The First Record of a North American Poplar Leaf Rust Fungus, Melampsora medusae, in China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Surveys and Specimens
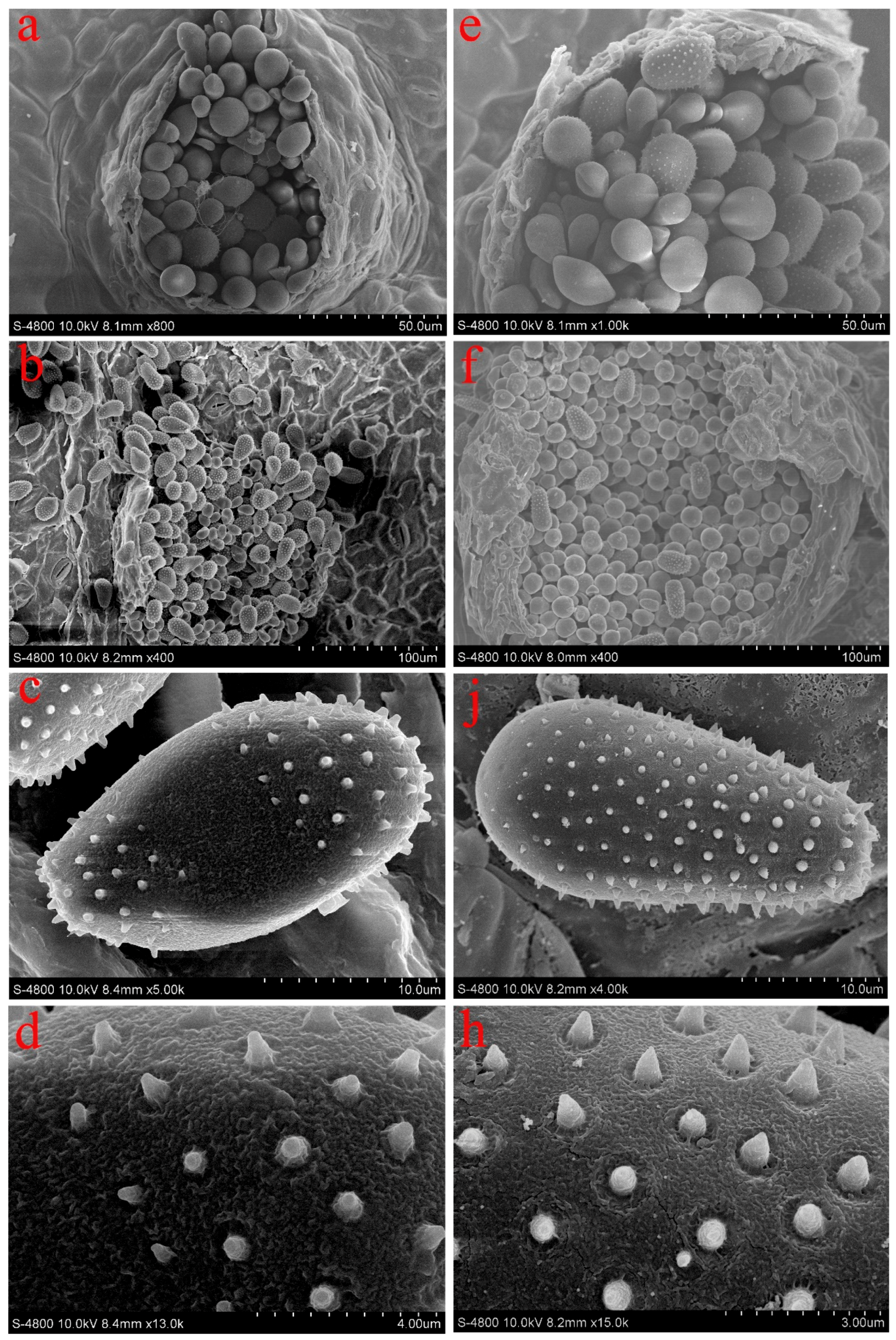
2.2. Morphological Observations
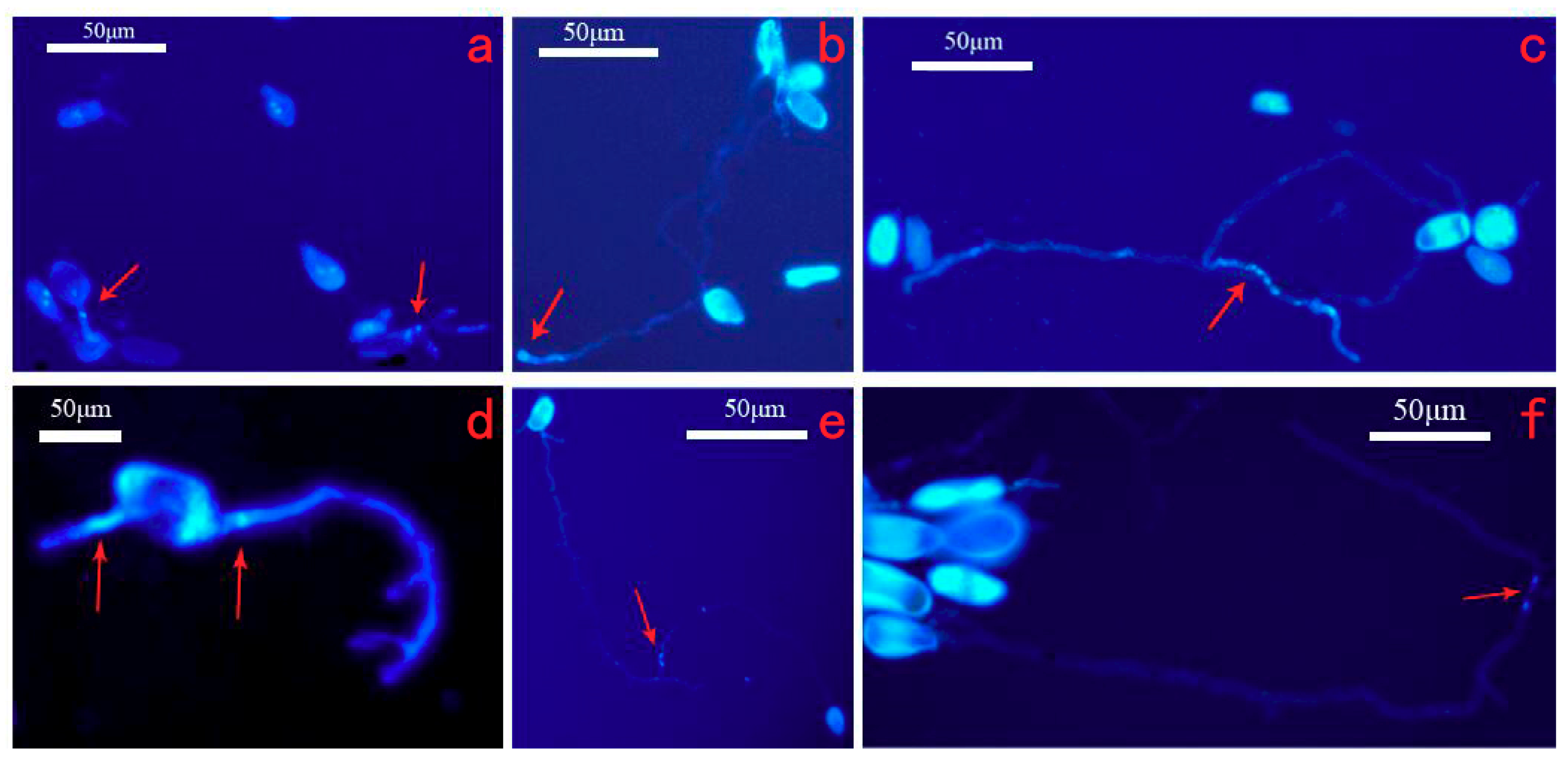
2.3. Germination of Urediospores on the Surface of 2% Agar
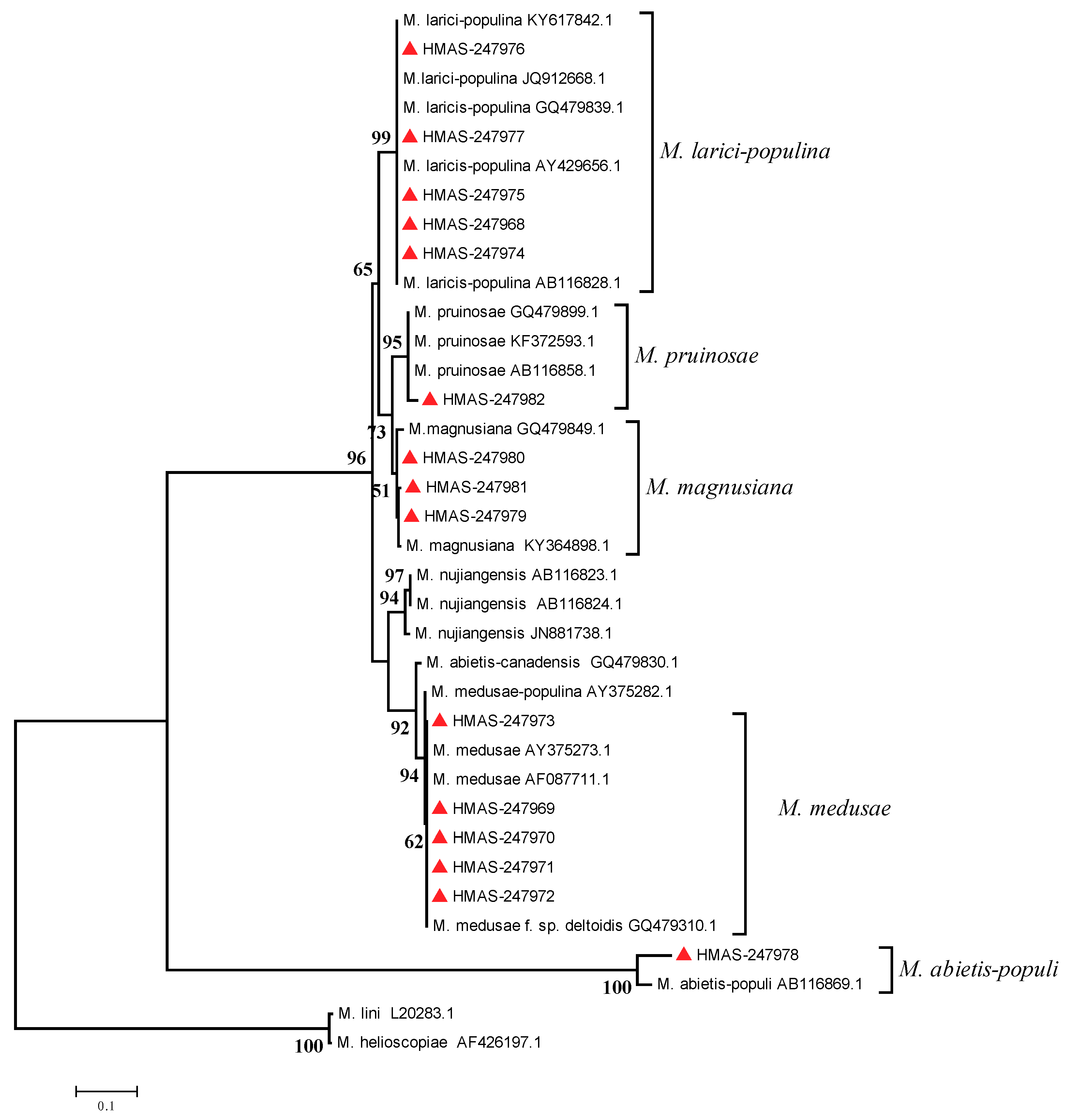
2.4. Phylogenetic Analysis
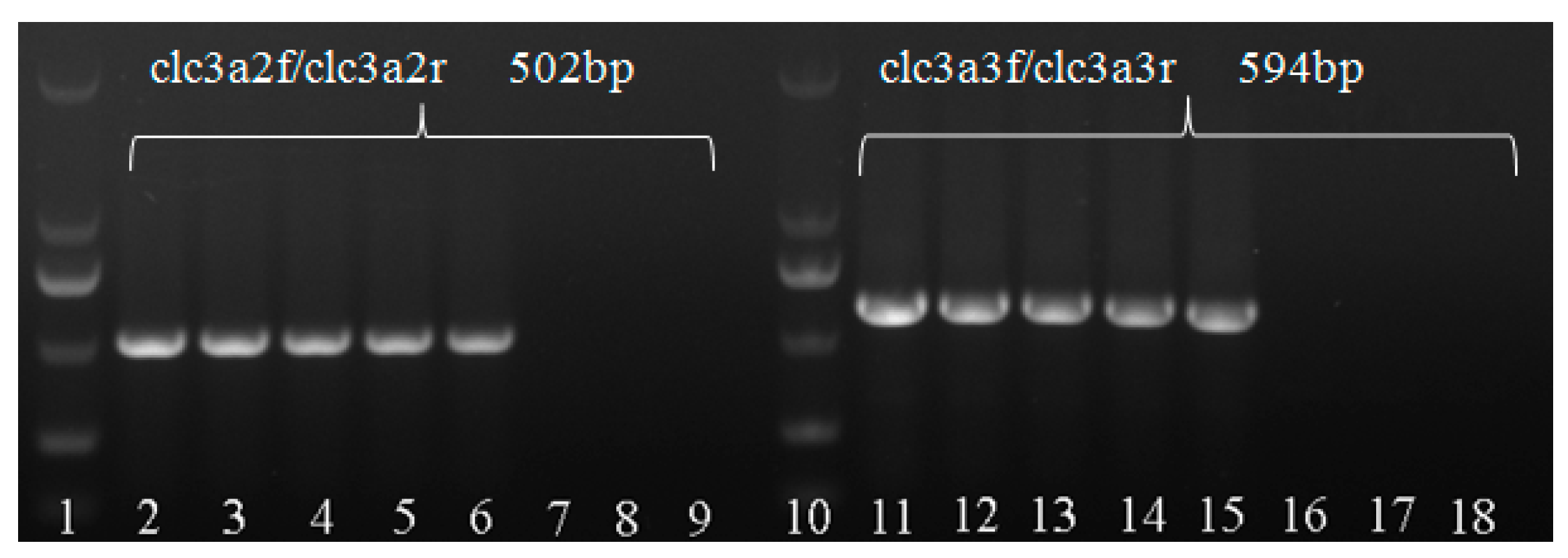
2.5. Confirmation of M. medusae
3. Results
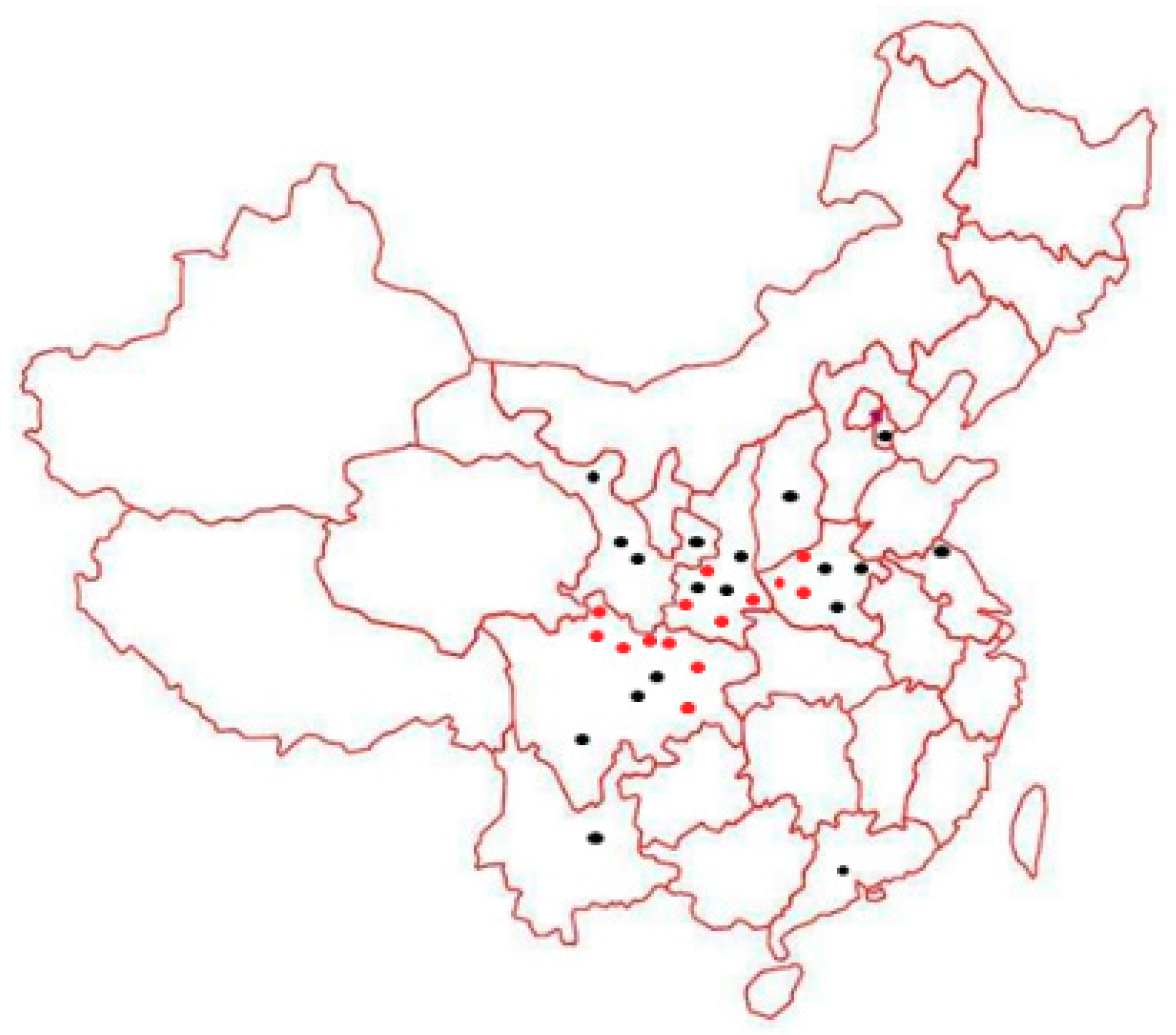
3.1. Surveyed Poplar Leaf Rust in China, 2015–2018
3.2. Germination of Urediniospores on 2% Agar
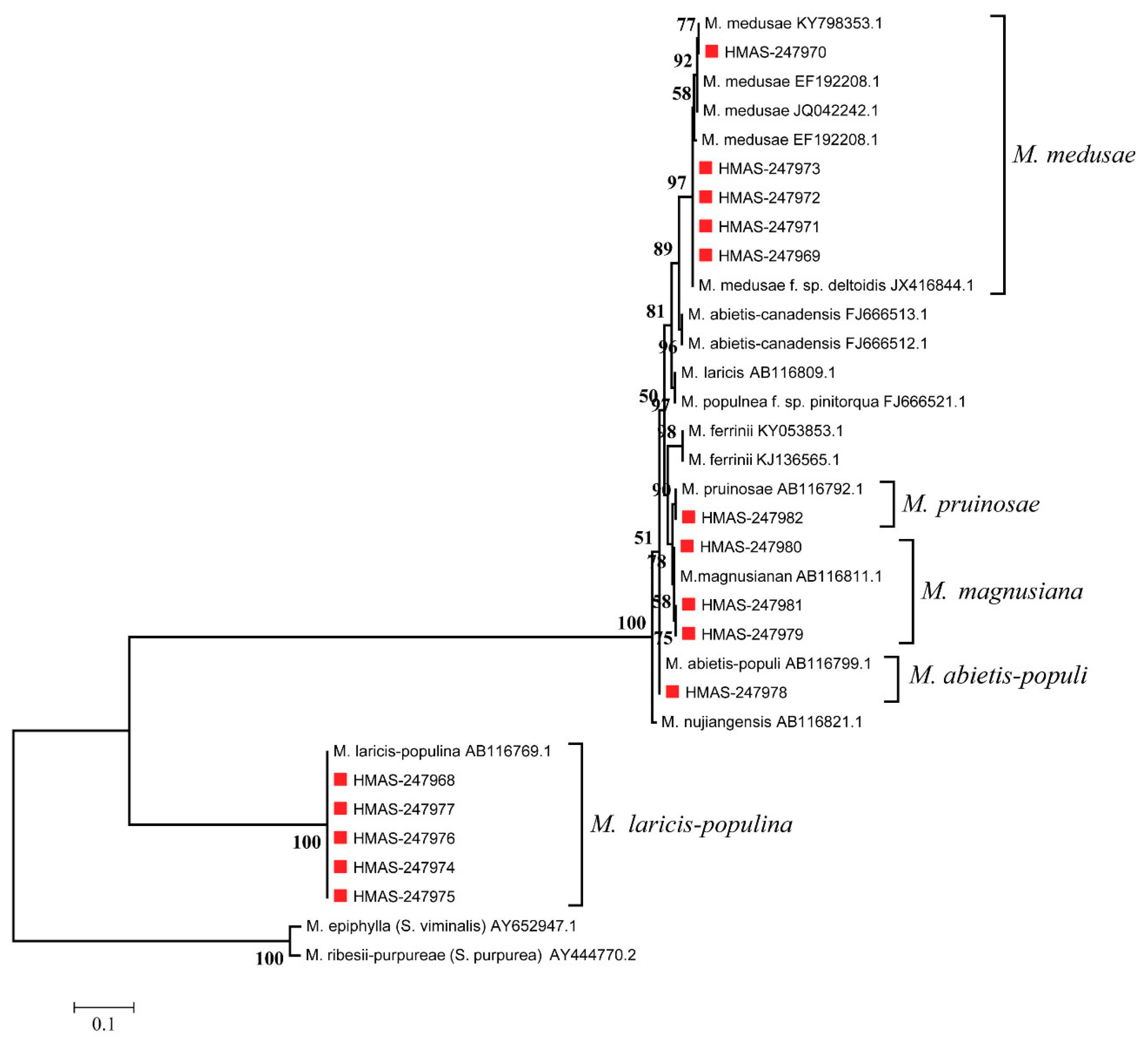
3.3. Molecular Phylogenetic Analysis
3.4. Formae Speciales Identification
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pinon, J.; Frey, P. Interaction between poplar clones and Melampsora populations and their implication for breeding for durable resistance. In Rust Diseases of Willow and Poplar; CABI: Oxford, England, 2005; Volume 12, pp. 139–154. [Google Scholar]
- Tian, C.M.; Shang, Y.Z.; Zhuang, J.Y.; Wang, Q.; Kakishima, M. Morphological and molecular phylogenetic analysis of Melampsora species on poplars in China. Mycoscience 2004, 45, 55–66. [Google Scholar] [CrossRef]
- Steenackers, J.; Steenackers, M.; Steenackers, V.; Stevens, M. Poplar diseases, consequences on growth and wood quality. Biomass Bioenerg. 1996, 10, 267–274. [Google Scholar] [CrossRef]
- Galović, V.; Orlović, S.; Pap, P.; Kovačević, B.; Marković, M. Specificity of SSR Loci for Melampsora Species on Poplars. Genetika-Belgrade 2010, 42, 513–520. [Google Scholar] [CrossRef]
- Newcombe, G.; Dugan, F.M. Fungal pathogens of plants in the Homogocene. In Molecular Identification of Fungi; Springer: Berlin, Heidelberg, Germany, 2010; pp. 3–34. [Google Scholar]
- Castagne, L. Observations sur quelques plantes acotylédonées de la famille des uredinées. In recueillies dans le département des Bouches-du-Rhône; impr. de Achard: Marseille, France, 1843. [Google Scholar]
- Vialle, A.; Frey, P.; Hambleton, S. Poplar rust systematics and refinement of Melampsora species delineation. Fungal Divers. 2011, 50, 227–248. [Google Scholar] [CrossRef]
- Newcombe, G.; Stirling, B.; Mcdonald, S.; Bradshaw, H.D. Melampsora × columbiana, a natural hybrid of M. medusae and M. occidentalis. Mycol. Res. 2000, 104, 261–274. [Google Scholar] [CrossRef]
- Spiers, A.G.; Hopcroft, D.H. Comparative studies of the poplar rusts Melampsora medusae, M. larici-populina and their interspecific hybrid M. medusae-populina. Mycol. Res. 1994, 98, 889–903. [Google Scholar] [CrossRef]
- Thümen, F.v. New species of North American Uredinei. Bull. Torrey Bot. Club 1878, 6, 215–216. [Google Scholar] [CrossRef]
- Farr, D.F.; Rossman, A.Y. Fungal Databases; U.S. National Fungus Collections, ARS, USDA: Washington, DC, USA, 6 December 2018. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 25 October 1984).
- Fresa, R. Argentine Republic: Melampsora larici-populina in the Delta of Paraná. Int. Bull. Pl. Prot. 1936, 10, 145–146. [Google Scholar]
- Dupias, G. Contribution à l'étude des Urédinées de la Haute-Garonne. Bull. Soc. Hist.Nat. Toulouse. 1943, 78, 32–52. [Google Scholar]
- Hiratsuka, N. Miscellaneous notes on the East Asiatic Uredinales with special reference to the Japanese species (VI). J. Jap. Bot. 1939, 15, 621–627. [Google Scholar]
- Hennebert, G.L. L'identification des rouilles du peuplier. Agricultura 1964, 12, 661–670. [Google Scholar]
- Pinon, J. Situation of Melampsora medusae in Europe. Bull 1986, 16, 547–551. [Google Scholar] [CrossRef]
- Pinon, J. Eléments de répartition des rouilles des peupliers cultivés en France. C. R. Acad. Agric. Fr. 1991, 77, 109–115. [Google Scholar]
- OEPP/EPPO. Data sheets on quarantine organisms no. 33, Melampsora medusae. Bull 1982, 12, 6. [Google Scholar]
- Husson, C.; Loos, R.; Andrieux, A.; Frey, P. Development and use of new sensitive molecular tools for diagnosis and detection of Melampsora rusts on cultivated poplar. For. Pathol. 2013, 43, 1–11. [Google Scholar] [CrossRef]
- Walker, J.; Hartigan, D.; Bertus, A.L. Poplar rusts in Australia with comments on potential conifer rusts. Eur. J. Plant Pathol. 1974, 4, 100–118. [Google Scholar] [CrossRef]
- Council of the European Union. Council Directive 2000/29/EC of 8 May 2000 on Protective Measures against the Introduction into the Community of Organisms Harmful to Plants or Plant Products and against their Spread within the Community. O.J.L.. 2000, Volume 169. Available online: https://publications.europa.eu/en/publication-detail/-/publication/6aab39f3-60ec-4851-99cf-d676e093b8a7/language-en (accessed on 28 June 2007).
- Feau, N.; Vialle, A.; Allaire, M.; Tanguay, P.; Joly, D.L.; Frey, P.; Callan, B.E.; Hamelin, R.C. Fungal pathogen (mis-) identifications: A case study with DNA barcodes on Melampsora rusts of aspen and white poplar. Mycol. Res. 2009, 113, 713–724. [Google Scholar] [CrossRef]
- Boutigny, A.L.; Guinet, C.; Vialle, A.; Hamelin, R.; Frey, P.; Ioos, R. A sensitive real-time PCR assay for the detection of the two Melampsora medusae formae speciales on infected poplar leaves. Eur. J. Plant Pathol. 2013, 136, 433–441. [Google Scholar] [CrossRef]
- Pinon, J.; Frey, P. Structure of Melampsora larici-populina populations on wild and cultivated poplar. Eur. J. Plant Pathol. 1997, 103, 159–173. [Google Scholar] [CrossRef]
- Nakamura, H.; Kaneko, S.; Yamaoka, Y.; Kakishima, M. Differentiation of Melampsora rust species on willows in Japan using PCR-RFLP analysis of ITS regions of ribosomal DNA. Mycoscience 1998, 39, 105–113. [Google Scholar] [CrossRef]
- Pei, M.H.; Bayon, C.; Ruiz, C. Phylogenetic relationships in some Melampsora rusts on Salicaceae assessed using rDNA sequence information. Mycol. Res. 2005, 109, 401–409. [Google Scholar] [CrossRef]
- Zhao, P.; Tian, C.M.; Yao, Y.J.; Wang, Q.; Kakishima, M.; Yamaoka, Y. Melampsora salicis-sinicae (Melampsoraceae, Pucciniales), a new rust fungus found on willows in China. Mycoscience 2014, 55, 390–399. [Google Scholar] [CrossRef]
- Heath, M.C.; Xu, H.X.; Eilam, T. Nuclear behavior of the cowpea rust fungus during the early stages of basidiospore- or urediospore- derived growth in resistant or susceptible cowpea cultivars. Phytopathology 1996, 86, 1057–1065. [Google Scholar] [CrossRef]
- Yu, Z.D.; Liang, J.; Cao, Z.M. Nuclear behavior in the life cycle of Melompsor larici-populina Kleb. J. Food Agric. Environ. 2009, 7, 165–169. [Google Scholar] [CrossRef]
- Virtudazo, E.V.; Nakamura, H.; Kakishima, M. Phylogenetic analysis of sugarcane rusts based on sequences of ITS, 5.8 S rDNA and D1/D2 regions of LSU rDNA. J. Gen. Plant Pathol. 2001, 67, 28–36. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes: application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequence of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K. Fusarium and its near relatives. In The Fungal Holomorph: Mitotic, Meiotic and Pleomophic Speciation in Fungal Systematics; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP 4.0: Phylogenetic Analysis using Parsimony, version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar] [CrossRef]
- Bourassa, M.; Bernier, L.; Hamelin, R.C. Direct genotyping of the poplar leaf rust fungus, Melampsora medusae f. sp. deltoidae, using codominant PCR-SSCP markers. Forest Pathol. 2005, 35, 245–261. [Google Scholar] [CrossRef]
- Maier, W.; Begerow, D.; Weiß, M.; Oberwinkler, F. Phylogeny of the rust fungi: an approach using nuclear large subunit ribosomal DNA sequences. Can. J. Bot. 2003, 81, 12–23. [Google Scholar] [CrossRef]
- Ihrmark, K.; Bodeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region—evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef]
- Innes, L.; Marchand, L.; Frey, P.; Bourassa, M.; Hamelin, R.C. First report of Melampsora larici-populina on Populus spp. in eastern North America. Plant Dis. 2004, 88, 85. [Google Scholar] [CrossRef]
- Giordano, L.; Giorcelli, A.; Gonthier, P.; Gullino, M.L. First report of leaf rust caused by Melampsora magnusiana on Populus alba in Italy. J. Plant Pathol. 2017, 99, 535. [Google Scholar]
- Gadgil, P.D. Fungi on trees and shrubs in New Zealand. In Fungi of New Zealand; Fungal Diversity Press: Hong Kong, China, 2005; Volume 4, p. 437. [Google Scholar]
- Newcombe, G.; Chastagner, G.A.; McDonald, S.K. Additional coniferous aecial hosts of the poplar leaf rusts, Melampsora larici-populina and M. medusae f. sp. deltoidae. Plant Dis. 1994, 78, 12–18. [Google Scholar]
- Lin, C.P.; Huang, J.P.; Wu, C.S.; Hsu, C.Y.; Chaw, S.M. Comparative chloroplast genomics reveals the evolution of Pinaceae genera and subfamilies. Genome Biol. Evol. 2010, 2, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.M.; Lin, Y.R.; Huang, H.Y.; Hou, C.L. A new species of Lophodermium associated with the needle cast of Cathay silver fir. Mycol. Prog. 2013, 12, 141–149. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Plants | Locality a/Year | Voucher Specimen no. b | GenBank Accession Accession no. | Species | |
|---|---|---|---|---|---|
| ITS | D1/D2 | ||||
| P. szechuanica | Luding Sichuan/2016 | HMAS247968 | MK028576 | MK064523 | M. larici-populina |
| P. yunnanensis | Jinyang Sichuan/2016 | HMAS 247969 | MK028588 | MK064536 | M. medusae |
| P. simonii | Xichang Sichuan/2017 | HMAS 247970 | MK028589 | MK064537 | M. medusae |
| P. szechuanica | Kangding Sichuan/2018 | HMAS 247971 | MK028590 | MK064535 | M. medusae |
| P. cathayana | Wutaishan Shanxi/2018 | HMAS 247974 | MK028577 | MK064526 | M. larici-populina |
| P. cathayana | Liupanshan Ningxia/2017 | HMAS 247975 | MK028578 | MK064527 | M. larici-populina |
| P. wilsonii | Qinling Shaanxi/2018 | HMAS 247978 | MK028579 | MK064529 | M. abietis-populi |
| P. simonii | Huangyuan Qinghai/2018 | HMAS 247976 | MK028584 | MK064525 | M. larici-populina |
| P. simonii | Haixi Qinghai/2018 | HMAS 247977 | MK028583 | MK064524 | M. larici-populina |
| P. alba var. pyramidalis | Yanqing Beijing/2018 | HMAS 247979 | MK028581 | MK064531 | M. magnusiana |
| P. alba var. pyramidalis | Yulin Shaanxi/2018 | HMAS 247980 | MK028580 | MK064532 | M. magnusiana |
| P. tomentosa | Tianshui Gansu/2018 | HMAS 247981 | MK028582 | MK064530 | M. magnusiana |
| P. euphratica | Inner Mongolia/2018 | HMAS 247982 | MK028585 | MK064533 | M. pruinosae |
| P. deltoides | Linyou Shaanxi/2017 | HMAS 247973 | MK028586 | MK064528 | M. medusae |
| P. deltoides | Yihai Sichuan/2015 | HMAS 247972 | MK028587 | MK064534 | M. medusae |
| Primer Name | Primer Sequence | Annealing Temperature (℃) | Size of Amplified Product (bp) | Sampling Locus/Year |
|---|---|---|---|---|
| clc3a2f | 5′-GGGGGTCTTTAGGACAAA-3′ | 54 | 502 | Shaanxi/2017,2018 Sichuan/2016 Henan/2015,2018 Qinhai/2018 Ningxia/2017 |
| clc3a2r | 5′-TTCGAGCCAGCATGA AACAC-3′ | |||
| clc3a3f | 5′-TTCGAGCCAGAAGTTTGTTTC-3′ | 52 | 594 | |
| clc3a3r | 5′-TTCGAGCCAGGATCACTT-3′ |
| Species | Urediniospores | |||
|---|---|---|---|---|
| Shape | Size (average) (μm) | Wall Equatorial Part Thickness (average) (μm) | Bald Spot | |
| M. larici-populina | Ellipsoid or Oblong | 35.13–48.11 × 20.02–25.31 (40.93) × (22.70) | 3.40–11.63 (5.60) | Apical |
| M. medusae | Obovate or Oval | 20.64–31.45 × 14.39–20.38 (26.56) × (18.66) | 1.76–3.62 (3.14) | Equatorial |
| M. magnusiana | Globose or Ovate | 17.02–29.34 × 14.16–24.22 (23.93) × (18.70) | 1.20–4.65 (2.48) | Absent |
| M. pruinosae | Globose or Ellipsoid | 23.18–31.11 × 23.59–28.53 (27.88) × (25.87) | 0.84–1.57 (1.26) | Absent |
| M. abietis-populi | Globose or Ovate | 20.77–26.34 × 15.14–21.52 (22.92) × (17.93) | 1.14–1.83 (1.59) | Absent |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, W.; Newcombe, G.; Hu, D.; Cao, Z.; Yu, Z.; Peng, Z. The First Record of a North American Poplar Leaf Rust Fungus, Melampsora medusae, in China. Forests 2019, 10, 182. https://doi.org/10.3390/f10020182
Zheng W, Newcombe G, Hu D, Cao Z, Yu Z, Peng Z. The First Record of a North American Poplar Leaf Rust Fungus, Melampsora medusae, in China. Forests. 2019; 10(2):182. https://doi.org/10.3390/f10020182
Chicago/Turabian StyleZheng, Wei, George Newcombe, Die Hu, Zhimin Cao, Zhongdong Yu, and Zijia Peng. 2019. "The First Record of a North American Poplar Leaf Rust Fungus, Melampsora medusae, in China" Forests 10, no. 2: 182. https://doi.org/10.3390/f10020182