The Short-Term Impact of Different Silvicultural Thinnings on Soil Nematode and Microarthropod Biodiversity in Artificial Black Pine Stands

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Treatments Description
- Thinning from below (TB)—only dominated, small, or standing dead trees were harvested (negative selection) in the forest.
- Selective thinning (ST)—(i) selection of 100 trees per hectare based on the quality of phenotype, i.e., its mechanical stability and vigor (candidate trees), (ii) making candidate trees free from their direct competitors, i.e., all the surrounding trees limiting the full crown growth of the candidate tree. The average number of candidate trees was around 100 trees per hectare. The density of 100 trees per hectare (10 m, the average distance between trees) came from the analysis of crown development patterns of black pine in the absence of lateral competition and by experimental data measured for this purpose. The aim of selective thinning was to leave free space available to the crowns of candidate trees. Selective thinning, changing both the horizontal and vertical layering, originates the stand structure variation [18].
2.2. Sampling Design
2.3. Soil Parameters
2.4. Meteorological Parameters
2.5. Forest Parameters
2.6. Soil Nematode Community Analysis
2.7. Soil Microarthropod Analysis
2.8. Statistical Analysis
3. Results
3.1. Meteorological and Hydrological Parameters
3.2. Effects of Silvicultural Treatments on Forest Structure and Dendrometrical Parameters
3.3. Similarity Analysis on Soil Nematode and Microarthropod Communities
3.4. Soil Nematode and Microarthropod Indicators
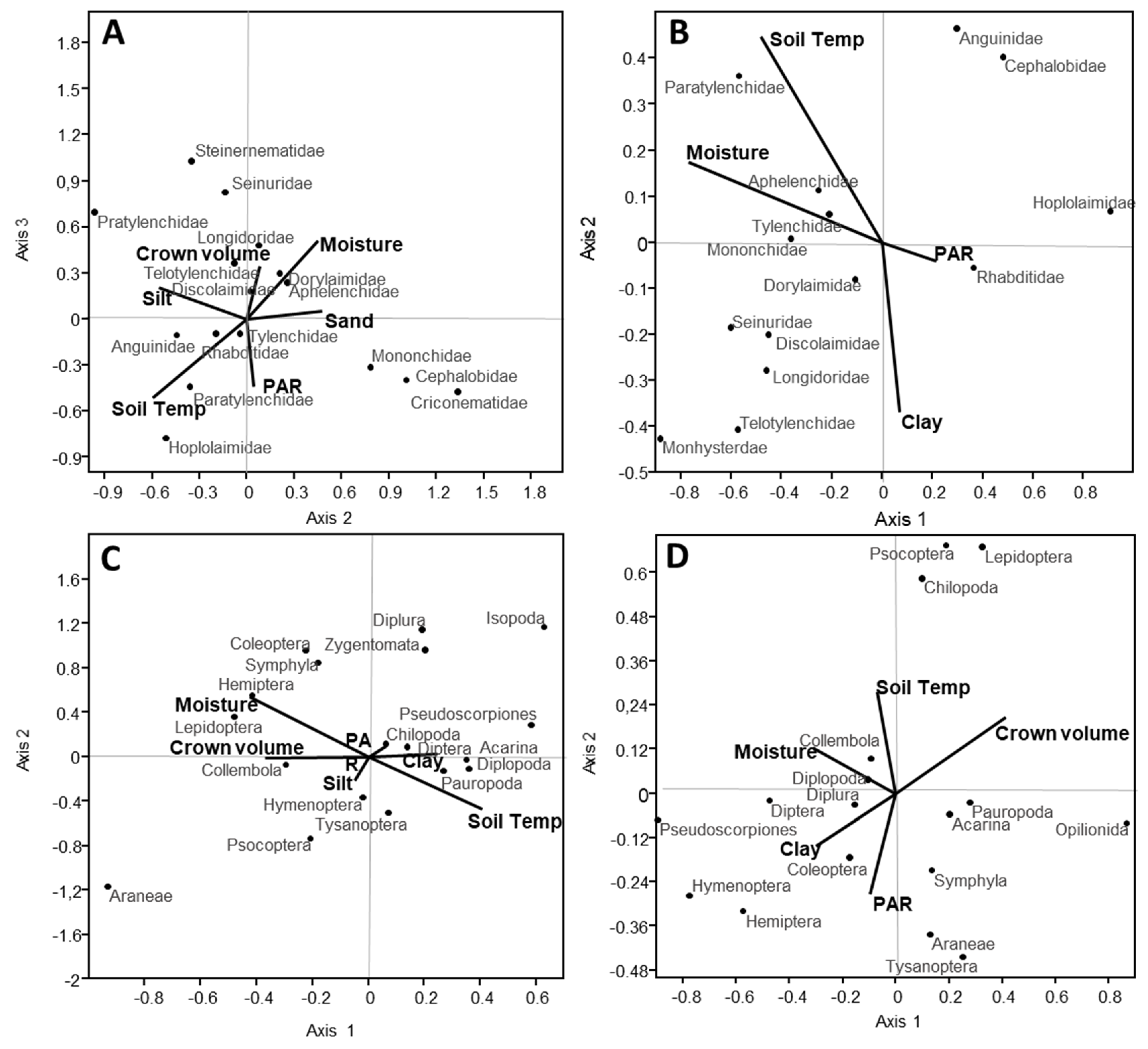
3.5. Relationship among Soil and Forest Variables on Nematode and Microarthropod Community Structure
4. Discussion
4.1. Effect of Thinnings on Soil Nematode and Microarthropod Community Structures
4.2. Environmental Parameters Influencing Soil Nematode and Microarthropod Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Schaefer, M.; Schauermann, J. The soil fauna of beech forests: Comparison between a mull and a moder soil. Pedobiologia 1990, 34, 299–314. [Google Scholar]
- Giller, K.E.; Beare, M.H.; Lavelle, P.; Izac, A.-M.; Swift, M.J. Agricultural intensification, soil biodiversity and agroecosystem function. Appl. Soil Ecol. 1997, 6, 3–16. [Google Scholar] [CrossRef]
- Irshad, U.; Villenave, C.; Brauman, A.; Plassard, C. Grazing by nematodes on rhizosphere bacteria enhances nitrate and phosphorus availability to Pinus pinaster seedlings. Soil Biol. Biochem. 2011, 43, 2121–2126. [Google Scholar] [CrossRef]
- Salome, C.; Coll, P.; Lardo, E.; Villenave, C.; Blanchart, E.; Hinsinger, P.; Marsden, C.; Le Cadre, E. Relevance of use-invariant soil properties to assess soil quality of vulnerable ecosystems: The case of Mediterranean vineyards. Ecol. Indic. 2014, 43, 83–93. [Google Scholar] [CrossRef]
- Bird, S.B.; Coulson, R.N.; Fisher, R.F. Changes in soil and litter arthropod abundance following tree harvesting and site preparation in a loblolly pine (Pinus taeda L.) plantation. For. Ecol. Manag. 2004, 202, 195–208. [Google Scholar] [CrossRef]
- Roversi, P.F.; Nannelli, R. Arthropods and Nematodes: Functional Biodiversity in Forest Ecosystems. In Forest Ecosystems–More than Just Trees; Blanco, J.A., Lo, Y.-H., Eds.; INTECH: London, UK, 2012; pp. 29–52. [Google Scholar]
- Marchi, M.; Chiavetta, U.; Cantiani, P. Assessing the mechanical stability of trees in artificial plantations of Pinus nigra JF Arnold using the LWN tool under different site indexes. Ann. Silvic. Res. 2017, 41, 48–53. [Google Scholar]
- Torras, O.; Saura, S. Effects of silvicultural treatments on forest biodiversity indicators in the Mediterranean. For. Ecol. Manag. 2008, 255, 3322–3330. [Google Scholar] [CrossRef]
- Pretzsch, H. Forest dynamics, growth, and yield. In Forest Dynamics, Growth and Yield; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–39. [Google Scholar]
- Kuehne, C.; Weiskittel, A.R.; Fraver, S.; Puettmann, K.J. Effects of thinning-induced changes in structural heterogeneity on growth, ingrowth, and mortality in secondary coastal Douglas-fir forests. Can. J. For. Res. 2015, 45, 1448–1461. [Google Scholar] [CrossRef] [Green Version]
- Huhta, V.; Karppinen, E.; Nurminen, M.; Valpas, A. Effect of silvicultural practices upon Arthropos, Annelid and Nematode populations in coniferous forest soil. Ann. Zool. Fenn. 1967, 4, 87–145. [Google Scholar]
- Peck, R.W.; Niwa, C.G. Longer-term effects of selective thinning on microarthropod communities in a late-successional coniferous forest. Environ. Entomol. 2005, 34, 646–655. [Google Scholar] [CrossRef]
- Hokazono, S.; Hayashi, K. Variability in environmental impacts during conversion from conventional to organic farming: A comparison among three rice production systems in Japan. J. Clean. Prod. 2012, 28, 101–112. [Google Scholar] [CrossRef]
- Landi, S.; Papini, R.; d’Errico, G.; Barzanti, G.P.; Roversi, P.F. Nematode indicators as integrative measures of soil condition during conversion from conventional to organic rice production in Italy: A case study. Biol. Agric. Hortic. 2018, 34, 141–153. [Google Scholar] [CrossRef]
- Cantiani, P. Il diradamento selettivo. In Accrescere Stabilità e Biodiversità in Boschi Artificiali di Pino Nero. Manuale Tecnico SelPiBioLife; Compagnia delle Foreste: Arezzo, Italy, 2016. [Google Scholar]
- United States Department of Agriculture. Soil Survey Staff Keys to Soil Taxonomy, 12th ed.; Natural Resources Conservation Service: Washington, DC, USA, 2014.
- Barbato, D.; Perini, C.; Mocali, S.; Bacaro, G.; Tordoni, E.; Maccherini, S.; Marchi, M.; Cantiani, P.; De Meo, I.; Bianchetto, E.; et al. Teamwork makes the dream work: Disentangling cross-taxon congruence across soil biota in black pine plantations. Sci. Total Environ. 2019, 656, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Marchi, M.; Paletto, A.; Cantiani, P.; Bianchetto, E.; De Meo, I. Comparing thinning system effects on ecosystem services provision in artificial black pine (Pinus nigra JF Arnold) forests. Forests 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Cantiani, P.; Marchi, M. A spatial dataset of forest mensuration collected in black pine plantations in central Italy. Ann. For. Sci. 2017, 74, 50. [Google Scholar] [CrossRef]
- Costantini, E.A.C. Linee Guida dei Metodi di rilevamento e Informatizzazione dei dati Pedologici; CRA-ABP: Firenze, Italy, 2007; p. 296. [Google Scholar]
- MiPAAF; Osservatorio Nazionale Pedologico; Marcello Pagliai. Metodi di Analisi Fisica del Suolo; Ministero per le Politiche Agricole e Forestali: Rome, Italy, 1998; p. 400. ISBN 8846404262. [Google Scholar]
- MiPAAF. Official Methods of Soil Chemical Analysis; Gazzetta Ufficiale Supplemento Ordinario No. 248; Istituto Poligrafico e Zecca dello Stato: Rome, Italy, 1999. [Google Scholar]
- Calzolari, C.; Sartori, G.; Magaldi, D. I suoli argillosi a nord di Campiglia d’Orcia (Siena); Quaderni di scienza del suolo, Vol. II-National Research Council: Florence, Italy, 1989. [Google Scholar]
- Gardin, L.; Vinci, A. Carta dei Suoli della Regione Toscana in Scala 1: 250.000. Available online: http://sit.lamma.rete.toscana.it/websuoli/ (accessed on 17 November 2020).
- Reynolds, W.D.; Elrick, D.E. A method for simultaneous in situ measurement in the vadose zone of field-saturated hydraulic conductivity, sorptivity and the conductivity-pressure head relationship. Groundw. Monit. Remediat. 1986, 6, 84–95. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J.; Koike, T.; Van Con, T.; Chavanne, A.; Seifert, T.; et al. Crown size and growing space requirement of common tree species in urban centres, parks, and forests. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Tabacchi, G.; Di Cosmo, L.; Gasparini, P. Aboveground tree volume and phytomass prediction equations for forest species in Italy. Eur. J. For. Res. 2011. [Google Scholar] [CrossRef]
- Mai, W.F. Pictorial Key to Genera of Plant-Parasitic Nematodes. In Nematode Identification and Expert System Technology; Fortuner, R., Ed.; Nato ASI Series (Series A: Life Sciences); Springer: Boston, MA, USA, 1988; Volume 7, pp. 31–34. [Google Scholar]
- Bongers, T. De Nematoden van Nederland: Een Identificatietabel voor de in Nederland Aangetroffen Zoetwater-en Bodembewonende Nematoden; Koninklijke Nederlandse Natuurhistorische Vereniging: Zeist, The Netherlands, 1988. [Google Scholar]
- Marinari-Palmisano, A.; Vinciguerra, M. Classificazione dei nematodi. In Nematologia Agraria Generale e Applicata; Ambrogioni, L., d’Errico, F.P., Greco, N., Marinari-Palmisano, A., Roversi, F.P., Eds.; Soc. Italiana di Nematologia: Bari, Italy, 2014; pp. 23–42. [Google Scholar]
- Yeates, G.W.; Bongers, T.D.; De Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Okada, H.; Harada, H.; Kadota, I. Fungal-feeding habits of six nematode isolates in the genus Filenchus. Soil Biol. Biochem. 2005, 37, 1113–1120. [Google Scholar] [CrossRef]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Bongers, T.; De Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Bedding, R.A.; Akhurst, R.J. A simple technique for the detection of insect paristic rhabditid nematodes in soil. Nematologica 1975, 21, 109–110. [Google Scholar] [CrossRef]
- Landi, S.; Pennacchio, F.; Papini, R.; d’Errico, G.; Torrini, G.; Strangi, A.; Barabaschi, D.; Roversi, P.F. Evaluation of sheep grazing effects on nematode community, insect infestation and soil fertility in sweet chestnut orchards: A case of study. Redia 2016, 99, 117. [Google Scholar]
- Kaya, H.K.; Stock, P. Techniques in insect nematology. In Manual of Techniques in Insect Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 1997; pp. 281–324. [Google Scholar]
- Nguyen, K.; Hunt, D. Entomopathogenic Nematodes: Systematics, Phylogeny and Bacterial Symbionts; Brill: Leiden, The Netherlands, 2007. [Google Scholar]
- Parisi, V.; Menta, C.; Gardi, C.; Jacomini, C.; Mozzanica, E. Microarthropod communities as a tool to assess soil quality and biodiversity: A new approach in Italy. Agric. Ecosyst. Environ. 2005, 105, 323–333. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Past version 1.95: Paleontological Statistical Software Package for Education and Data Analysis. Paleontología Electrónica 2001, 4, 9. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Jiang, Y.; Yin, X.; Wang, F. Composition and Spatial Distribution of Soil Mesofauna along an Elevation Gradient on the North Slope of the Changbai Mountains, China. Pedosphere 2015, 25, 811–824. [Google Scholar] [CrossRef]
- Lazzaro, L.; Mazza, G.; d’Errico, G.; Fabiani, A.; Giuliani, C.; Inghilesi, A.F.; Lagomarsino, A.; Landi, S.; Lastrucci, L.; Pastorelli, R. How ecosystems change following invasion by Robinia pseudoacacia: Insights from soil chemical properties and soil microbial, nematode, microarthropod and plant communities. Sci. Total Environ. 2018, 622, 1509–1518. [Google Scholar] [CrossRef]
- Bird, G.A.; Chatarpaul, L. Effect of forest harvest on decomposition and colonization of maple leaf litter by soil microarthropods. Can. J. Soil Sci. 1988, 68, 29–40. [Google Scholar] [CrossRef]
- Becagli, C.; Puletti, N.; Chiavetta, U.; Cantiani, P.; Salvati, L.; Fabbio, G. Early impact of alternative thinning approaches on structure diversity and complexity at stand level in two beech forests in Italy. Ann. Silvic. Res. 2013, 37, 55–63. [Google Scholar]
- Di Salvatore, U.; Tonti, D.; Chiavetta, U.; Cantiani, P.; Fabbio, G.; Becagli, C.; Bertini, G.; Sansone, D.; Skudnik, M.; Kobal, M. ManFor C.BD sites and the drivers of forest functions. Ital. J. Agron. 2016, 11, 64–95. [Google Scholar]
- Forge, T.; Simard, S. Structure of nematode communities in forest soils of southern British Columbia: Relationships to nitrogen mineralization and effects of clearcut harvesting and fertilization. Biol. Fertil. Soils 2001, 34, 170–178. [Google Scholar]
- Sohlenius, B. Influence of clear-cutting and forest age on the nematode fauna in a Swedish pine forest soil. Appl. Soil Ecol. 2002, 19, 261–277. [Google Scholar] [CrossRef]
- Tarasco, E.; Clausi, M.; Rappazzo, G.; Panzavolta, T.; Curto, G.; Sorino, R.; Oreste, M.; Longo, A.; Leone, D.; Tiberi, R. Biodiversity of entomopathogenic nematodes in Italy. J. Helminthol. 2014, 89, 359–366. [Google Scholar] [CrossRef]
- De Luca, F.; Clausi, M.; Troccoli, A.; Curto, G.; Rappazzo, G.; Tarasco, E. Entomopathogenic nematodes in Italy: Occurrence and use in microbial control strategies. In Nematode Pathogenesis of Insects and Other Pests; Springer: Berlin/Heidelberg, Germany, 2015; pp. 431–449. [Google Scholar]
- Bird, S.; Coulson, R.N.; Crossley, D.A., Jr. Impacts of silvicultural practices on soil and litter arthropod diversity in a Texas pine plantation. For. Ecol. Manag. 2000, 131, 65–80. [Google Scholar] [CrossRef]
- Landi, S.; d’Errico, G.; Simoncini, S.; L’Abate, G.; Priori, S. Nematode communities as indicators of soil quality in vineyard system: A case of study in degraded areas. EQA-Int. J. Environ. Qual. 2018, 31, 41–46. [Google Scholar] [CrossRef]
- Mackay, W.P.; Silva, S.; Lightfoot, D.C.; Pagani, M.I.; Whitford, W.G. Effect of increased soil moisture and reduced soil temperature on a desert soil arthropod community. Am. Midl. Nat. 1986, 116, 45–56. [Google Scholar] [CrossRef]
- Baujard, P.; Martiny, B. Ecology and pathogenicity of the Hoplolaimidae (Nemata) from the sahelian zone of West Africa. 2. Laboratory studies on Scutellonema cavenessi Sher, 1964. Fundam. Appl. Nematol. 1995, 18, 335–346. [Google Scholar]
- Špaldoňová, A.; Frouz, J. Decomposition of Forest Litter and Feces of Armadillidium vulgare (Isopoda: Oniscidea) Produced from the Same Litter Affected by Temperature and Litter Quality. Forests 2019, 10, 939. [Google Scholar] [CrossRef] [Green Version]
- Throne, J.E.; Flinn, P.W. Distribution of psocids (Psocoptera) in temperature gradients in stored wheat. J. Stored Prod. Res. 2013, 55, 27–31. [Google Scholar] [CrossRef]
- Villarreal, E.; Martínez, N.; Ortiz, C.R. Diversity of Pseudoscorpiones (Arthropoda: Arachnida) in two fragments of dry tropical forest in the colombian Caribbean region. Caldasia 2019, 41, 139–151. [Google Scholar] [CrossRef]
- Edwards, C.A. The ecology of symphyla part III. Factors controlling soil distributions. Entomologia Experimentalis et Applicata 1961, 4, 239–256. [Google Scholar] [CrossRef]
- Mitrovski-Bogdanovic, A.; Blesic, B. Seasonal dynamics of Campodeidae (Diplura) in an oak forest in Kragujevac (Serbia). Acta Entomologica Serbica 2007, 12, 1–10. [Google Scholar]
- Shakir, M.M.; Ahmed, S. Seasonal abundance of soil arthropods in relation to meteorological and edaphic factors in the agroecosystems of Faisalabad, Punjab, Pakistan. Int. J. Biometeorol. 2015, 59, 605–616. [Google Scholar] [CrossRef]
- Tsafack, N.; Xie, Y.; Wang, X.; Fattorini, S. Influence of Climate and Local Habitat Characteristics on Carabid Beetle Abundance and Diversity in Northern Chinese Steppes. Insects 2020, 11, 19. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Treatment | N. Trees per ha | Basal Area | Standing Volume | Quadratic Mean Diameter | Mean Height | Crown Volume | PAR |
|---|---|---|---|---|---|---|---|---|
| Pratomagno | Thinning from below | −35.9% (*) | −22.6% (*) | −19.3% (*) | +9.55% | −4.18% | −11.3% (*) | +87% |
| Selective thinning | −30.8% (*) | −29.4% (*) | −29.7% (*) | 0.00% | +0.53% | −21.2% (*) | +232% | |
| Amiata | Thinning from below | −30.4% (*) | −19.7% (*) | −18.7% (*) | −6.75% | +2.23% | −24.4% (*) | +133% |
| Selective thinning | −34.3% (*) | −31.9% (*) | −30.7% (*) | +2.00% | +1.09% | −11.2% (*) | +413% |
| Year | Nematodes | Microarthropods | ||||||
|---|---|---|---|---|---|---|---|---|
| Pratomagno | Amiata | Pratomagno | Amiata | |||||
| R | p | R | p | R | p | R | p | |
| 2016 | 0.36 | 0.04 | −0.23 | 0.86 | 0.03 | 0.20 | 0.05 | 0.13 |
| 2017 | 0.16 | 0.37 | 0.13 | 0.24 | 0.11 | 0.02 | 0.07 | 0.05 |
| 2018 | 0.17 | 0.05 | −0.01 | 0.47 | −0.03 | 0.68 | −0.04 | 0.86 |
| Management | Year | Significant Effect | |||||||
|---|---|---|---|---|---|---|---|---|---|
| TB | ST | CTR | 2016 | 2017 | 2018 | M | Y | M + Y | |
| Pratomagno | |||||||||
| MI | 2.1 ± 0.2 | 1.9 ± 0.1 | 1.9 ± 0.2 | 2.3 ± 0.1 a | 1.6 ± 0.1 b | 2.1 ± 0.1 ab | 0.36 | 0.002 | 0.31 |
| PPI | 2.7 ± 0.3 a | 2.2 ± 0.1 b | 2.7 ± 0.3 a | 3.1 ± 0.3 a | 2.4 ± 0.04 b | 2.1 ± 0.1 b | 0.02 | 0.0005 | 0.02 |
| EI | 89.4 ± 2.0 | 93.6 ± 1.5 | 92.0 ± 2.6 | 93.9 ± 1.4 a | 96.1 ± 1.0 a | 85.0 ± 1.5 b | 0.10 | 0.00001 | 0.56 |
| SI | 83.5 ± 4.5 | 74.8 ± 10.4 | 79.7 ± 6.0 | 91.1 ± 2.6 | 77.5 ± 5.8 | 69.4 ± 10.0 | 0.70 | 0.65 | 0.65 |
| CI | 8.0 ± 1.9 | 6.0 ± 1.6 | 7.7 ± 2.6 | 4.1 ± 0.8 b | 3.4 ± 0.8 b | 14.1 ± 1.8 a | 0.50 | 0.00001 | 0.67 |
| H | 1.5 ± 0.1 | 1.3 ± 0.1 | 1.2 ± 0.2 | 1.5 ± 0.1 a | 1.0 ± 0.2 b | 1.5 ± 0.1 a | 0.16 | 0.01 | 0.72 |
| D | 0.3 ± 0.04 | 0.4 ± 0.04 | 0.4 ± 0.05 | 0.3 ± 0.02 b | 0.5 ± 0.04 a | 0.3 ± 0.02 b | 0.18 | 0.0006 | 0.90 |
| Amiata | |||||||||
| MI | 2.0 ± 0.2 | 2.1 ± 0.2 | 2.2 ± 0.1 | 2.3 ± 0.1 a | 1.7 ± 0.1 b | 2.4 ± 0.1 a | 0.56 | 0.004 | 0.77 |
| PPI | 2.4 ± 0.2 | 2.1 ± 0.05 | 2.2 ± 0.1 | 2.3 ± 0.1 | 2.3 ± 0.1 | 2.2 ± 0.2 | 0.25 | 0.96 | 0.96 |
| EI | 88.1 ± 3.3 a | 79.9 ± 5.3 ab | 74.0 ± 6.8 b | 82.4 ± 3.7 a | 93.4 ± 2.2 a | 66.2 ± 7.0 b | 0.05 | 0.0003 | 0.04 |
| SI | 78.8 ± 4.8 | 70.6 ± 5.3 | 73.6 ± 6.8 | 74.9 ± 6.5 | 76.6 ± 6.5 | 71.4 ± 3.8 | 0.64 | 0.83 | 0.68 |
| CI | 11.4 ± 3.3 b | 18.2 ± 4.5 | 24.9 ± 7.4 a | 17.6 ± 3.7 b | 5.9 ± 2.1 c | 31.0 ± 6.3 a | 0.04 | 0.0002 | 0.02 |
| H | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.1 | 1.3 ± 0.03 a | 1.1 ± 0.04 b | 1.4 ± 0.1 a | 0.79 | 0.002 | 0.55 |
| D | 0.4 ± 0.04 | 0.3 ± 0.03 | 0.4 ± 0.03 | 0.4 ± 0.02 | 0.5 ± 0.03 | 0.3 ± 0.03 | 0.33 | 0.001 | 0.03 |
| Management | Year | Significant Effect | |||||||
|---|---|---|---|---|---|---|---|---|---|
| TB | ST | CTR | 2016 | 2017 | 2018 | M | Y | M + Y | |
| Pratomagno | |||||||||
| QBS-ar | 99.8 ± 7.6 a | 82.0 ± 2.9 b | 88.9 ± 7.4 | 79.3 ± 16.6 b | 101.0 ± 5.4 a | 90.3 ± 7.5 | 0.05 | 0.03 | 0.06 |
| H | 1.3 ± 0.04 a | 1.1 ± 0.1 | 1.0 ± 0.1 b | 1.3 ± 0.1 a | 1.2 ± 0.05 b | 0.9 ± 0.1 b | 0.02 | 0.01 | 0.16 |
| D | 0.3 ± 0.02 b | 0.4 ± 0.1 | 0.5 ± 0.05 a | 0.4 ± 0.05 b | 0.4 ± 0.03 b | 0.5 ± 0.05 a | 0.04 | 0.02 | 0.36 |
| Amiata | |||||||||
| QBS-ar | 123.0 ± 8.2 | 123.8 ± 4.4 | 136.8 ± 9.7 | 127.7 ± 5.3 | 144.2 ± 7.7 a | 111.7 ± 6.4 b | 0.26 | 0.008 | 0.42 |
| H | 1.3 ± 0.1 a | 1.1 ± 0.1 b | 1.2 ± 0.05 | 1.3 ± 0.1 a | 1.1 ± 0.03 b | 1.1 ± 0.1 b | 0.02 | 0.01 | 0.16 |
| D | 0.3 ± 0.03 b | 0.5 ± 0.03 a | 0.4 ± 0.05 | 0.3 ± 0.05 b | 0.5 ± 0.02 a | 0.4 ± 0.04 a | 0.03 | 0.02 | 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landi, S.; d’Errico, G.; Binazzi, F.; Di Salvatore, U.; Gardin, L.; Marchi, M.; Mazza, G.; Roversi, P.F.; Simoncini, S.; Torrini, G.; et al. The Short-Term Impact of Different Silvicultural Thinnings on Soil Nematode and Microarthropod Biodiversity in Artificial Black Pine Stands. Forests 2020, 11, 1212. https://doi.org/10.3390/f11111212
Landi S, d’Errico G, Binazzi F, Di Salvatore U, Gardin L, Marchi M, Mazza G, Roversi PF, Simoncini S, Torrini G, et al. The Short-Term Impact of Different Silvicultural Thinnings on Soil Nematode and Microarthropod Biodiversity in Artificial Black Pine Stands. Forests. 2020; 11(11):1212. https://doi.org/10.3390/f11111212
Chicago/Turabian StyleLandi, Silvia, Giada d’Errico, Francesco Binazzi, Umberto Di Salvatore, Lorenzo Gardin, Maurizio Marchi, Giuseppe Mazza, Pio Federico Roversi, Stefania Simoncini, Giulia Torrini, and et al. 2020. "The Short-Term Impact of Different Silvicultural Thinnings on Soil Nematode and Microarthropod Biodiversity in Artificial Black Pine Stands" Forests 11, no. 11: 1212. https://doi.org/10.3390/f11111212