T.M.; Weller, S.G.; Sakai, A.K. The evolution of wind pollination in angiosperms. Trends Ecol. Evol. 2002, 17, 361–369. [Google Scholar] [CrossRef]
- Petersen, A.; Dresselhaus, T.; Grobe, K.; Becker, W.M. Proteome analysis of maize pollen for allergy-relevant components. Proteomics 2006, 6, 6317–6325. [Google Scholar] [CrossRef]
- Lin, H.; Gomez, I.; Meredith, J.C. Pollenkitt wetting mechanism enables species-specific tunable pollen adhesion. Langmuir 2013, 29, 3012–3023. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Makra, L.; Harry, S.K.; Bruffaerts, N.; Hendrickx, M.; Coates, F.; Saarto, A.; Thibaudon, M.; Oliver, G.; Damialis, A.; et al. Temperature-related changes in airborne allergenic pollen abundance and seasonality across the northern hemisphere: A retrospective data analysis. Lancet Planet Health 2019, 3, e124–e131. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Liccardi, G.; Popov, T.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy 2007, 62, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Lake, I.R.; Jones, N.R.; Agnew, M.; Goodess, C.M.; Giorgi, F.; Hamaoui-Laguel, L.; Semenov, M.A.; Solomon, F.; Storkey, J.; Vautard, R.; et al. Climate change and future pollen allergy in Europe. Environ. Health Perspect. 2017, 125, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Singer, B.D.; Ziska, L.H.; Frenz, D.A.; Gebhard, D.E.; Straka, J.G. Increasing Amb a 1 content in common ragweed (Ambrosia artemisiifolia) pollen as a function of rising atmospheric CO2 concentration. Funct. Plant Biol. 2005, 32, 667–670. [Google Scholar] [CrossRef]
- Anenberg, S.C.; Weinberger, K.R.; Roman, H.; Neumann, J.E.; Crimmins, A.; Fann, N.; Martinich, J.; Kinney, P.L. Impacts of oak pollen on allergic asthma in the United States and potential influence of future climate change. GeoHealth 2017, 1, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Yoon, M.G.; Kim, M.; Jin, H.J.; Shin, Y.S.; Park, H.S. Identification of immunoglobulin E binding components of two major tree pollens, birch and alder. Allergy Asthma Respir. Dis. 2013, 1, 216–220. [Google Scholar] [CrossRef]
- Hong, C.S. Pollen allergy plants in Korea. Allergy Asthma Respir. Dis. 2015, 3, 237–252. [Google Scholar] [CrossRef]
- Heath, M.D.; Collis, J.; Batten, T.; Hutchings, J.W.; Swan, N.; Skinner, M.A. Molecular, proteomic and immunological parameters of allergens provide inclusion criteria for new candidates within established grass and tree homologous groups. World Allergy Organ. J. 2015, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann-Sommergruber, K. Pathogenesis-related (PR)-proteins identified as allergens. Biochem. Soc. Trans. 2002, 30, 930–935. [Google Scholar] [CrossRef] [Green Version]
- Park, K.J.; Kim, H.; Kim, K.R.; Oh, J.W.; Lee, S.Y.; Choi, Y.J. Characteristics of regional distribution of pollen concentration in Korean Peninsula. Korean J. Agric. For. Meteorol. 2008, 10, 167–176. [Google Scholar] [CrossRef]
- Smouse, P.E.; Dyer, R.J.; Westfall, R.D.; Sork, V.L. Two-generation analysis of pollen flow across a landscape. I. Male gamete heterogeneity among females. Evolution 2001, 55, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Garcıa-Mozo, H.; Galán, C.; Aira, M.J.; Belmonte, J.; de la Guardia, C.D.; Fernández, D.; Gutierrez, A.M.; Rodriguez, F.J.; Trigo, M.M.; Dominguez-Vilches, E. Modelling start of oak pollen season in different climatic zones in Spain. Agric. For. Meteorol. 2002, 110, 247–257. [Google Scholar] [CrossRef]
- Panahi, P.; Pourmajidian, M.R.; Pourhashemi, M. Pollen morphology of Quercus (subgenus Quercus, section Quercus) in Iran and its systematic implication. Acta Soc. Bot. Pol. 2012, 81, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Codina, R.; Lockey, R.F. Pollen used to produce allergen extracts. Ann. Allergy Asthma Immunol. 2017, 118, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Solomon, W.R.; Burge, H.R.; Boise, J.R.; Becker, M. Comparative particle recoveries by the retracting rotorod, rotoslide and burkard spore trap sampling in a compact array. Int. J. Biometeorol. 1980, 24, 107–116. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, J.H.; Park, K.H.; Kim, K.R.; Han, M.J.; Choe, H.; Oh, J.W.; Hong, C.S. A six-year study on the changes in airborne pollen counts and skin positivity rates in Korea: 2008–2013. Yonsei Med. J. 2016, 57, 714–720. [Google Scholar] [CrossRef] [Green Version]
- Wrońska-Pilarek, D.; Danielewicz, W.; Bocianowski, J.; Maliński, T.; Janyszek, M. Comparative pollen morphological analysis and its systematic implications on three European Oak (Quercus L., Fagaceae) species and their spontaneous hybrids. PLoS ONE 2016, 11, e0161762. [Google Scholar] [CrossRef]
- Morris, J.K. A formaldehyde glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 1965, 27, 137–139. [Google Scholar]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- de Souza, E.H.; Souza, F.V.D.; Rossi, M.L.; Brancalleão, N.; da Silva-Ledo, C.A.; Martinelli, A.P. Viability, storage and ultrastructure analysis of Aechmea bicolor (Bromeliaceae) pollen grains, an endemic species to the Atlantic forest. Euphytica 2015, 204, 13–28. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sousa, R.; Duque, L.; Duarte, A.J.; Gomes, C.R.; Ribeiro, H.; Cruz, A.; da Silva, J.C.G.E.; Abreu, I. In vitro exposure of Acer negundo pollen to atmospheric levels of SO2 and NO2: Effects on allergenicity and germination. Environ. Sci. Technol. 2012, 46, 2406–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harlow, E.D.; Lane, D. A Laboratory Manual New York: Cold Spring Harbor Laboratory; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1998; p. 579. [Google Scholar]
- Bogawski, P.; Borycka, K.; Grewling, Ł.; Kasprzyk, I. Detecting distant sources of airborne pollen for Poland: Integrating back-trajectory and dispersion modelling with a satellite-based phenology. Sci. Total Environ. 2019, 689, 109–125. [Google Scholar] [CrossRef]
- McInnes, R.N. Pollen, Allergens, and Human Health. In Oxford Research Encyclopedia of Environmental Science; Oxford University Press: Oxford, UK, 2019; pp. 1–38. [Google Scholar]
- Scialla, T.; Wanner, A. Fundamentals of Asthma Treatment. Pulmão RJ 2012, 21, 33–40. [Google Scholar]
- Lo, F.; Bitz, C.M.; Battisti, D.S.; Hess, J.J. Pollen calendars and maps of allergenic pollen in North America. Aerobiologia 2019, 35, 613–633. [Google Scholar] [CrossRef] [Green Version]
- McInnes, R.N.; Hemming, D.; Burgess, P.; Lyndsay, D.; Osborne, N.J.; Skjøth, C.A.; Thomas, S.; Vardoulakis, S. Mapping allergenic pollen vegetation in UK to study environmental exposure and human health. Sci. Total Environ. 2017, 599, 483–499. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.W.; Lee, H.B.; Kang, I.J.; Kim, S.W.; Park, K.S.; Kook, M.H.; Kim, B.S.; Baek, H.S.; Kim, J.H.; Lee, D.J.; et al. The revised edition of Korean calendar for allergenic pollens. Allergy Asthma Immunol. Res. 2012, 4, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Camacho, I.C. Airborne pollen in Funchal city, (Madeira Island, Portugal)-first pollinic calendar and allergic risk assessment. Ann. Agric. Environ. Med. 2015, 22, 608–613. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Casero, M.T.; Galán, C.; Domínguez-Vilches, E. Flowering phenology of Mediterranean Quercus species in different locations (Córdoba, SW Iberian Peninsula). Acta Bot. Malac. 2007, 32, 127–146. [Google Scholar]
- Aguilera, F.; Valenzuela, L.R. Study of the floral phenology of Olea europaea L. in Jaén province (SE Spain) and its relation with pollen emission. Aerobiologia 2009, 25, 217–225. [Google Scholar] [CrossRef]
- Chesnoiu, E.N.; Șofletea, N.; Curtu, A.L.; Toader, A.; Radu, R.; Enescu, M. Bud burst and flowering phenology in a mixed oak forest from Eastern Romania. Ann. For. Res. 2009, 52, 199–206. [Google Scholar]
- Choi, S.H.; Jung, I.Y.; Kim, D.Y.; Kim, Y.H.; Lee, J.H.; Oh, I.B.; Choi, K.R. Seasonal distribution of airborne pollen in Ulsan, Korea in 2009–2010. Korean J. Environ. Ecol. 2011, 34, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.K.; Kong, M.J.; Song, J.H.; Kim, S.Y.; Kim, J.S.; Jung, E.H.; Park, C.H.; Lee, B.Y.; Hong, S.P. Morphological characteristics of major airborne pollen in Korea peninsula. J. Species Res. 2015, 4, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.K.; Kim, K.R.; Cho, C.; Kim, M.; Choi, H.S.; Han, M.J.; Oh, I.B.; Kim, B.J. Development of a Oak Pollen Emission and Transport Modeling Framework in Korea. Atmosphere 2015, 25, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Celenk, S. Detection of reactive allergens in long-distance transported pollen grains: Evidence from Ambrosia. Atmos. Environ. 2019, 209, 212–219. [Google Scholar] [CrossRef]
- Aylor, D.E. Settling speed of corn (Zea mays) pollen. J. Aerosol. Sci. 2002, 33, 1601–1607. [Google Scholar] [CrossRef]
- Tseng, Y.T.; Kawashima, S.; Kobayashi, S.; Takeuchi, S.; Nakamura, K. Forecasting the seasonal pollen index by using a hidden Markov model combining meteorological and biological factors. Sci. Total Environ. 2020, 698, 134246. [Google Scholar] [CrossRef]
- Tseng, Y.T.; Kawashima, S. Applying a pollen forecast algorithm to the Swiss Alps clarifies the influence of topography on spatial representativeness of airborne pollen data. Atmos. Environ. 2019, 212, 153–162. [Google Scholar] [CrossRef]
- Williams, C.G. How meso-scale pollen dispersal and its gene flow shape gene conservation decisions. New For. 2017, 48, 217–224. [Google Scholar] [CrossRef]
- Oteros, J.; Bartusel, E.; Alessandrini, F.; Núñez, A.; Moreno, D.A.; Behrendt, H.; Schmidt-Weber, C.; Traidl-Hoffmann, C.; Buters, J. Artemisia pollen is the main vector for airborne endotoxin. J. Allergy Clin. Immunol. 2019, 143, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traidl-Hoffmann, C.; Kasche, A.; Menzel, A.; Jakob, T.; Thiel, M.; Ring, J.; Behrendt, H. Impact of pollen on human health: More than allergen carriers? Int. Arch. Allergy Immunol. 2003, 131, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Nakamura, S.; Lu, S.; Xiu, G.; Nakajima, D.; Suzuki, M.; Sakamoto, K.; Miwa, M. Release behavior of small sized daughter allergens from Cryptomeria japonica pollen grains during urban rainfall event. Aerobiologia 2012, 28, 71–81. [Google Scholar] [CrossRef]
- Dahl, Å. Pollen lipids can play a role in allergic airway inflammation. Front. Immunol. 2018, 9, 2816. [Google Scholar] [CrossRef]
- Visez, N.; Chassard, G.; Azarkan, N.; Naas, O.; Sénéchal, H.; Sutra, J.P.; Poncet, P.; Choël, M. Wind-induced mechanical rupture of birch pollen: Potential implications for allergen dispersal. J. Aerosol. Sci. 2015, 89, 77–84. [Google Scholar] [CrossRef]
- Silva, M.; Ribeiro, H.; Abreu, I.; Cruz, A.; da Silva, J.E. Effects of CO2 on Acer negundo pollen fertility, protein content, allergenic properties, and carbohydrates. Environ. Sci. Pollut. Res. 2015, 22, 6904–6911. [Google Scholar] [CrossRef]
- Ribeiro, H.; Duque, L.; Sousa, R.; Abreu, I. Ozone effects on soluble protein content of Acer negundo, Quercus robur and Platanus spp. pollen. Aerobiologia 2013, 29, 443–447. [Google Scholar] [CrossRef]
- Cuinica, L.G.; Abreu, I.; da Silva, J.C.E. In vitro exposure of Ostrya carpinifolia and Carpinus betulus pollen to atmospheric levels of CO, O3, and SO2. Environ. Sci. Pollut. Res. 2014, 21, 2256–2262. [Google Scholar] [CrossRef]
- Toro, R.; Córdova, A.; Canales, M.; Mardones, P. Trends and threshold exceedances analysis of airborne pollen concentrations in Metropolitan Santiago Chile. PLoS ONE 2015, 10, e0123077. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Diameter of Pollen Grain (μm) | PL/EL Ratio | Shape | |
|---|---|---|---|---|
| EL (μm) | PL (μm) | |||
| Q. acutissima | 38.12 ± 2.03 a | 29.63 ± 1.64 b | 0.78 c | Suboblate |
| Q. aliena | 34.37 ± 2.94 b | 28.21 ± 1.76 c | 0.83 b | Suboblate |
| Q. dentata | 34.33 ± 2.14 b | 29.01 ± 1.92 b | 0.85 a | Suboblate |
| Q. mongolica | 39.06 ± 2.35 a | 31.72 ± 1.77 a | 0.81 b | Suboblate |
| Q. serrata | 32.32 ± 1.61 c | 26.47 ± 1.11 d | 0.82 b | Suboblate |
| Q. variabilis | 33.40 ± 1.94 b | 25.42 ± 1.83 e | 0.76 c | Suboblate |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, I.; Kwak, M.J.; Lee, J.K.; Lim, Y.; Park, S.; Kim, H.; Lee, K.-A.; Woo, S.Y. Flowering Phenology and Characteristics of Pollen Aeroparticles of Quercus Species in Korea. Forests 2020, 11, 232. https://doi.org/10.3390/f11020232
Kim I, Kwak MJ, Lee JK, Lim Y, Park S, Kim H, Lee K-A, Woo SY. Flowering Phenology and Characteristics of Pollen Aeroparticles of Quercus Species in Korea. Forests. 2020; 11(2):232. https://doi.org/10.3390/f11020232
Chicago/Turabian StyleKim, Iereh, Myeong Ja Kwak, Jong Kyu Lee, Yeaji Lim, Sanghee Park, Handong Kim, Keum-Ah Lee, and Su Young Woo. 2020. "Flowering Phenology and Characteristics of Pollen Aeroparticles of Quercus Species in Korea" Forests 11, no. 2: 232. https://doi.org/10.3390/f11020232