Soil Available Phosphorus Loss Caused by Periodical Understory Management Reduce Understory Plant Diversity in a Northern Subtropical Pinus massoniana Plantation Chronosequence

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
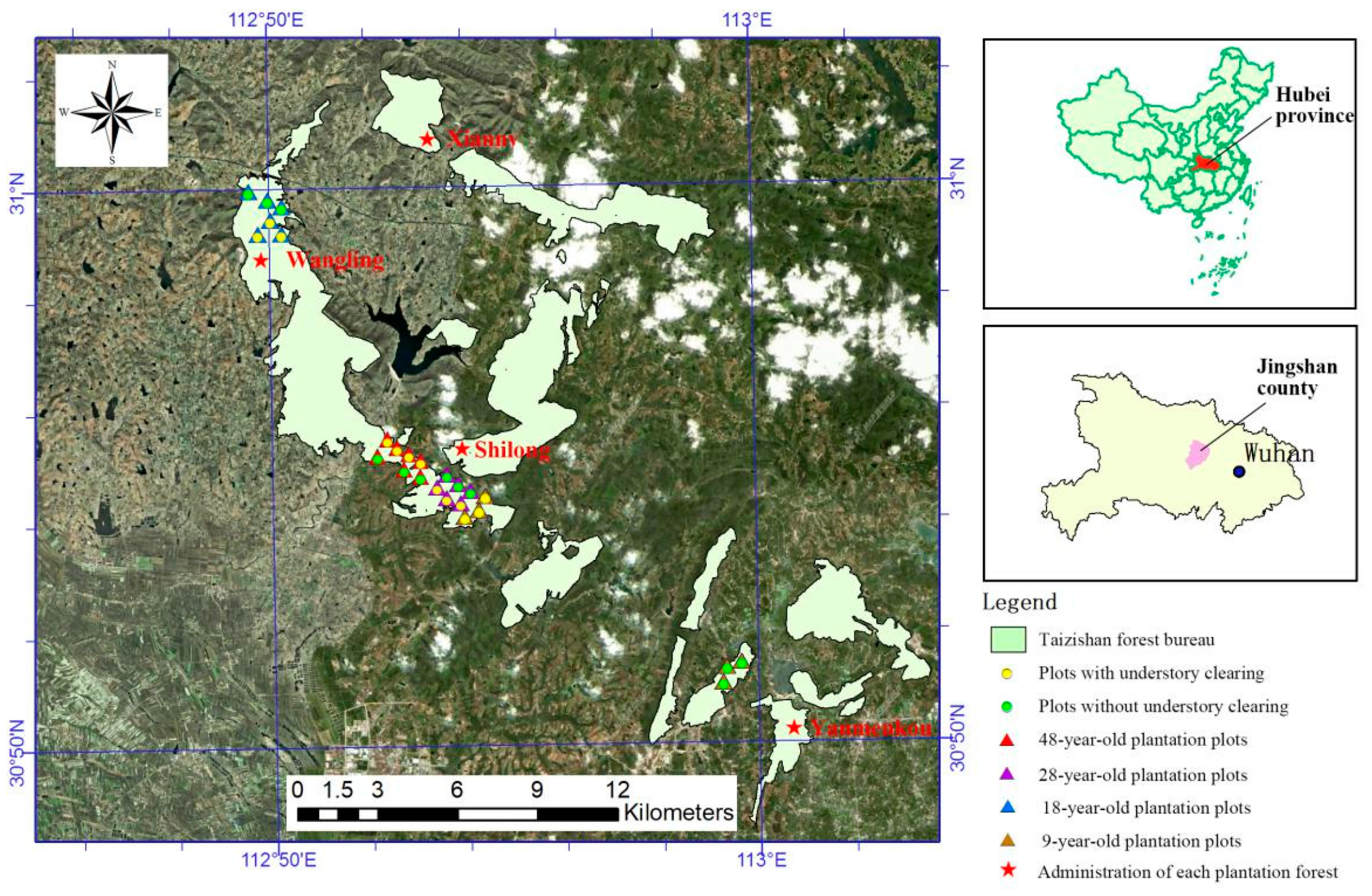
2.1. Study Site
2.2. Plot Settings and Investigation
2.3. Soil Collection and Measurement
2.4. Statistical Analyses
3. Results
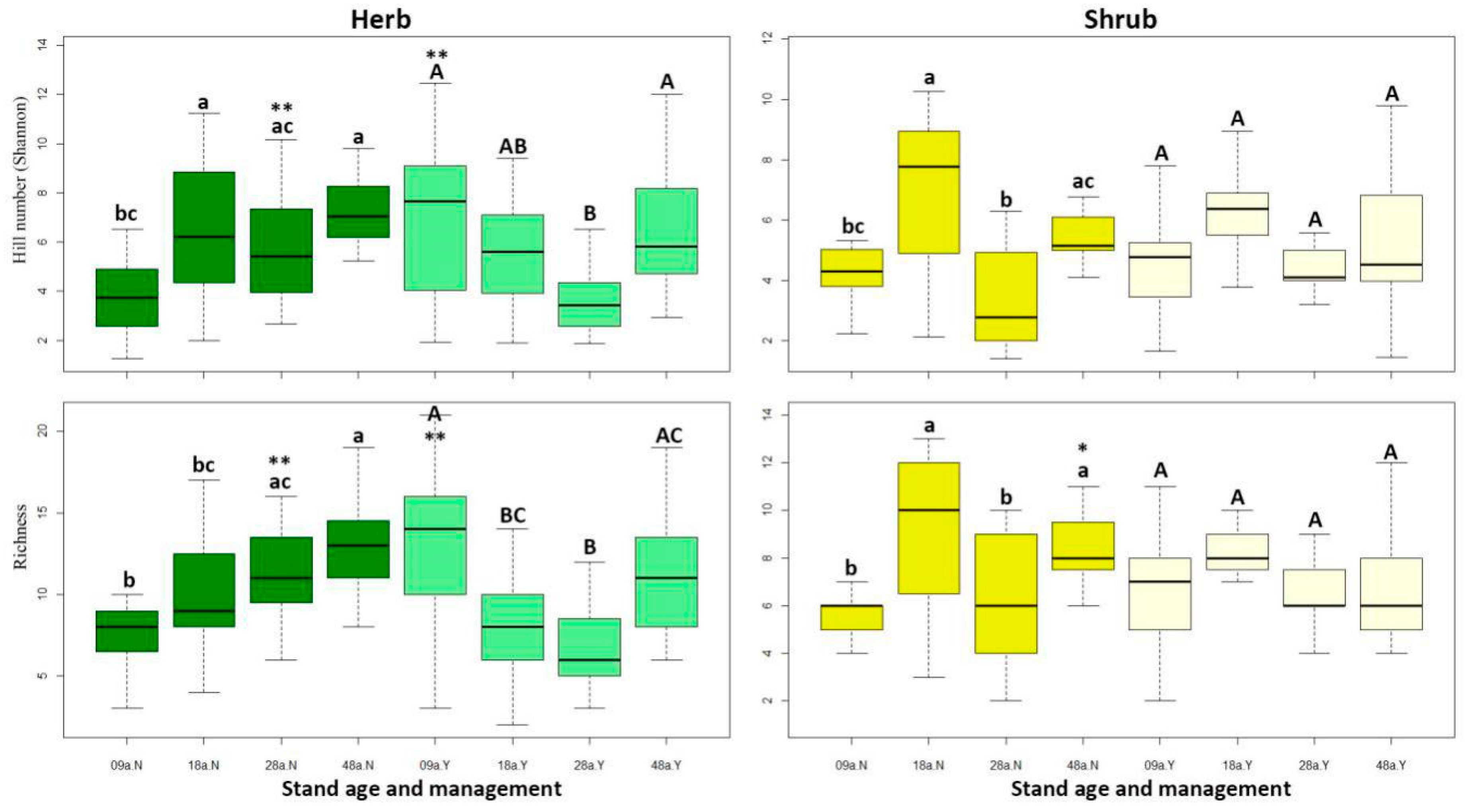
3.1. Influence of Stand Age and Understory Clearing on Understory Diversity
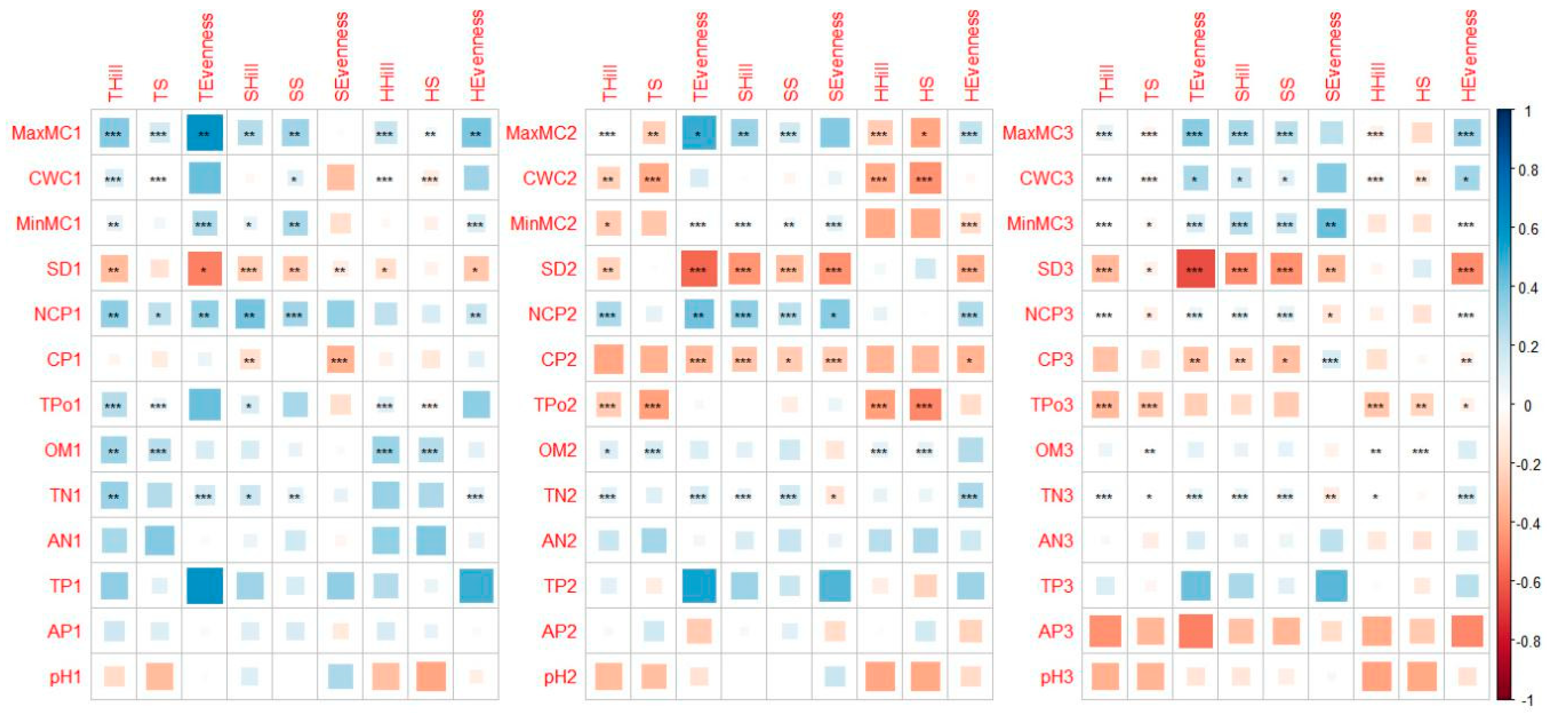
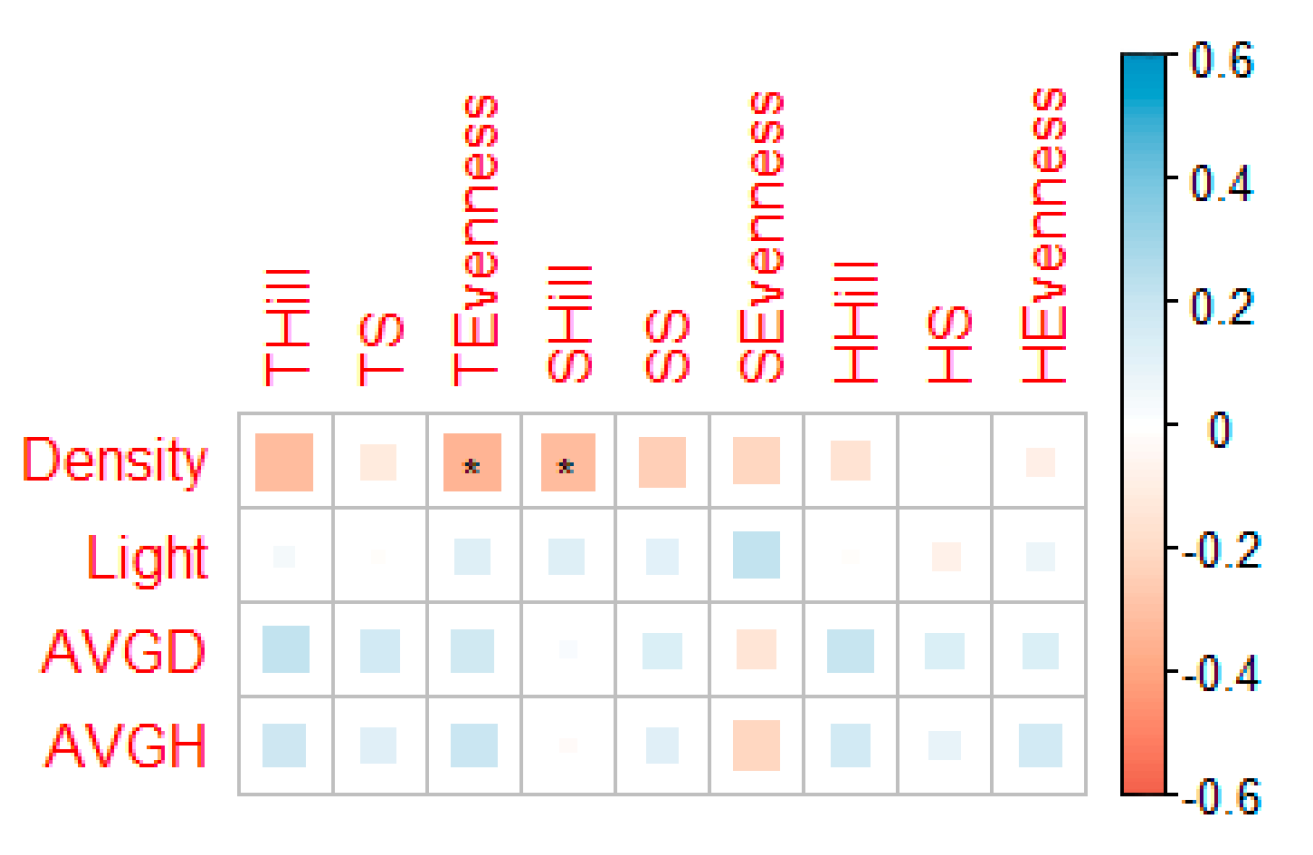
3.2. Relation Between Soil, Forest Factors, and Understory Vegetation Diversity
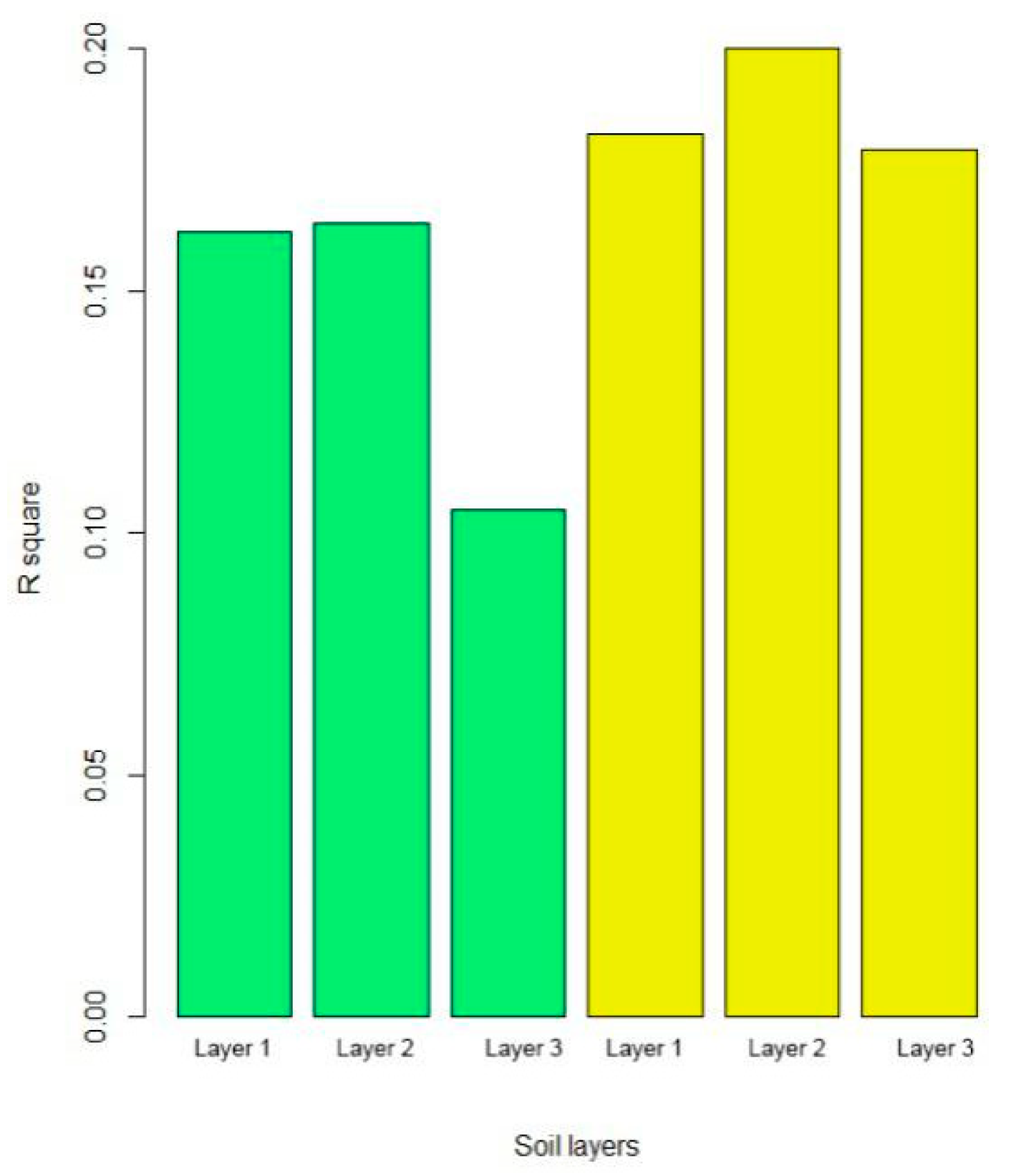
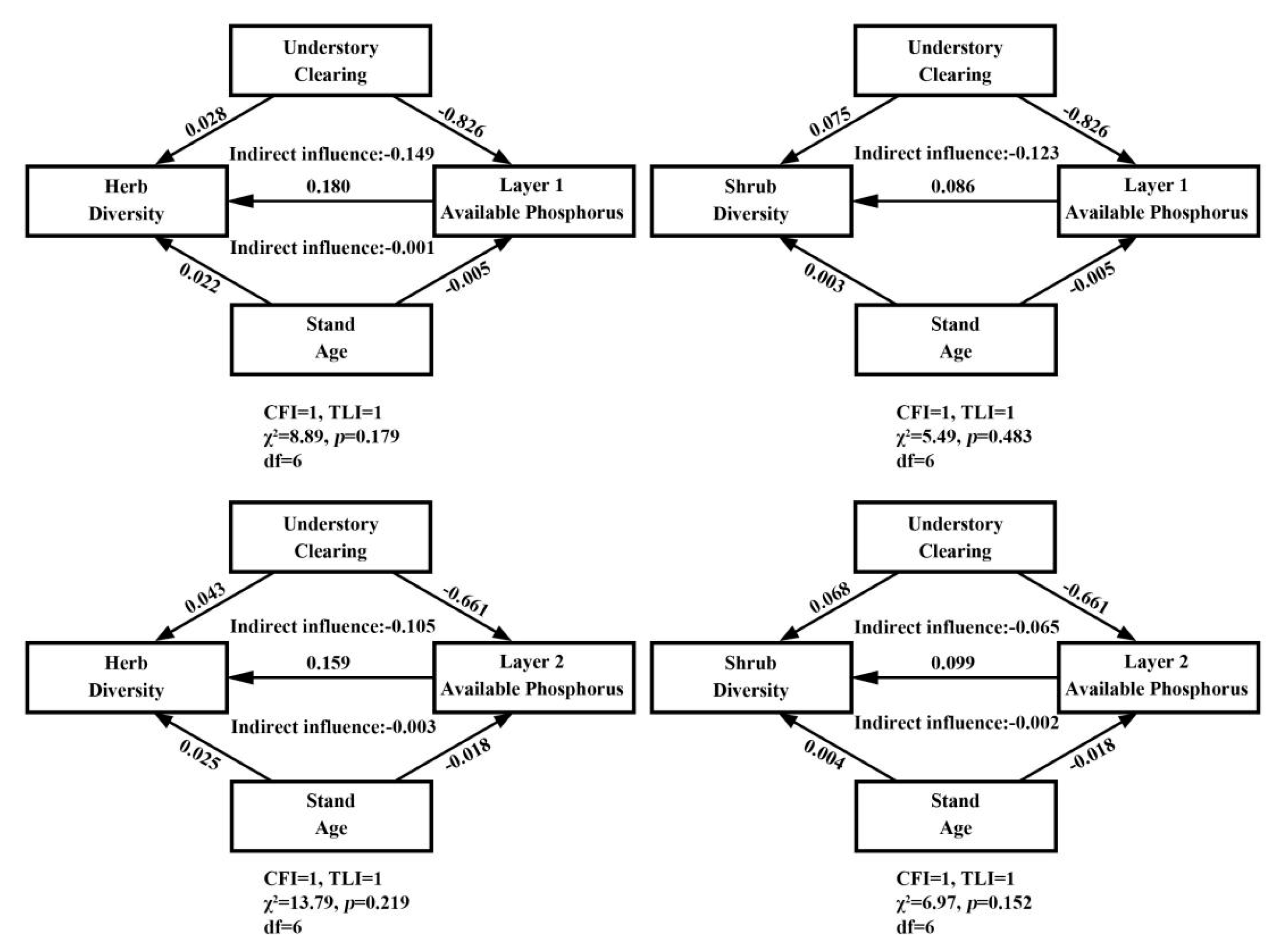
3.3. Structural Equation Modeling
4. Discussion
4.1. Influence of Forest Factors and Understory Management on Understory Diversity
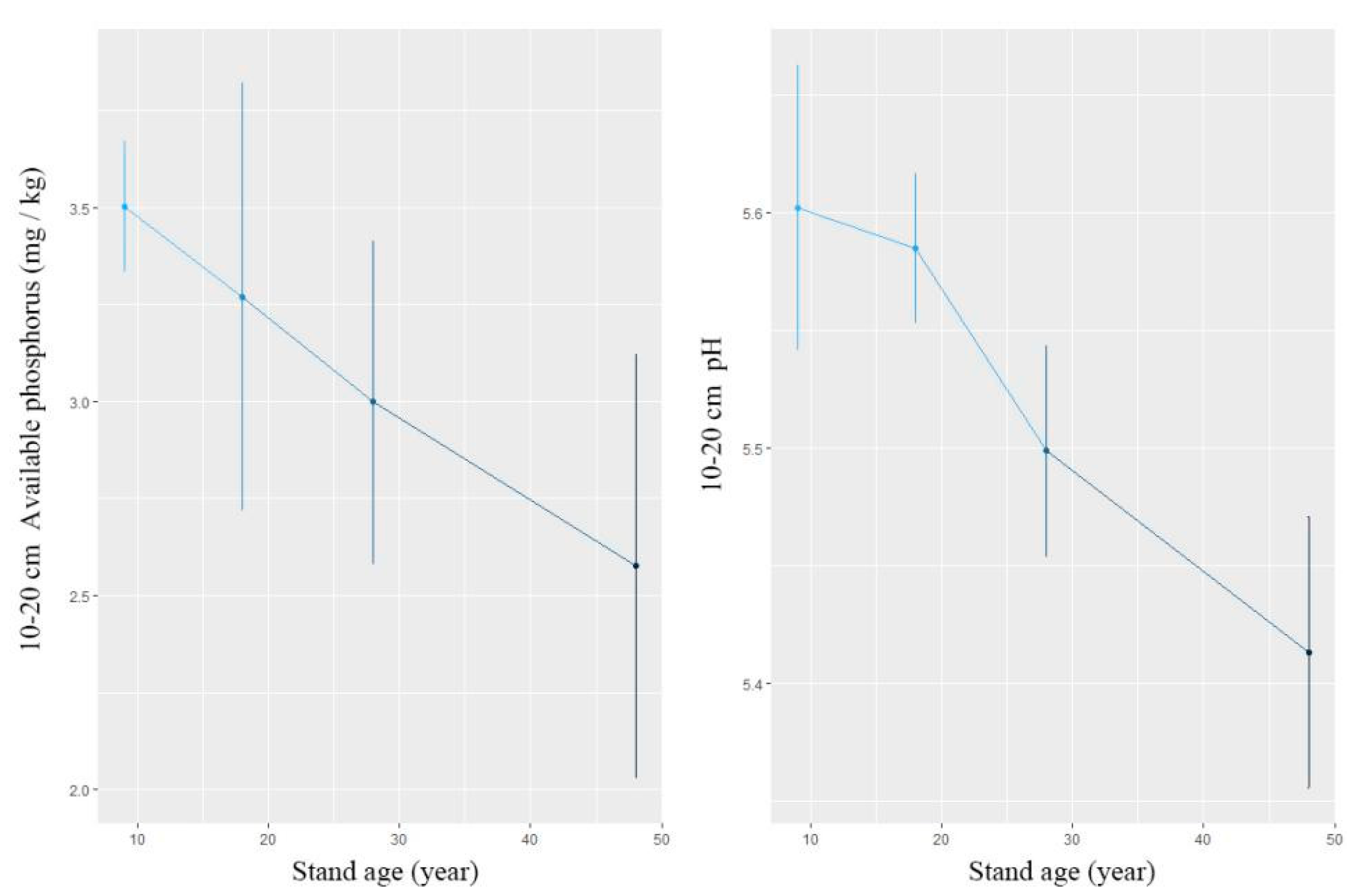
4.2. Relation Between Soil Factors and Understory Diversity
4.3. SEM Analysis on Available Phosphorus
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Ferraz, S.F.B. Role of eucalypt and other planted forests in biodiversity conservation and the provision of biodiversity-related ecosystem services. Forest Ecol. Manag. 2013, 301, 43–50. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Ferraz, S.F.B. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Ohsawa, M.; Shimokawa, T. Extending the rotation period in larch plantations increases canopy heterogeneity and promotes species richness and abundance of native beetles: Implications for the conservation of biodiversity. Biol. Conserv. 2011, 144, 3106–3116. [Google Scholar] [CrossRef]
- Tyler, N.; McFadden, R.D. Opening the silvicultural toolbox: A new framework for conserving biodiversity in Chilean timber plantations. Forest Ecol. Manag. 2018, 425, 75–84. [Google Scholar]
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. BioScience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Nearing, M.A.; Jetten, V.; Baffaut, C.; Cerdan, O.; Couturier, A.; Hernandez, M.; Le Bissonnais, Y.; Nichols, M.H.; Nunes, J.P.; Renschler, C.S.; et al. Modeling response of soil erosion and runoff to changes in precipitation and cover. Catena 2005, 61, 131–154. [Google Scholar] [CrossRef]
- Muller, R.N.; Bormann, F.H. Role of Erythronium americanum Ker. in Energy Flow and Nutrient Dynamics of a Northern Hardwood Forest Ecosystem. Science 1976, 193, 1126–1128. [Google Scholar] [CrossRef]
- Small, C.J.; McCarthy, B.C. Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. Forest Ecol. Manag. 2005, 217, 229–243. [Google Scholar] [CrossRef]
- Brevik, E.C.; Cerdà, A.; Mataix-Solera, J.; Pereg, L.; Quinton, J.N.; Six, J.; van Oost, K. The interdisciplinary nature of SOIL. Soil 2015, 1, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Rolo, V.; López-Díaz, M.L.; Moreno, G. Shrubs affect soil nutrients availability with contrasting consequences for pasture understory and tree overstory production and nutrient status in Mediterranean grazed open woodlands. Nutr. Cycl. Agroecosys. 2012, 93, 89–102. [Google Scholar] [CrossRef]
- Finzi, A.C.; Canham, C.D.; Breemen, N.V. Canopy Tree–Soil Interactions Within Temperate Forests: Species Effects on pH and Cations. Ecol. Appl. 1998, 8, 447–454. [Google Scholar]
- Jobbagy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Ray, G.; Ochoa, C.G.; Deboodt, T.; Mata-Gonzalez, R. Overstory–Understory Vegetation Cover and Soil Water Content Observations in Western Juniper Woodlands: A Paired Watershed Study in Central Oregon, USA. Forests 2019, 10, 151. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.J.; Liu, S.L.; Ma, K.M.; Zhu, Y.G. Relationships between soil characteristics, topography and plant diversity in a heterogeneous deciduous broad-leaved forest near Beijing, China. Plant. Soil 2004, 261, 47–54. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, W.; Ren, H.; Chen, H.; Wang, J. Impact of atmospheric nitrogen deposition on soil properties and herb-layer diversity in remnant forests along an urban–rural gradient in Guangzhou, southern China. Plant. Ecol. 2012, 213, 1187–1202. [Google Scholar] [CrossRef]
- Gao, Y.; Sun, S.; Xing, F.; Mu, X.; Bai, Y. Nitrogen addition interacted with salinity-alkalinity to modify plant diversity, microbial PLFAs and soil coupled elements: A 5-year experiment. Appl. Soil Ecol. 2019, 137, 78–86. [Google Scholar] [CrossRef]
- Laliberté, E.; Lambers, H.; Burgess, T.I.; Wright, S.J. Phosphorus limitation, soil-borne pathogens and the coexistence of plant species in hyperdiverse forests and shrublands. New Phytol. 2015, 206, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Zemunik, G.; Turner, B.L.; Lambers, H.; Laliberté, E. Diversity of plant nutrient- acquisition strategies increases during long-term ecosystem development. Nat. Plants 2015, 1, 15050. [Google Scholar] [CrossRef]
- Goebes, P.; Schmidt, K.; Seitz, S.; Both, S.; Bruelheide, H.; Erfmeier, A.; Scholten, T.; Kuhn, P. The strength of soil-plant interactions under forest is related to a Critical soil Depth. Sci. Rep. 2019, 9, 8635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, N.; Zhang, D.; Liu, Y.; Zhang, J.; Ou, J.; Zhang, J.; Deng, C.; Wei, J. Plant diversity and soil physicochemical properties under different aged Pinus Massoniana plantation. Shengtaixue Zazhi 2014, 10, 2610–2617. [Google Scholar]
- Chao, L.; Hong, T.; Li, J.; Chen, C.; Hong, W.; Wu, C. Analyses on diameter class structure and species diversity of understory of artificial forest of Cunninghamia lanceolata with different forest ages in mid-subtropical region. J. Plant. Resour. Environ. 2015, 24, 88–96. [Google Scholar]
- Chen, X.; Zhao, A.; Zhang, J.; Zeng, H. Understory Plant Diversity and Environmental Characteristics of Different Age Pinus massoniana Plantations. J. Sichuan Agric. Univ. 2017, 35, 186–192, 233. [Google Scholar]
- Rytter, L.; Stener, L.-G. Productivity and thinning effects in hybrid aspen (Populus tremula L. × P. tremuloides Michx.) stands in southern Sweden. Forestry 2005, 78, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Verschuyl, J.; Riffell, S.; Miller, D.; Wigley, T.B. Biodiversity response to intensive biomass production from forest thinning in North American forests—A meta-analysis. Forest Ecol. Manag. 2011, 261, 221–232. [Google Scholar] [CrossRef]
- Jiménez, M.N.; Spotswood, E.N.; Cañadas, E.M.; Navarro, F.B.; Verheyen, K. Stand management to reduce fire risk promotes understorey plant diversity and biomass in a semi-aridPinus halepensisplantation. Appl. Veg. Sci. 2015, 18, 467–480. [Google Scholar] [CrossRef]
- Ali, A.; Dai, D.; Akhtar, K.; Teng, M.; Yan, Z.; Urbina-Cardona, N.; Mullerova, J.; Zhou, Z. Response of understory vegetation, tree regeneration, and soil quality to manipulated stand density in a Pinus massoniana plantation. Glob. Ecol. Conserv. 2019, 20, e00775. [Google Scholar] [CrossRef]
- Misik, T.; Coll, E.K. Long-term response of understorey cover, basal area and diversity to stand density in a mixed oak forest on the Sikfokut plot in Hungary. J. For. Sci. 2013, 59, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Cao, Y. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol. Eng. 2014, 73, 238–245. [Google Scholar] [CrossRef]
- Agee, K.J.; Skinner, N.C. Basic principles of forest fuel reduction treatments. Forest Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Porto, M.; Correia, O.; Beja, P. Long-term consequences of mechanical fuel management for the conservation of Mediterranean forest herb communities. Biodivers. Conserv. 2011, 20, 2669–2691. [Google Scholar] [CrossRef]
- Decocq, G.; Valentin, B.; Toussaint, B.; Hendoux, F.; Saguez, R.; Bardat, J. Soil seed bank composition and diversity in a managed temperate deciduous forest. Biodivers. Conserv. 2004, 13, 2485–2509. [Google Scholar] [CrossRef]
- Santana, J.; Porto, M.; Reino, L.; Beja, P. Long-term understory recovery after mechanical fuel reduction in Mediterranean cork oak forests. Forest Ecol. Manag. 2011, 261, 447–459. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Jørgensen, H.; Larsen, J.R. Long-term influence of hay-cutting on plant species richness, biodiversity and soil fertility in a Danish fen. Ecol. Eng. 2019, 134, 93–100. [Google Scholar] [CrossRef]
- Thomas, S.C.; Halpern, C.B.; Falk, D.A.; Liguori, D.A.; Austin, K.A. Plant diversity in managed forests: Understory responses to thinning and fertilization. Ecol. Appl. 1999, 9, 864–879. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Ecroyd, C.E.; Leckie, A.C.; Kimberley, M.O. Diversity and succession of adventive and indigenous vascular understorey plants in Pinus radiata plantation forests in New Zealand. Forest Ecol. Manag. 2003, 185, 307–326. [Google Scholar] [CrossRef]
- Bao, F.; Jiang, Z. Wood Properties of Main Tree Species from Plantation in China, 2nd ed.; China Forestry Publishing House: Beijing, China, 1998; pp. 56–82. [Google Scholar]
- Granke, O. Assessment of Ground Vegetation. In ForestBIOTA Work Report; Federal Research Centre for Forestry and Forest Products: Hamburg, Germany, 2006; pp. 1–20. [Google Scholar]
- Rosseel, Y. Lavaan: An R package for structural equation modeling. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A language and environment for statstical computing, Version 3.4.0.; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Poudel, D.R.; Chen, H.Y.H.; Kc, M.; Ge, Z.; Bown, H.E.; Ruan, H. Understory Vegetation Dynamics across a Poplar Plantation Chronosequence in Reclaimed Coastal Saline Soil. Forests 2019, 10, 764. [Google Scholar] [CrossRef] [Green Version]
- Eldegard, K.; Scholten, J.; Stokland, J.N.; Granhus, A.; Lie, M. The influence of stand density on bilberry (Vaccinium myrtillus L.) cover depends on stand age, solar irradiation, and tree species composition. For. Ecol. Manag. 2019, 432, 582–590. [Google Scholar] [CrossRef]
- Griffis, K.L.; Crawford, J.A.; Wagner, M.R.; Moir, W.H. Understory response to management treatments in northern Arizona ponderosa pine forests. Forest Ecol. Manag. 2001, 146, 239–245. [Google Scholar] [CrossRef]
- Seer, F.K.; Schrautzer, J. Status, future prospects, and management recommendations for alkaline fens in an agricultural landscape: A comprehensive survey. J. Nat. Conserv. 2014, 22, 358–368. [Google Scholar] [CrossRef]
- Zhang, C.; Yan, X.; Yang, J. Estimation of Nitrogen Reserves in Forest Soils of China. J. Southwest Agric. Univ. 2004, 26, 572–579. [Google Scholar]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; Deyn, G.B.D.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Laliberté, E.; Grace, J.B.; Huston, M.A.; Lambers, H.; Teste, F.P.; Turner, B.L.; Wardle, D.A. How does pedogenesis drive plant diversity? Trends Ecol. Evol. 2013, 28, 331–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, E.A.; de Carvalho, C.J.R.; Vieira, I.C.G.; Figueiredo, R.D.; Moutinho, P.; Ishida, F.Y.; dos Santos, M.T.P.; Guerrero, J.B.; Kalif, K.; Saba, R.T. Nitrogen and phosphorus limitation of biomass growth in a tropical secondary forest. Ecol. Appl. 2004, 14, 150–163. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Modern Plant Physiology, 3rd ed.; Higher Education Press: Beijing, China, 2012; pp. 62–83. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Class/Age (Year) | Understory Treatment | Stand Density (Tree · ha−1) | Height (m) | DBH (cm) | Light Transmittance (%) |
|---|---|---|---|---|---|
| Young/9 | Cleared | 3651.3 ± 392.8 | 5.74 ± 0.3 | 9.1 ± 0.7 | 35.1 ± 2.8 |
| Intermediate/18 | Cleared | 1555.6 ± 239.1 | 11.4 ± 0.9 | 14.9 ± 0.2 | 33.6 ± 2.5 |
| Near mature/28 | Cleared | 1195.7 ± 51.3 | 15.3 ± 0.6 | 21.6 ± 0.4 | 49.8 ± 3.5 |
| Mature/48 | Cleared | 773.6 ± 91.8 | 16.8 ± 2.2 | 25.5 ± 2.9 | 48.2 ± 5.4 |
| Young/9 | No-cleared | 3466.2 ± 366.3 | 4.4 ± 0.3 | 8.9 ± 0.6 | 28.9 ± 7.6 |
| Intermediate/18 | No-cleared | 1455.3 ± 251.2 | 11.1 ± 0.5 | 15.4 ± 1.3 | 35.3 ± 4.9 |
| Near mature/28 | No-cleared | 1247.6 ± 282.5 | 14.2 ± 0.7 | 20.5 ± 1.6 | 30.4 ± 9.9 |
| Mature/48 | No-cleared | 1177.7 ± 143.2 | 15.8 ± 2.5 | 23.3 ± 2.3 | 34.2 ± 1.4 |
| Categories | Index | Stand Age | Understory Management | Interaction | |||
|---|---|---|---|---|---|---|---|
| F-Value | p | F-Value | p | F-Value | p | ||
| Herb | Shannon | 6.78 | <0.001 *** | 0.82 | 0.37 | 5.42 | <0.001 *** |
| Richness | 5.29 | <0.001 *** | 2.20 | 0.14 | 8.61 | <0.001 *** | |
| Shrub | Shannon | 6.97 | <0.001 *** | 0.02 | 0.89 | 2.46 | 0.06 |
| Richness | 4.75 | <0.001 *** | 0.59 | 0.44 | 2.35 | 0.07 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, D.; Ali, A.; Huang, X.; Teng, M.; Wu, C.; Zhou, Z.; Liu, Y. Soil Available Phosphorus Loss Caused by Periodical Understory Management Reduce Understory Plant Diversity in a Northern Subtropical Pinus massoniana Plantation Chronosequence. Forests 2020, 11, 231. https://doi.org/10.3390/f11020231
Dai D, Ali A, Huang X, Teng M, Wu C, Zhou Z, Liu Y. Soil Available Phosphorus Loss Caused by Periodical Understory Management Reduce Understory Plant Diversity in a Northern Subtropical Pinus massoniana Plantation Chronosequence. Forests. 2020; 11(2):231. https://doi.org/10.3390/f11020231
Chicago/Turabian StyleDai, Dong, Ashfaq Ali, Xin Huang, Mingjun Teng, Changguang Wu, Zhixiang Zhou, and Yu Liu. 2020. "Soil Available Phosphorus Loss Caused by Periodical Understory Management Reduce Understory Plant Diversity in a Northern Subtropical Pinus massoniana Plantation Chronosequence" Forests 11, no. 2: 231. https://doi.org/10.3390/f11020231