Integrating Neighborhood Effect and Supervised Machine Learning Techniques to Model and Simulate Forest Insect Outbreaks in British Columbia, Canada




Abstract
:1. Introduction
- Compare the performance of two methodologies, namely binomial regression and random forests, to model the MPB spread between 1999 and 2014.
- Evaluate the usefulness of a set of predictor variables, describing the influence of local topography and the state (i.e., infested/non-infested) of neighboring localities, to determine the extent and speed of the MPB infestation.
- Simulate possible land cover changes in 2020, due to MPB infestation.
2. Materials and Methods
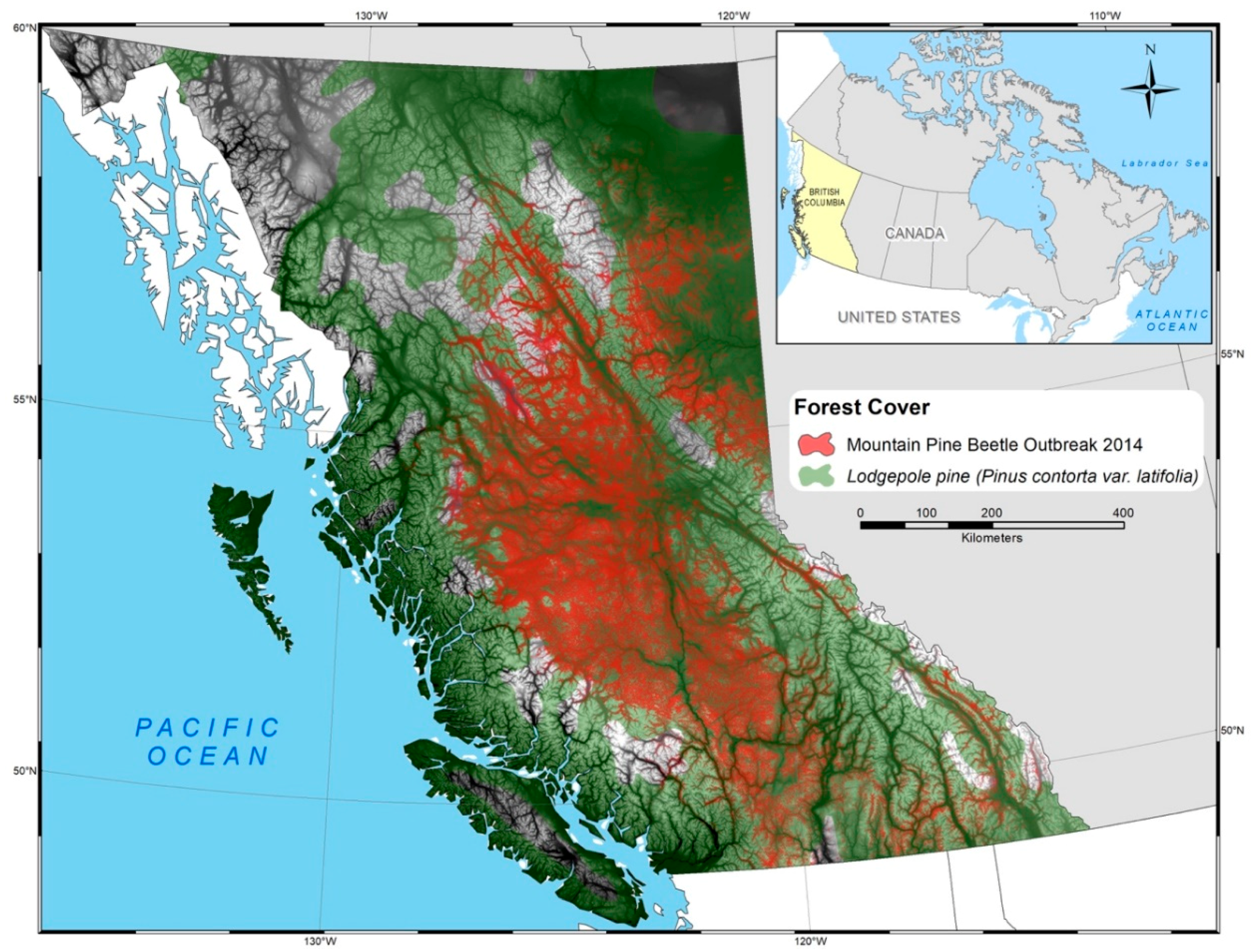
2.1. Study Area
2.2. Pine Mortality Dataset
2.3. Predictor Variables
- Elevation: MPB infestation has been observed to take place mostly at low or medium heights [47]. Elevation is defined as height above sea level per pixel. We used a Digital Elevation Map provided by GeoBC. The original pixel size of 500 was changed to 400 to match the resolution of the MPB infestation map.
- Aspect: The spread of the MPB infestation may benefit from milder temperatures [20,38,49] on south-oriented slopes. For that reason, aspect was calculated from the elevation map as the compass direction of the pixel slope face. We employed the “terrain” function of the “raster” R package. Next, it was sine-transformed to avoid the discontinuity at point 0–2π radians (0°–360°). Sine and cosine functions were used to avoid ambiguity at 0 radians.
- No-weighting ():
- Linear weighting (): weights decrease linearly until
- Inverse-distance weighting (): weights decrease as a function of the inverse of distance until
- Squared-inverse-distance weighting (): weights decrease as a function of the inverse of the squared distance until
2.4. Approaches to Model and Simulate Land Cover Changes
2.4.1. Generalized Linear Regression (GLM)
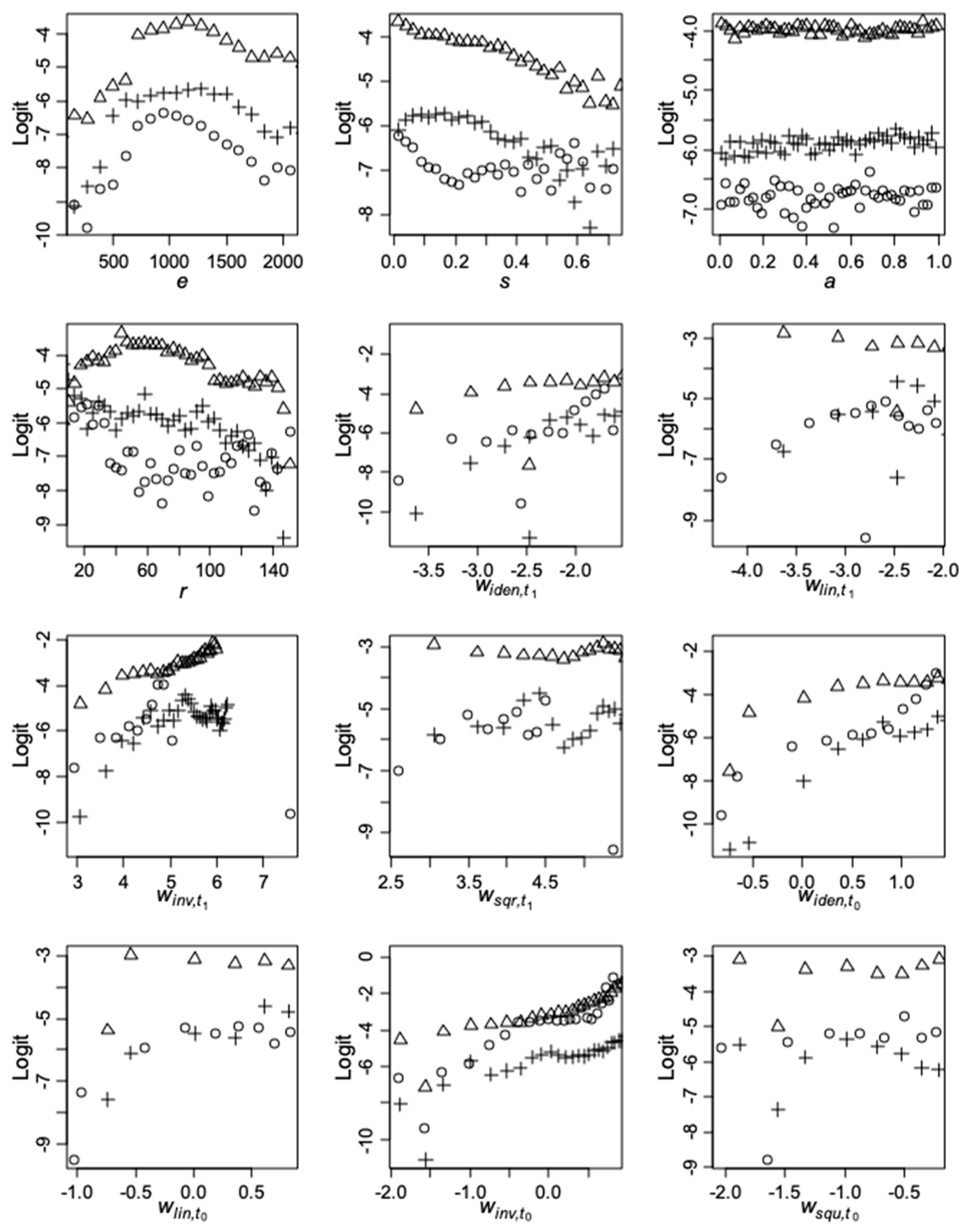
2.4.2. Random Forests (RF)
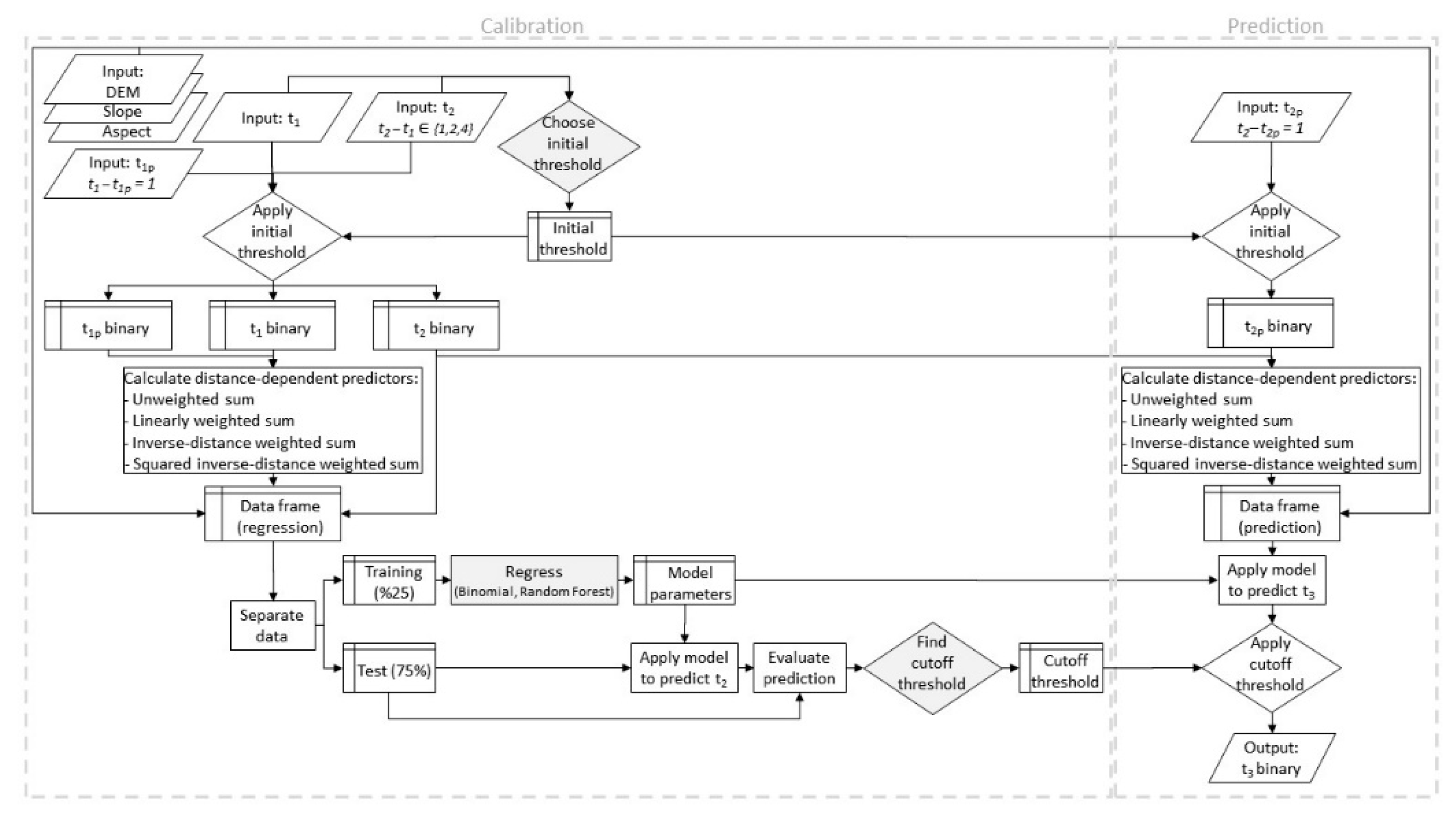
2.5. Model Calibration and Validation
2.6. Software
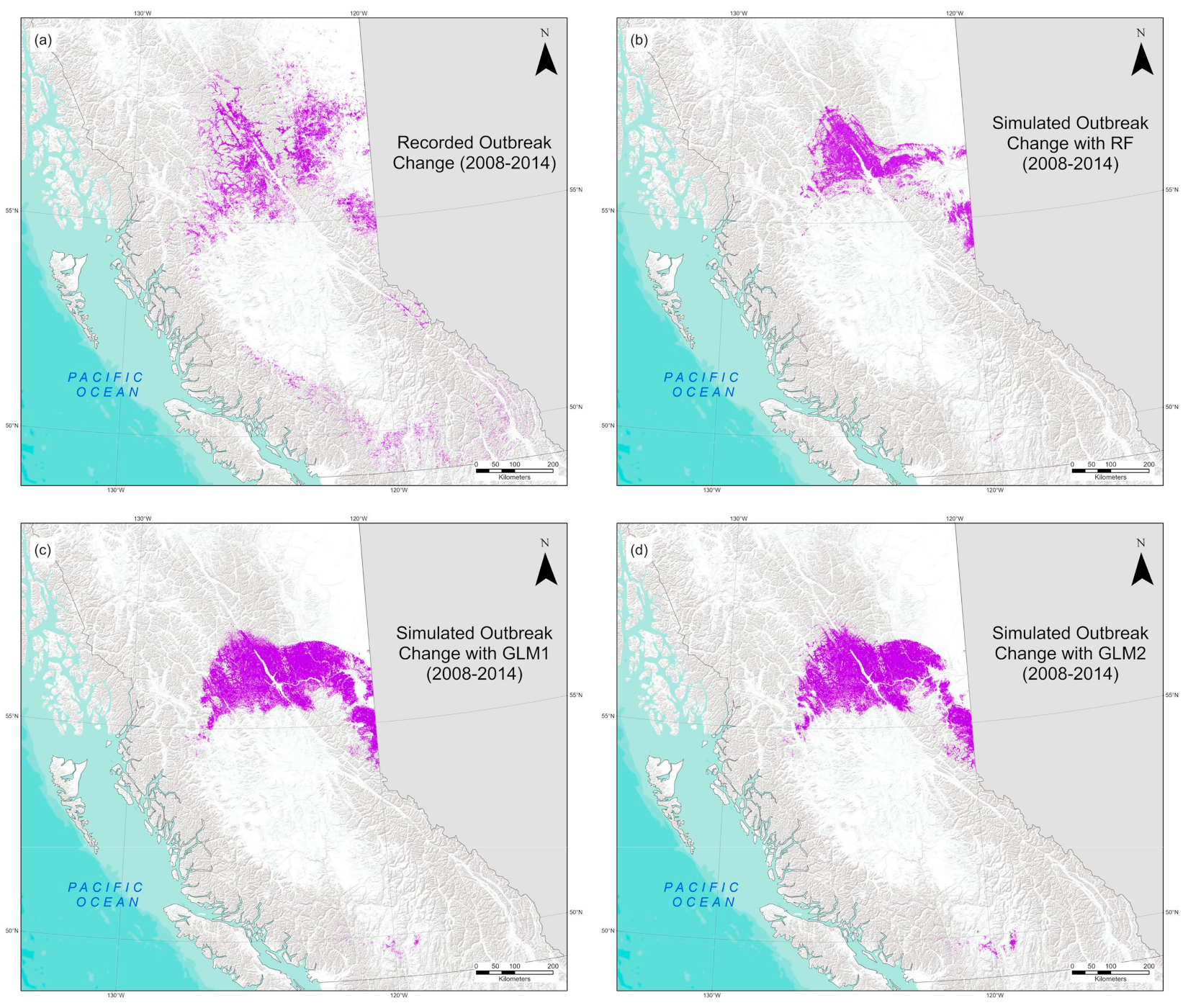
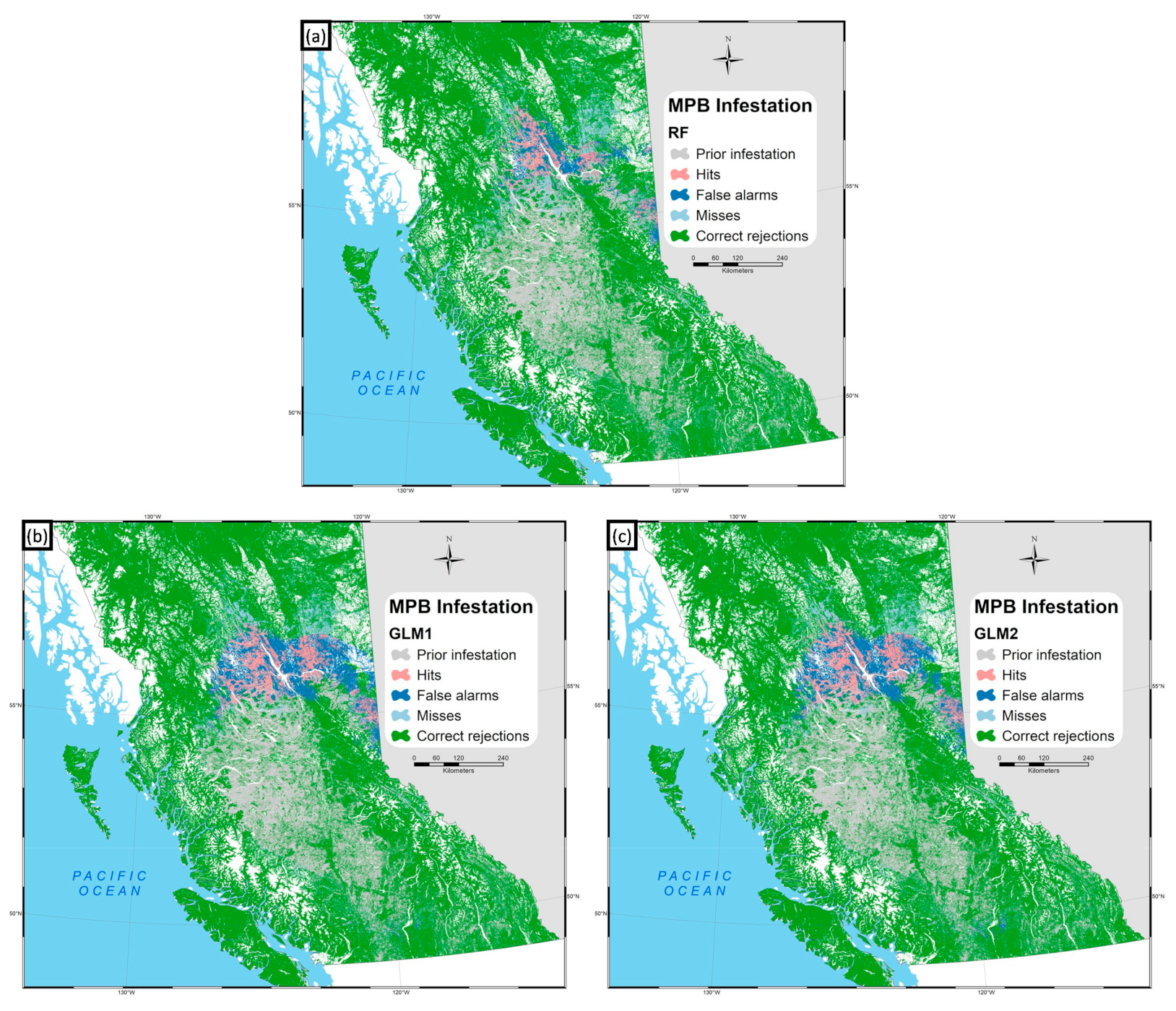
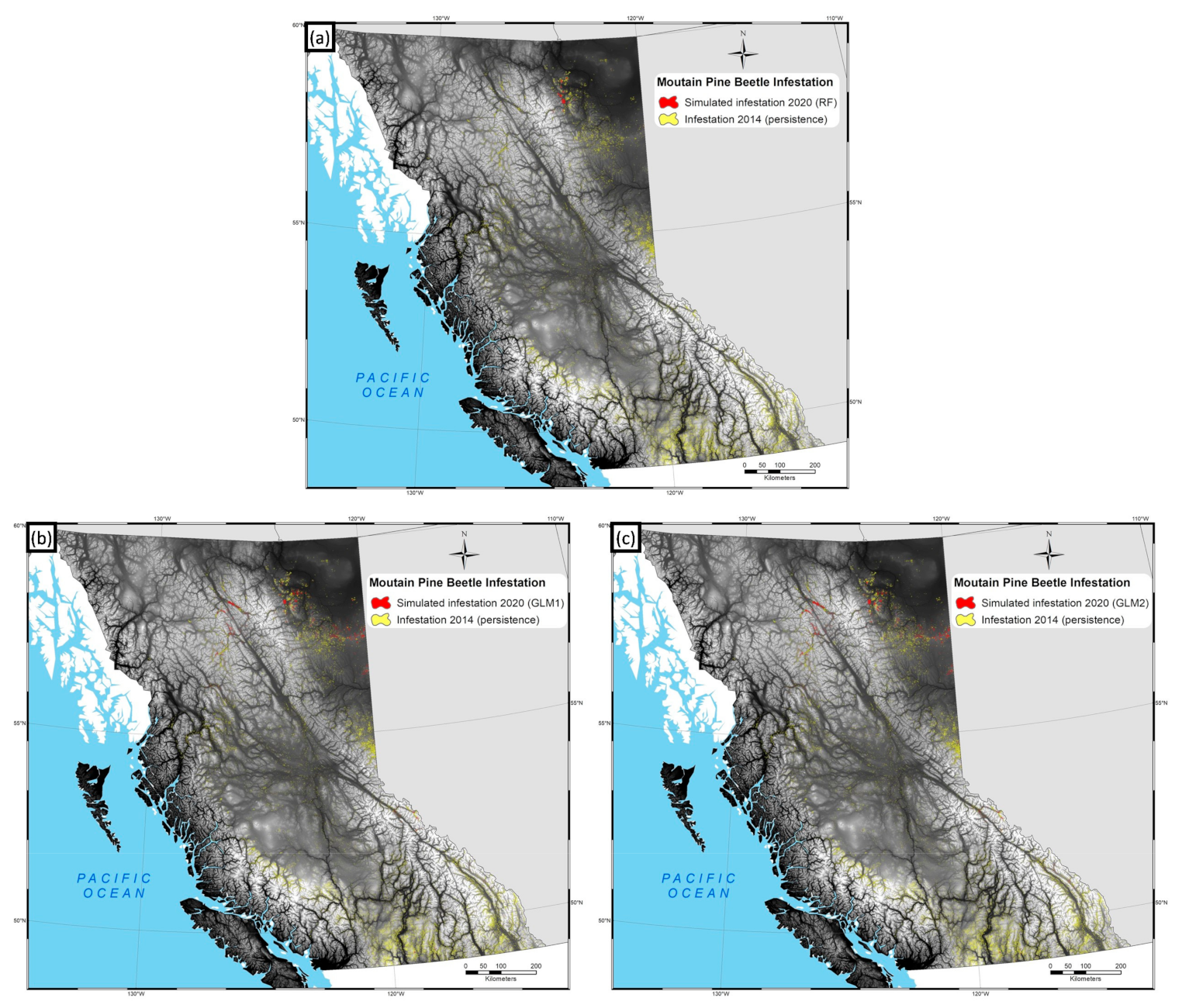
3. Results
4. Discussion
4.1. Model Validation
4.2. Model Predictions
4.3. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
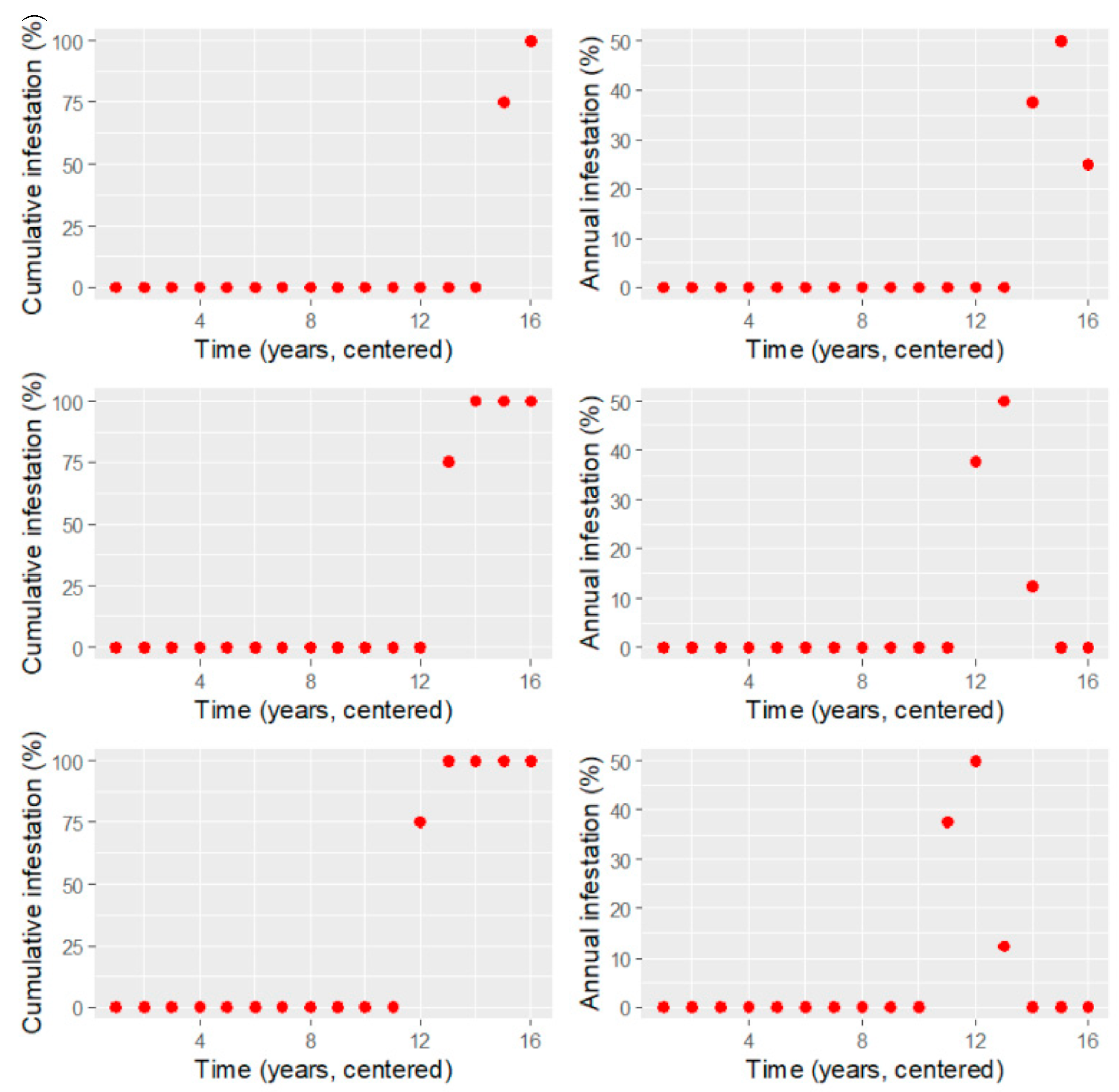
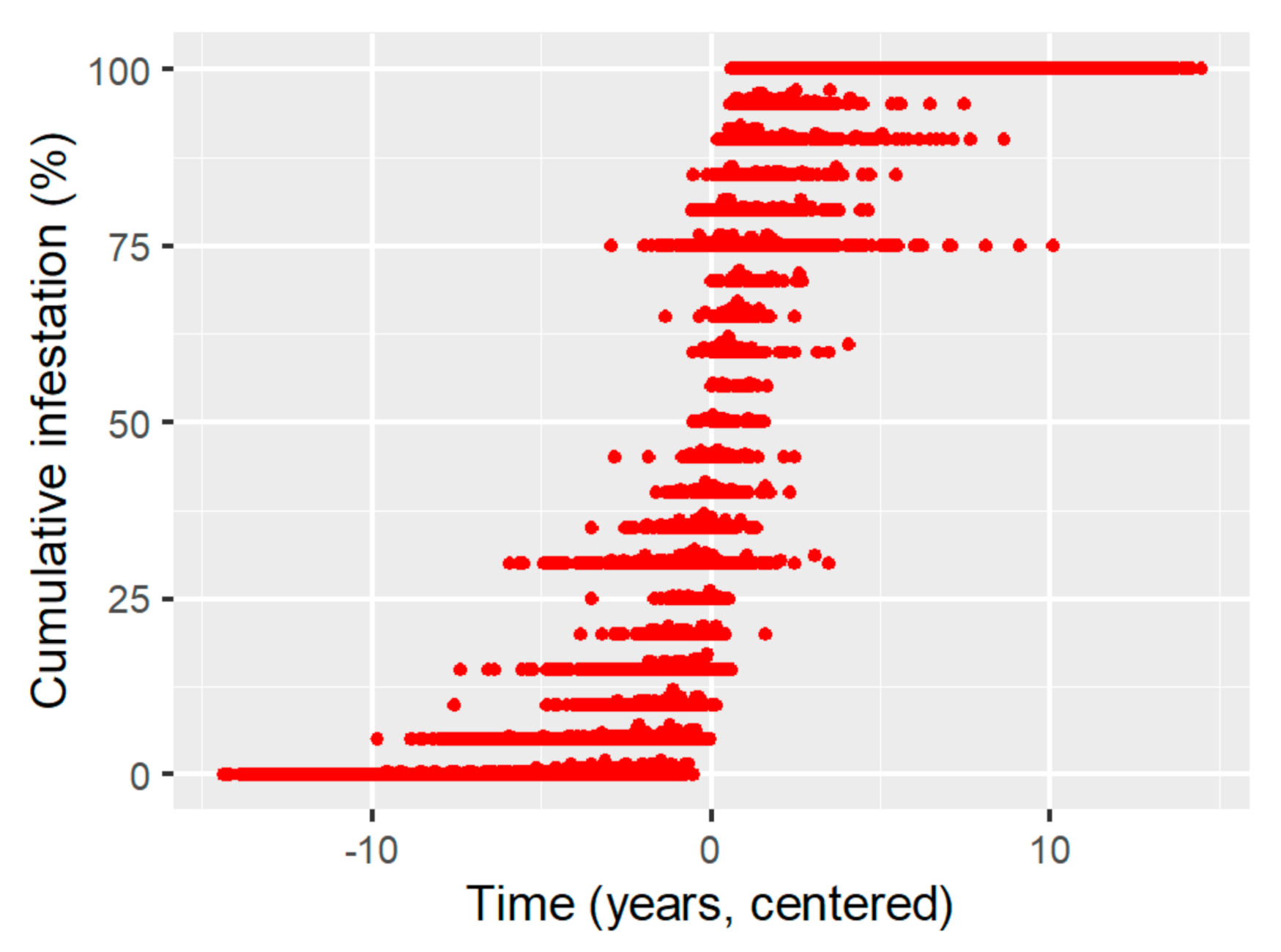
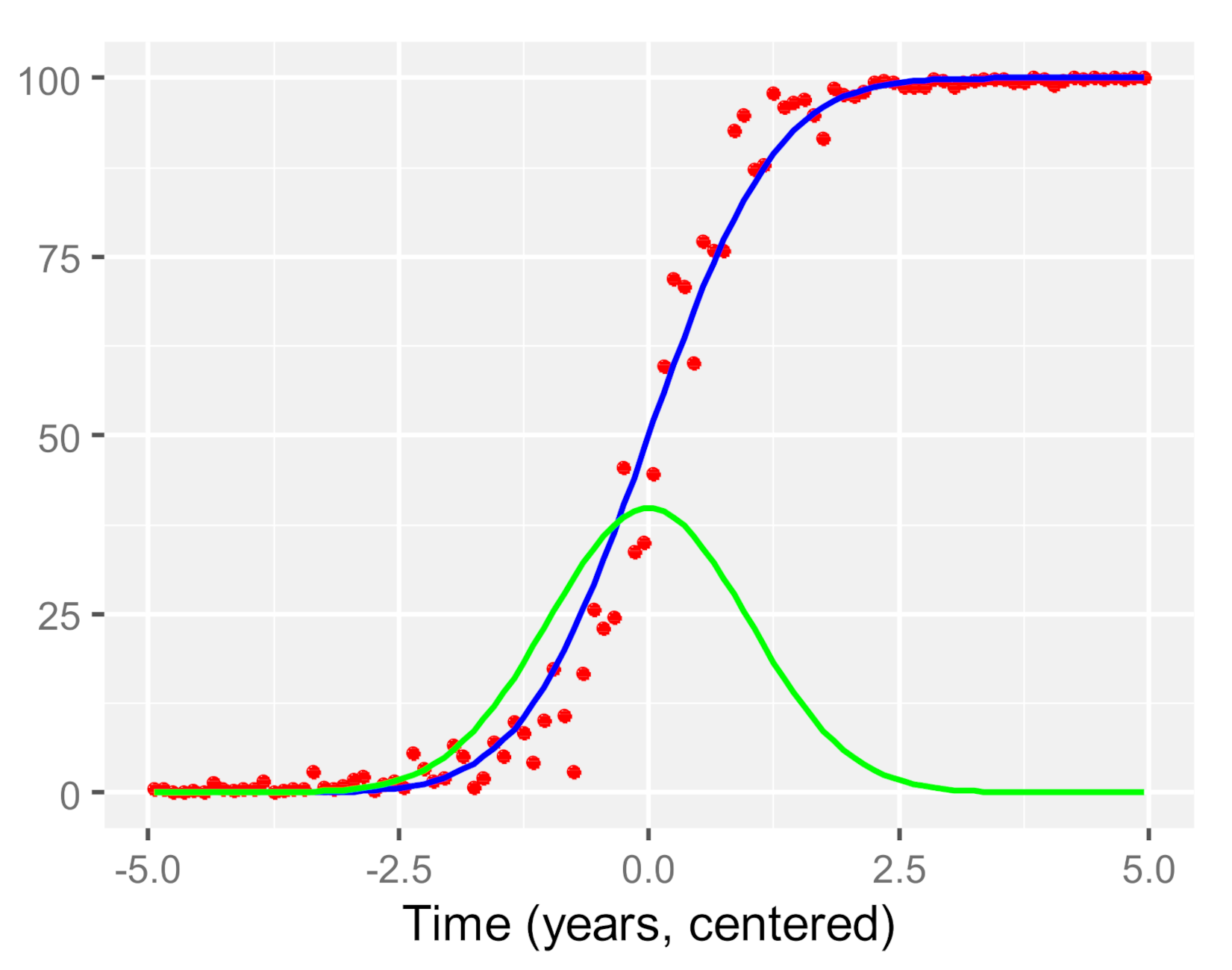
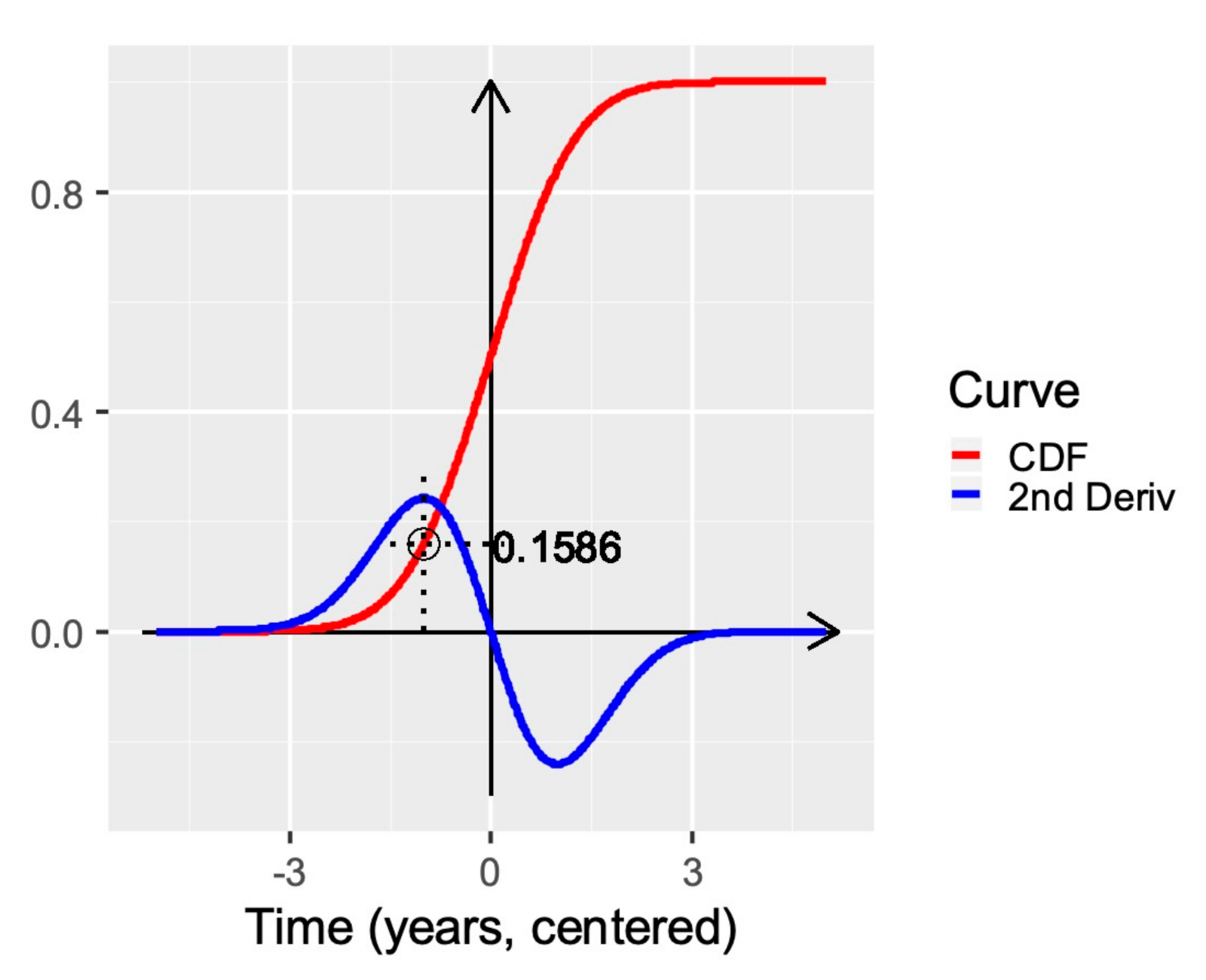
Appendix A. Calculation of Threshold Value on Cumulative Pinus contorta Mortality Data



- initial negligible or very low infestation that spreads slowly ( and are close to zero);
- a transitional phase in which the infestation starts picking up speed ( still low but increases);
- fast but steady infestation that increases constantly ( increases but reaches a maximum);
- transitional phase during which the infestation slows down ( increases further, decreases);
- saturation level ( highest, very close to zero).

Appendix B. Plots of Average Mortality vs. Predictors

Appendix C. Graphic Representation of the Four Different Neighborhood Types Implemented

Appendix D. Model Parameterization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Acronym | Estimate | Std. Error | z Value | Pr (>|z|) |
|---|---|---|---|---|---|
| (Intercept) | - | −2.518927 | 7.437363 × 10−3 | −338.685488 | 0.000000 |
| elevation | 2.415789 × 10−4 | 5.497418 × 10−6 | 43.944072 | 0.000000 | |
| ruggedness | 3.418483 × 10−4 | 3.661161 × 10−5 | 9.337157 | 9.895502 × 10−21 | |
| aspect.sin (sine) | −1.249106 × 10−2 | 2.968032 × 10−3 | −4.208534 | 2.570333 × 10−5 | |
| aspect.cos (cosine) | −8.139534 × 10−3 | 2.982952 × 10−3 | −2.728684 | 6.358761 × 10−3 | |
| slope | −2.682364 | 1.821996 × 10−2 | −147.221146 | 0.000000 | |
| identity.1 | −2.990441 × 10 | 3.971936 × 10−1 | −75.289257 | 0.000000 | |
| linear.1 | 6.979188 × 10−6 | 1.514023 × 10−7 | 46.096967 | 0.000000 | |
| inverse.1 | −3.289507 × 10−2 | 1.136399 × 10−3 | −28.946755 | 3.083004 × 10−184 | |
| squared.1 | −9.227856 × 10−2 | 4.722606 × 10−3 | −19.539752 | 5.042652 × 10−85 | |
| identity.2 | 3.895161 × 10 | 3.035022 × 10−1 | 128.340446 | 0.000000 | |
| linear.2 | −6.558742 × 10−6 | 1.114139 × 10−7 | −58.868233 | 0.000000 | |
| inverse.2 | 1.187859 × 10−2 | 8.019164 × 10−4 | 14.812753 | 1.211739 × 10−49 | |
| squared.2 | 1.341532 × 10−1 | 3.245616 × 10−3 | 41.333659 | 0.000000 |
| Variable | Acronym | Estimate | Std. Error | Z Value | Pr (>|z|) |
|---|---|---|---|---|---|
| (Intercept) | - | −5.792189 | 2.340479 × 10−2 | −247.478780 | 0.000000 |
| elevation | 6.035065 × 10−3 | 3.824616 × 10−5 | 157.795349 | 0.000000 | |
| ruggedness | 9.025530 × 10−4 | 3.765714 × 10−5 | 23.967649 | 6.049586 × 10−127 | |
| aspect.sin (sine) | −1.940276 × 10−2 | 2.988868 × 10−3 | −6.491677 | 8.488584 × 10−11 | |
| aspect.cos (cosine) | −8.848104 × 10−3 | 2.996480 × 10−3 | −2.952833 | 3.148722 × 10−3 | |
| slope | −2.494952 | 1.827487 × 10−2 | −136.523626 | 0.000000 | |
| identity.1 | −3.198807 × 10 | 3.989017 × 10−1 | −80.190364 | 0.000000 | |
| linear.1 | 6.664911 × 10−6 | 1.514503 × 10−7 | 44.007261 | 0.000000 | |
| inverse.1 | −2.847800 × 10−2 | 1.135279 × 10−3 | −25.084582 | 7.326996 × 10−139 | |
| squared.1 | −9.944317 × 10−2 | 4.717755 × 10−3 | −21.078495 | 1.253054 × 10−98 | |
| identity.2 | 3.926965 × 10 | 3.044064 × 10−1 | 129.004015 | 0.000000 | |
| linear.2 | −6.281823 × 10−6 | 1.115482 × 10−7 | −56.314888 | 0.000000 | |
| inverse.2 | 1.006898 × 10−2 | 8.026541 × 10−4 | 12.544607 | 4.255184 × 10−36 | |
| squared.2 | 1.190556 × 10−1 | 3.241775 × 10−3 | 36.725455 | 2.866455 × 10−295 | |
| I(elevation^2) | −2.332311 × 10−6 | 1.521917 × 10−8 | −153.248243 | 0.000000 |
| Variable | Acronym | Relative Importance Score |
|---|---|---|
| inverse.2 | 100.00 | |
| identity.2 | 99.96 | |
| linear.1 | 90.85 | |
| inverse.1 | 90.23 | |
| linear.2 | 80.11 | |
| identity.1 | 75.00 | |
| squared.1 | 74.04 | |
| squared.2 | 61.32 | |
| elevation | 22.43 | |
| slope | 11.16 | |
| aspect.cos (cosine) | 6.95 | |
| aspect.sin (sine) | 6.76 | |
| ruggedness | 3.83 |
Appendix E. Code and Data Availability
References
- McCullough, D.G.; Werner, R.A.; Neumann, D. Fire and Insects in Northern and Boreal Forest Ecosystems of North America. Annu. Rev. Entomol. 1998, 43, 107–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourbonnais, M.L.; Nelson, T.A.; Wulder, M.A. Geographic analysis of the impacts of mountain pine beetle infestation on forest fire ignition. Can. Geogr. Géographe Can. 2014, 58, 188–202. [Google Scholar] [CrossRef]
- Robbins, J. Bark Beetles Kill Millions of Acres of Trees in West-NYTimes.com. New York Times, 17 November 2008. [Google Scholar]
- Axelson, J.N.; Alfaro, R.I.; Hawkes, B.C. Changes in stand structure in uneven-aged lodgepole pine stands impacted by mountain pine beetle epidemics and fires in central British Columbia. For. Chron. 2010, 86, 87–99. [Google Scholar] [CrossRef]
- MacLean, D.A. Impacts of insect outbreaks on tree mortality, productivity, and stand development. Can. Entomol. 2016, 148, S138–S159. [Google Scholar] [CrossRef] [Green Version]
- Pelz, K.A.; Smith, F.W. Thirty year change in lodgepole and lodgepole/mixed conifer forest structure following 1980s mountain pine beetle outbreak in western Colorado, USA. For. Ecol. Manag. 2012, 280, 93–102. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Sturtevant, B.R.; Achtemeier, G.L.; Charney, J.J.; Anderson, D.P.; Cooke, B.J.; Townsend, P.A. Long-distance dispersal of spruce budworm (Choristoneura fumiferana Clemens) in Minnesota (USA) and Ontario (Canada) via the atmospheric pathway. Agric. For. Meteorol. 2013, 168, 186–200. [Google Scholar] [CrossRef]
- Patriquin, M.N.; Wellstead, A.M.; White, W.A. Beetles, trees, and people: Regional economic impact sensitivity and policy considerations related to the mountain pine beetle infestation in British Columbia, Canada. For. Policy Econ. 2007, 9, 938–946. [Google Scholar] [CrossRef]
- Chang, W.-Y.; Lantz, V.A.; Hennigar, C.R.; MacLean, D.A. Economic impacts of forest pests: A case study of spruce budworm outbreaks and control in New Brunswick, Canada. Can. J. For. Res. 2012, 42, 490–505. [Google Scholar] [CrossRef]
- Corbett, L.J.; Withey, P.; Lantz, V.A.; Ochuodho, T.O. The economic impact of the mountain pine beetle infestation in British Columbia: Provincial estimates from a CGE analysis. Forestry 2016, 89, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Petersen, B.; Stuart, D. Explanations of a changing landscape: A critical examination of the British Columbia bark beetle epidemic. Environ. Plan. A 2014, 46, 598–613. [Google Scholar] [CrossRef]
- Flint, C.G.; McFarlane, B.; Müller, M. Human dimensions of forest disturbance by insects: An international synthesis. Environ. Manag. 2009, 43, 1174–1186. [Google Scholar] [CrossRef]
- Strohm, S.; Reid, M.L.; Tyson, R.C. Impacts of management on Mountain Pine Beetle spread and damage: A process-rich model. Ecol. Model. 2016, 337, 241–252. [Google Scholar] [CrossRef]
- James, P.M.A.; Huber, D.P.W. TRIA-Net: 10 years of collaborative research on turning risk into action for the mountain pine beetle epidemic. Can. J. For. Res. 2019, 49, iii–v. [Google Scholar] [CrossRef]
- Bone, C.; Nelson, M.F. Improving Mountain Pine Beetle Survival Predictions Using Multi-Year Temperatures Across the Western USA. Forests 2019, 10, 866. [Google Scholar] [CrossRef] [Green Version]
- Safranyik, L.; Carroll, A. The biology and epidemiology of the mountain pine beetle in lodgepole pine forests. In The Mountain Pine Beetle: A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Safranyik, L., Wilson, W.R., Eds.; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2007; pp. 3–66. [Google Scholar]
- Bentz, B.J.; Boone, C.; Raffa, K.F. Tree response and mountain pine beetle attack preference, reproduction and emergence timing in mixed whitebark and lodgepole pine stands. Agric. For. Entomol. 2015, 17, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Logan, J.A.; Powell, J.A. Ghost forests, global warming, and the mountain pine beetle (Coleoptera: Scolytidae). Am. Entomol. 2001, 47, 160–173. [Google Scholar] [CrossRef]
- Safranyik, L.; Carroll, A.L.; Régnière, J.; Langor, D.W.; Riel, W.G.; Shore, T.L.; Peter, B.; Cooke, B.J.; Nealis, V.G.; Taylor, S.W. Potential for range expansion of mountain pine beetle into the boreal forest of North America. Can. Entomol. 2010, 142, 415–442. [Google Scholar] [CrossRef]
- Hodge, J.; Cooke, B.; McIntosh, R. A Strategic Approach to Slow the Spread of Mountain Pine Beetle across Canada; Natural Resources Canada, Canadian Forest Service, Canadian Council of Forest Ministers: Ottawa, ON, Canada, 2017.
- Hall, R.J.; Castilla, G.; White, J.C.; Cooke, B.J.; Skakun, R.S. Remote sensing of forest pest damage: A review and lessons learned from a Canadian perspective. Can. Entomol. 2016, 148, S296–S356. [Google Scholar] [CrossRef]
- Ferretti, M. Forest health assessment and monitoring–Issues for consideration. Environ. Monit. Assess. 1997, 48, 45–72. [Google Scholar] [CrossRef]
- Bentz, B.J.; Jönsson, A.M. Modeling Bark Beetle Responses to Climate Change. Bark Beetles 2015, 533–553. [Google Scholar] [CrossRef]
- Cooke, B.J.; Carroll, A.L. Predicting the risk of mountain pine beetle spread to eastern pine forests: Considering uncertainty in uncertain times. For. Ecol. Manag. 2017, 396, 11–25. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.A.; Waring, R.H. Modeling lodgepole and jack pine vulnerability to mountain pine beetle expansion into the western Canadian boreal forest. For. Ecol. Manag. 2012, 274, 161–171. [Google Scholar] [CrossRef]
- Wulder, M.A.; Ortlepp, S.M.; White, J.C.; Nelson, T.; Coops, N.C. A provincial and regional assessment of the mountain pine beetle epidemic in British Columbia: 1999–2008. J. Environ. Inform. 2010, 15, 1–13. [Google Scholar] [CrossRef]
- Liang, L.; Hawbaker, T.J.; Chen, Y.; Zhu, Z.; Gong, P. Characterizing recent and projecting future potential patterns of mountain pine beetle outbreaks in the Southern Rocky Mountains. Appl. Geogr. 2014, 55, 165–175. [Google Scholar] [CrossRef]
- Powell, J.A.; Logan, J.A.; Bentz, B.J. Local projections for a global model of mountain pine beetle attacks. J. Theor. Biol. 1996, 179, 243–260. [Google Scholar] [CrossRef] [Green Version]
- Safranyik, L.; Barclay, H.; Thomson, A.; Riel, W.G. A Population Dynamics Model for the Mountain Pine Beetle, Dendroctonus Ponderosae Hopk. (Coleoptera: Scolytidae); Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 1999.
- Lewis, M.A.; Nelson, W.; Xu, C. A Structured Threshold Model for Mountain Pine Beetle Outbreak. Bull. Math. Biol. 2010, 72, 565–589. [Google Scholar] [CrossRef] [Green Version]
- Bone, C.; Wulder, M.A.; White, J.C.; Robertson, C.; Nelson, T.A. A GIS-based risk rating of forest insect outbreaks using aerial overview surveys and the local Moran’s I statistic. Appl. Geogr. 2013, 40, 161–170. [Google Scholar] [CrossRef]
- Macias Fauria, M.; Johnson, E.A. Large-scale climatic patterns and area affected by mountain pine beetle in British Columbia, Canada. J. Geophys. Res. 2009, 114, G01012. [Google Scholar] [CrossRef] [Green Version]
- Bone, C.; Dragicevic, S.; Roberts, A. Integrating high resolution remote sensing, GIS and fuzzy set theory for identifying susceptibility areas of forest insect infestations. Int. J. Remote Sens. 2005, 26, 4809–4828. [Google Scholar] [CrossRef]
- Liang, L.; Li, X.; Huang, Y.; Qin, Y.; Huang, H. Integrating remote sensing, GIS and dynamic models for landscape-level simulation of forest insect disturbance. Ecol. Model. 2017, 354, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bone, C.; Altaweel, M. Modeling micro-scale ecological processes and emergent patterns of mountain pine beetle epidemics. Ecol. Model. 2014, 289, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Pérez, L.; Dragićević, S. ForestSimMPB: A swarming intelligence and agent-based modeling approach for mountain pine beetle outbreaks. Ecol. Inform. 2011, 6, 62–72. [Google Scholar] [CrossRef]
- Perez, L.; Dragicevic, S. Landscape-level simulation of forest insect disturbance: Coupling swarm intelligent agents with GIS-based cellular automata model. Ecol. Model. 2012, 231, 53–64. [Google Scholar] [CrossRef]
- Haughian, S.R.; Burton, P.J.; Taylor, S.W.; Curry, C. Expected effects of climate change on forest disturbance regimes in British Columbia. BC J. Ecosyst. Manag. 2012, 13. [Google Scholar]
- Axelson, J.N.; Alfaro, R.I.; Hawkes, B.C. Influence of fire and mountain pine beetle on the dynamics of lodgepole pine stands in British Columbia, Canada. For. Ecol. Manag. 2009, 257, 1874–1882. [Google Scholar] [CrossRef]
- Klenner, W.; Walton, R.; Arsenault, A.; Kremsater, L. Dry forests in the Southern Interior of British Columbia: Historic disturbances and implications for restoration and management. For. Ecol. Manag. 2008, 256, 1711–1722. [Google Scholar] [CrossRef]
- Lemmen, D.S.; Warren, F.J.; Lacroix, J.; Bush, E. From Impacts to Adaptation: Canada in a Changing Climate 2007; Government of Canada: Ottawa, QC, Canada, 2008; ISBN 987-0662-05176-3.
- Natural Resources Canada Mountain Pine Beetle. Available online: http://www.nrcan.gc.ca/forests/fire-insects-disturbances/top-insects/13381 (accessed on 5 November 2020).
- BC Ministry of Forests. Forest Health Aerial Overview Survey Standards for British Columbia: The B.C. Ministry of Forests Adaptation of the Canadian Forest Service’s FHN Report 97-1 “Overview Aerial Survey Standards for British Columbia and the Yukon”; BC Ministry of Forests: Victoria, BC, Canada, 2000.
- Shore, T.; Safranyik, L. Susceptibility and Risk Rating Systems for the Mountain Pine Beetle in Lodgepole Pine Stands; Pacific Forestry Centre: Victoria, BC, Canada, 1992.
- Carroll, A.L.; Aukema, B.H.; Raffa, K.F.; Linton, D.A.; Smith, G.D.; Lindgren, B.S. Mountain Pine Beetle Outbreak Development: The Endemic–Incipient Epidemic Transition; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2006.
- Wulder, M.A.; Dymond, C.C.; White, J.C.; Erickson, B.; Safranyik, L.; Wilson, B. Detection, mapping, and monitoring of the mountain pine beetle. In The Mountain Pine Beetle: A Synthesis of Biology, Management, and Impacts on Lodgepole Pine; Safranyik, L., Wilson, B., Eds.; Natural Resources Canada, Canadian Forest Service, Pacific Forestry Centre: Victoria, BC, Canada, 2006; pp. 123–154. ISBN 0-662-42623-1. [Google Scholar]
- McIntire, E.J.B. Understanding natural disturbance boundary formation using spatial data and path analysis. Ecology 2004, 85, 1933–1943. [Google Scholar] [CrossRef]
- Milne, M.; Lewis, D. Considerations for rehabilitating naturally disturbed stands: Part 1—Watershed hydrology. BC J. Ecosyst. Manag. 2011, 11, 55–65. [Google Scholar]
- Reid, R.W. Biology of the Mountain Pine Beetle, Dendroctonus monticolae Hopkins, in the East Kootenay Region of British Columbia II. Behaviour in the Host, Fecundity, and Internal Changes in the Female. Can. Entomol. 1962, 94, 605–613. [Google Scholar] [CrossRef]
- Riley, S.J.; DeGloria, S.D.; Elliot, S.D. A Terrain Ruggedness Index that Quantifies Topographic Heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman and Hall: London, UK, 1989; ISBN 0412317605. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, version 3.5.0; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Pontius, R.G.; Huffaker, D.; Denman, K. Useful techniques of validation for spatially explicit land-change models. Ecol. Model. 2004, 179, 445–461. [Google Scholar] [CrossRef]
- Rutherford, G.N.; Guisan, A.; Zimmermann, N.E. Evaluating sampling strategies and logistic regression methods for modelling complex land cover changes. J. Appl. Ecol. 2007, 44, 414–424. [Google Scholar] [CrossRef]
- White, R.; Engelen, G. Cellular automata and fractal urban form: A cellular modelling approach to the evolution of urban land-use patterns. Environ. Plan. A 1993, 25, 1175–1199. [Google Scholar] [CrossRef] [Green Version]
- ESRI. ArcGIS Pro; version 2.4; Environmental Systems Research Institute: Redlands, CA, USA, 2019.
- BC Ministry of Forests. [Dataset] BC MPB Observed Cumilative Kil; BC Ministry of Forests: Victoria, BC, Canada, 2015; Volume 12.





| Predictor Description | Acronym | Units | Time |
|---|---|---|---|
| Elevation | m | - | |
| Aspect | Arbitrary | - | |
| Slope | Radians | - | |
| Ruggedness | Arbitrary | - | |
| Identity | Arbitrary | ||
| Linear weight | Arbitrary | ||
| Inverse-distance weight | Arbitrary | ||
| Square-inverse-distance weight | Arbitrary | ||
| No-weight | Arbitrary | ||
| Linear weight | Arbitrary | ||
| Inverse-distance weight | Arbitrary | ||
| Square-inverse-distance weight | Arbitrary |
| Algorithm | Cutoff | Hits (Pixels) | Misses (Pixels) | False Alarms (Pixels) | Figure of Merit (%) | Overall Accuracy (%) |
|---|---|---|---|---|---|---|
| Binomial (GLM1) | Kappa | 115,511 | 149,877 | 227,472 | 23.4 | 91.0 |
| Binomial (GLM1) | Youden’s J | 171,359 | 94,029 | 433,560 | 24.5 | 87.4 |
| Binomial-parabolic elevation (GLM2) | Kappa | 120,653 | 144,735 | 214,695 | 25.1 | 91.5 |
| Binomial-parabolic elevation (GLM2) | Youden’s J | 182,475 | 82,913 | 488,038 | 24.2 | 86.4 |
| Random forest (RF) | Kappa | 74,883 | 190,505 | 108,450 | 20 | 92.9 |
| Random forest (RF) | Youden’s J | 90,631 | 174,757 | 144,737 | 22.1 | 92.4 |
| Algorithm | Cutoff | Cumulative Volume of Pine Killed (%) |
|---|---|---|
| Binomial (GLM1) | Kappa | 57.5 |
| Binomial (GLM1) | Youden’s J | 62.7 |
| Binomial-parabolic elevation (GLM2) | Kappa | 57.8 |
| Binomial-parabolic elevation (GLM2) | Youden’s J | 63.3 |
| Random forest (RF) | Kappa | 54 |
| Random forest (RF) | Youden’s J | 55 |
| Algorithm | Cutoff | Cumulative Volume of Pine Killed (%) |
|---|---|---|
| Binomial (GLM1) | Kappa | 64.1 |
| Binomial (GLM1) | Youden’s J | 70.5 |
| Binomial-parabolic elevation (GLM2) | Kappa | 64.2 |
| Binomial-parabolic elevation (GLM2) | Youden’s J | 69.9 |
| Random forest (RF) | Kappa | 64 |
| Random forest (RF) | Youden’s J | 64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harati, S.; Perez, L.; Molowny-Horas, R. Integrating Neighborhood Effect and Supervised Machine Learning Techniques to Model and Simulate Forest Insect Outbreaks in British Columbia, Canada. Forests 2020, 11, 1215. https://doi.org/10.3390/f11111215
Harati S, Perez L, Molowny-Horas R. Integrating Neighborhood Effect and Supervised Machine Learning Techniques to Model and Simulate Forest Insect Outbreaks in British Columbia, Canada. Forests. 2020; 11(11):1215. https://doi.org/10.3390/f11111215
Chicago/Turabian StyleHarati, Saeed, Liliana Perez, and Roberto Molowny-Horas. 2020. "Integrating Neighborhood Effect and Supervised Machine Learning Techniques to Model and Simulate Forest Insect Outbreaks in British Columbia, Canada" Forests 11, no. 11: 1215. https://doi.org/10.3390/f11111215