Effects of Prescribed Fire, Site Factors, and Seed Sources on the Spread of Invasive Triadica sebifera in a Fire-Managed Coastal Landscape in Southeastern Mississippi, USA

 , ,
, ,
Abstract
:1. Introduction
2. Methods
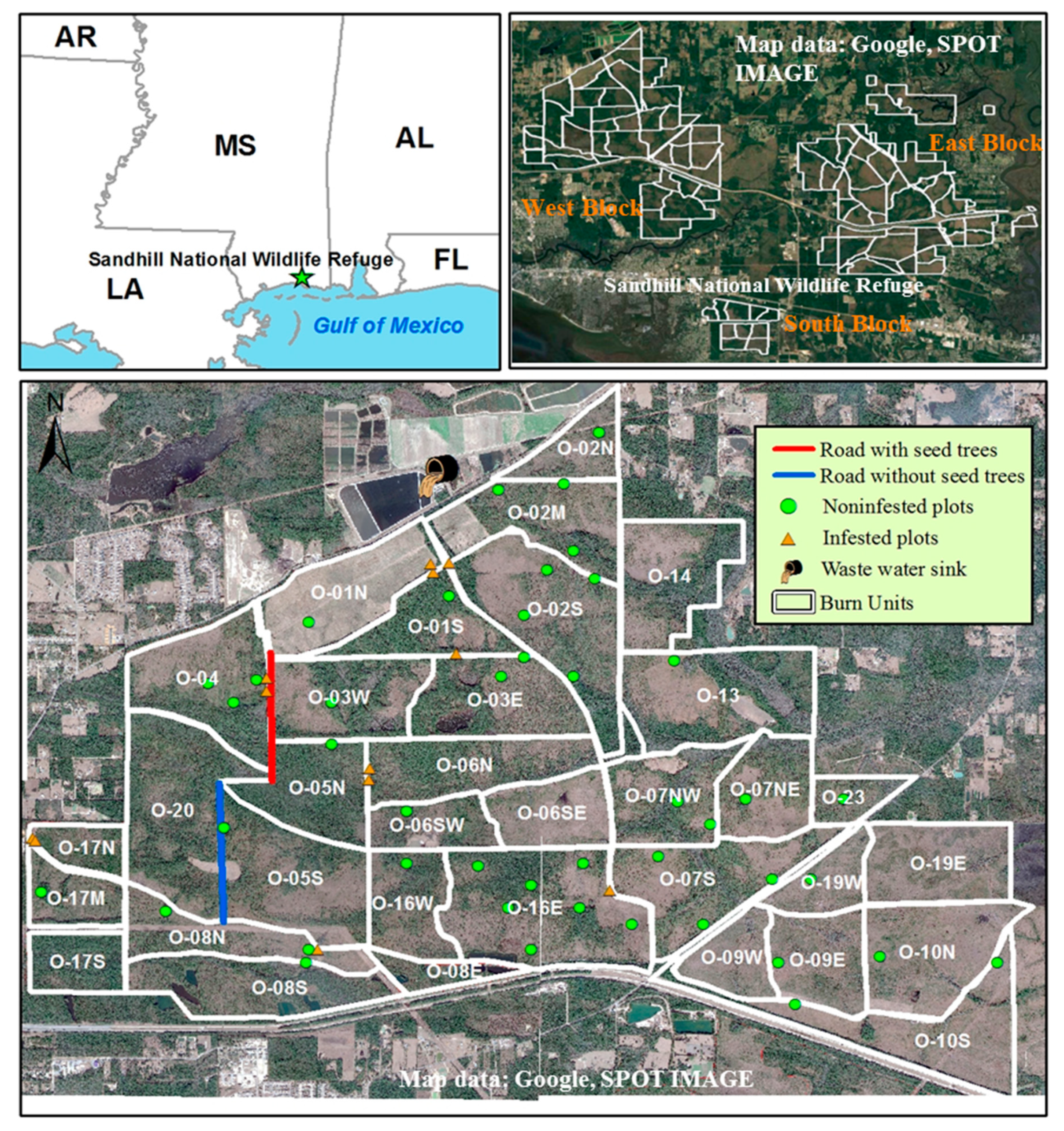
2.1. Research Site
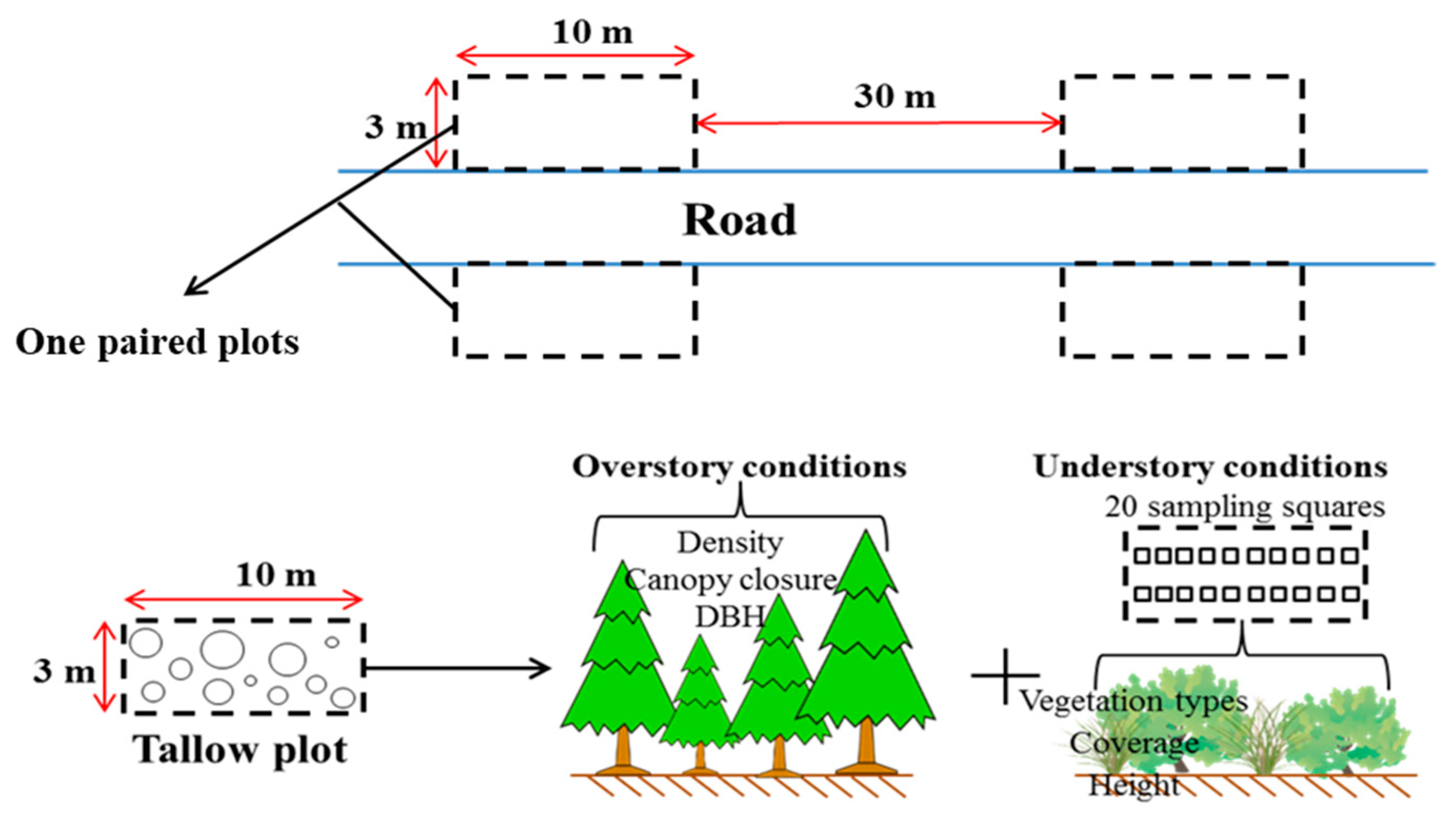
2.2. Data Collection
2.3. Spatial Data Analysis and Modeling
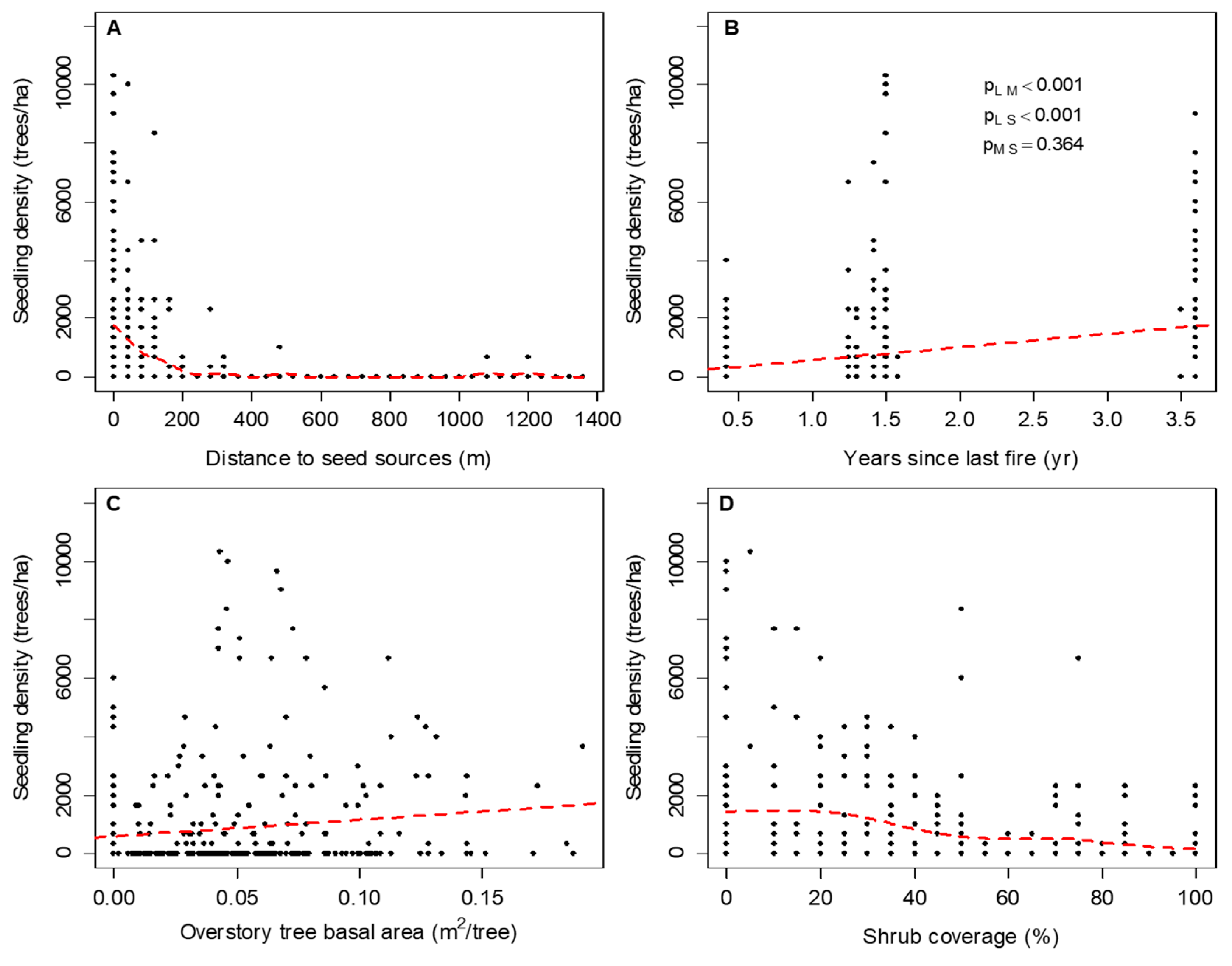
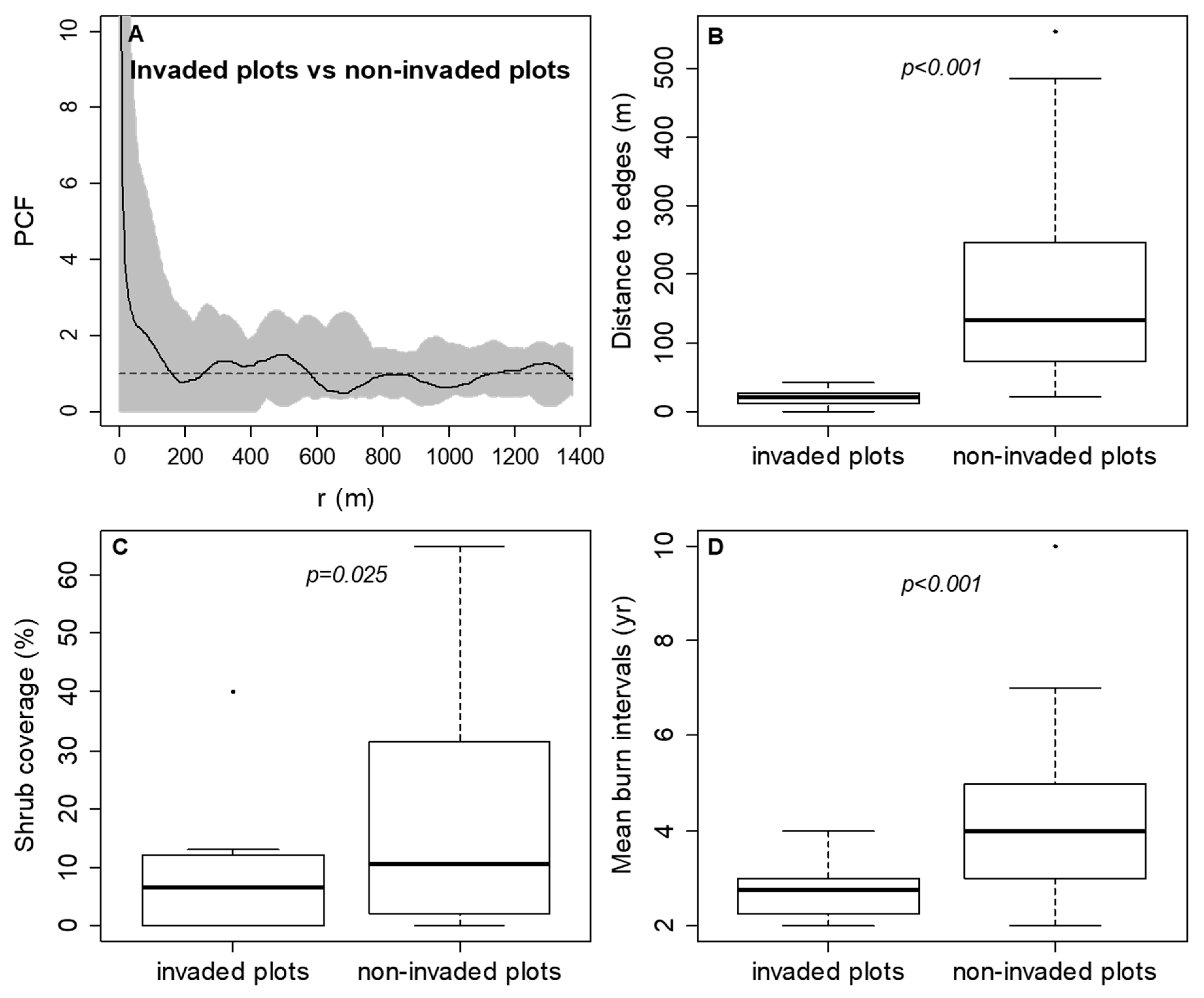
3. Results
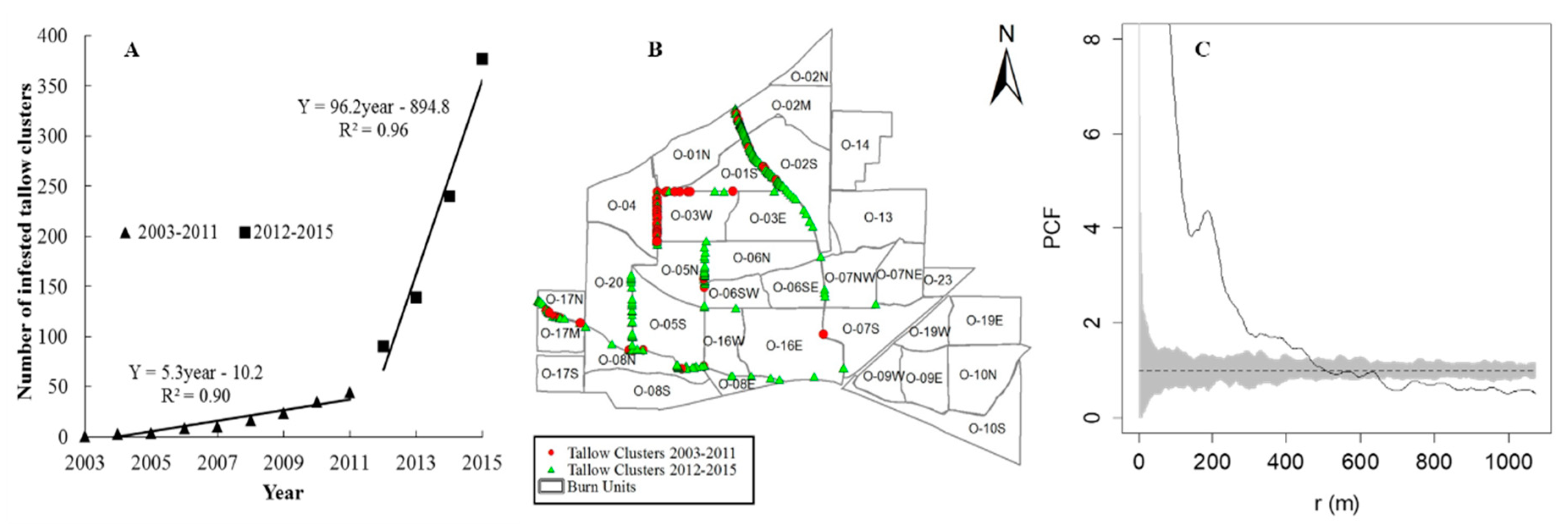
3.1. The Spatiotemporal Pattern and Associated Risk Factors of Tallow Spread along Habitat Edges
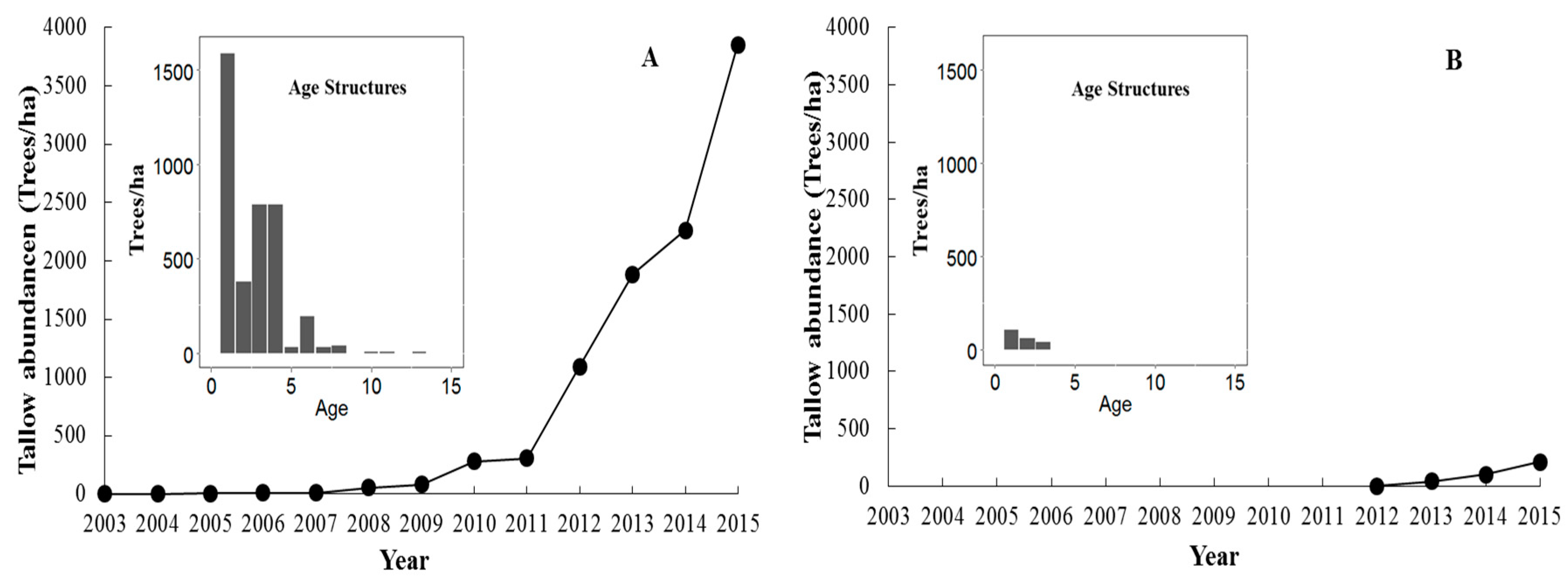
3.2. The Spread of Tallow into Habitat Interiors
4. Discussion
4.1. The Spatiotemproal Pattern and Associated Risk Factors of Tallow Spread along Habitat Edges
4.2. The Spread of Tallow into Habitat Interiors
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lavoie, M.; Starr, G.; Mack, M.; Martin, T.A.; Gholz, H. Effects of a prescribed fire on understory vegetation, carbon pools, and soil nutrients in a longleaf pine-slash pine forest in Florida. Nat. Area J. 2010, 30, 82–94. [Google Scholar] [CrossRef]
- Ford, C.R.; Minor, E.S.; Fox, G.A. Long-term effects of fire and fire-return interval on population structure and growth of longleaf pine (Pinus palustris). Can. J. For. Res. 2010, 40, 1410–1420. [Google Scholar] [CrossRef]
- Imperata brasiliensis, I. cylindrica. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Fort Collins, CO, USA. Available online: http://www.fs.fed.us/database/feis/ (accessed on 25 October 2012).
- Gan, J.; Miller, J.H.; Wang, H.; Taylor, J.W. Invasion of tallow tree into southern US forests: Influencing factors and implications for mitigation. Can. J. For. Res. 2009, 39, 1346–1356. [Google Scholar] [CrossRef]
- Triadica sebifera. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory: Fort Collins, CO, USA. Available online: http://www.fs.fed.us/database/feis/ (accessed on 22 October 2012).
- Fan, Z.; Tan, Y.; Crosby, M.K. Factors associated with the spread of Chinese Tallow in East Texas forestlands. Open J. Ecol. 2012, 02, 121–130. [Google Scholar] [CrossRef]
- Grace, J.B. Can prescribed fire save the endangered coastal prairie ecosystem from Chinese tallow invasion? Endanger. Species Update 1998, 15, 70–76. [Google Scholar]
- Pile, L.S.; Wang, G.G.; Knapp, B.O.; Walker, J.L.; Stambaugh, M.C. Chinese tallow (Triadica sebifera) invasion in maritime forests: The role of anthropogenic disturbance and its management implication. For. Ecol. Manag. 2017, 398, 10–24. [Google Scholar] [CrossRef]
- Jubinsky, G.; Anderson, L.C. The invasive potential of Chinese tallow-tree (Sapium sebiferum Roxb.) in the Southeast. Castanea 1996, 1, 226–231. [Google Scholar]
- Miller, J.H. Nonnative invasive plants of southern forests: A field guide for identification and control. Gen. Tech. Rep. 2006, 62, 103. [Google Scholar]
- Randall, J.M.; Marinelli, J. Invasive Plants: Weeds of the Global Garden, 1st ed.; Brooklyn Botanic Garden: Brooklyn, NY, USA, 1996; p. 111. [Google Scholar]
- Barrilleaux, T.C.; Grace, J.B. Growth and invasive potential of Sapium sebiferum (Euphorbiaceae) within the coastal prairie region: The effects of soil and moisture regime. Am. J. Bot. 2000, 87, 1099–1106. [Google Scholar] [CrossRef]
- Dewalt, S.J.; Siemann, E.; Rogers, W.E. Geographic distribution of genetic variation among native and introduced populations of Chinese tallow tree, Triadica sebifera (Euphorbiaceae). Am. J. Bot. 2011, 98, 1128–1138. [Google Scholar] [CrossRef]
- Paudel, S.; Battaglia, L.L. The role of light, soil and human factors on the probability of occurrence of an invasive and three native plant species in coastal transitions of coastal Mississippi, USA. J. Plant Ecol. 2015, 8, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Renne, I.J.; Spira, T.P.; Bridges, W.C. Effects of habitat, burial, age and passage through birds on germination and establishment of Chinese Tallow Tree in Coastal South Carolina. J. Torrey Bot. Soc. 2001, 128, 109. [Google Scholar] [CrossRef]
- Rogers, W.E.; Siemann, E. Effects of simulated herbivory and resource availability on native and invasive exotic tree seedlings. Basic Appl. Ecol. 2002, 3, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Tian, N.; Fan, Z.; Matney, T.G.; Schultz, E.B. Growth and Stem Profiles of Invasive Triadica sebifera in the Mississippi Coast of the United States. For. Sci. 2017, 63, 569–576. [Google Scholar] [CrossRef]
- Zou, J.; Rogers, W.E.; Siemann, E. Increased competitive ability and herbivory tolerance in the invasive plant Sapium sebiferum. Biol. Invasions 2007, 10, 291–302. [Google Scholar] [CrossRef]
- Flack, S.; Furlow, E. America’s least wanted purple plague, green cancer and 10 other ruthless environmental thugs. Nat. Conserv. 1996, 46, 17–23. [Google Scholar]
- Burns, J.H.; Miller, T.E. Invasion of Chinese Tallow (Sapium sebiferum) in the Lake Jackson area, Northern Florida. Am. Midl. Nat. 2004, 152, 410–417. [Google Scholar] [CrossRef]
- Pattison, R.R.; Mack, R.N. Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: Evaluating CLIMEX predictions with field trials. Glob. Chang. Biol. 2008, 14, 813–826. [Google Scholar] [CrossRef]
- Battaglia, L.L.; Denslow, J.S.; Hargis, T.G. Does woody species establishment alter herbaceous community composition of freshwater floating marshes? J. Coast. Res. 2007, 236, 1580–1587. [Google Scholar] [CrossRef]
- Nolfo-Clements, L.E. Vegetative survey of wetland habitats at Jean Lafitte national historical park and preserve in Southeastern Louisiana. Southeast. Nat. 2006, 5, 499–514. [Google Scholar] [CrossRef]
- Zomlefer, W.B.; Giannasi, D.E.; Bettinger, K.A.; Echols, S.L.; Kruse, L.M. Vascular Plant Survey of Cumberland Island National Seashore, Camden County, Georgia. Castanea 2008, 73, 251–282. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z. Spatial analyses of invasion patterns of Chinese Tallow (Triadica sebifera) in a Wet Slash Pine (Pinus elliottii) flatwood in the coastal plain of Mississippi, USA. For. Sci. 2018, 64, 555–563. [Google Scholar] [CrossRef]
- Iii, E.R.; Rangoonwala, A.; Nelson, G.; Ehrlich, R. Mapping the invasive species, Chinese tallow, with EO1 satellite Hyperion hyperspectral image data and relating tallow occurrences to a classified Landsat Thematic Mapper land cover map. Int. J. Remote Sens. 2005, 26, 1637–1657. [Google Scholar]
- Loewenstein, N.J.; Loewenstein, E.F. Non-native plants in the understory of riparian forests across a land use gradient in the Southeast. Urban Ecosyst. 2005, 8, 79–91. [Google Scholar] [CrossRef]
- Conner, W.H.; Mixon, W.D.; Wood, G.W. Maritime forest habitat dynamics on Bulls Island, Cape Romain National Wildlife Refuge, SC, following Hurricane Hugo. For. Ecol. Manag. 2005, 212, 127–134. [Google Scholar] [CrossRef]
- Keeland, B.D.; Gorham, L.E. Delayed tree mortality in the Atchafalaya Basin of southern Louisiana following Hurricane Andrew. Wetlands 2009, 29, 101–111. [Google Scholar] [CrossRef]
- Vance, M.D.; Fahrig, L.; Flather, C.H. Effect of reproductive rate on minimum habitat requirements of forest-breeding birds. Ecology 2003, 84, 2643–2653. [Google Scholar] [CrossRef]
- Grace, J.B.; Allain, L.K.; Baldwin, H.Q.; Billock, A.G.; Eddleman, W.R.; Given, A.M.; Jeske, C.W.; Moss, R. Effects of Prescribed Fire in the Coastal Prairies of Texas; USGS: Reston, VA, USA, 2005; p. 46. [Google Scholar]
- Samuels, I. Invasion of Chinese Tallow (Sapium sebiferum): A Test of Dispersal and Recruitment Limitation in Multiple Habitats. Ph.D. Thesis, University of Forida, Gainesville, FL, USA, 2004. [Google Scholar]
- Wang, H.-H.; Grant, W.E.; Gan, J.; Rogers, W.E.; Swannack, T.M.; Koralewski, T.E.; Miller, J.H.; Taylor, J.W. Integrating spread dynamics and economics of timber production to manage Chinese tallow invasions in Southern U.S. Forestlands. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Henkel, T.K.; Chambers, J.Q.; Baker, D.A. Delayed tree mortality and Chinese tallow (Triadica sebifera) population explosion in a Louisiana bottomland hardwood forest following Hurricane Katrina. For. Ecol. Manag. 2016, 378, 222–232. [Google Scholar] [CrossRef]
- Theoharides, K.A.; Dukes, J.S. Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytol. 2007, 176, 256–273. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.; Yang, S.; Liu, X. Spatiotemporal Patterns and Mechanisms of Chinese Tallowtree (Triadica sebifera) Spread along Edge Habitat in a Coastal Landscape, Mississippi, USA. Invas. Plant Sci. Manag. 2018, 11, 117–126. [Google Scholar] [CrossRef]
- US Fish and Wildlife Service. Mississippi Sandhill Crane National Wildlife Refuge Comprehensive Conservation Plan; US Department of the Interior Fish and Wildlife Service Southeast Region: Atlanta, GA, USA, 2007; p. 151.
- Cressie, N.A.C. Statistics for Spatial Data, Revised ed.; Wiley: New York, NY, USA, 1993; p. 900. [Google Scholar]
- Baddeley, A.; Turner, R. Spatstat: An R Package for Analyzing Spatial Point Patterns. J. Stat. Softw. 2005, 12, 1–42. [Google Scholar] [CrossRef]
- Lutz, J.A.; Larson, A.J.; Furniss, T.J.; Donato, D.C.; Freund, J.A.; Swanson, M.E.; Bible, K.J.; Chen, J.; Franklin, J.F. Spatially nonrandom tree mortality and ingrowth maintain equilibrium pattern in an old-growth Pseudotsuga–Tsugaforest. Ecology 2014, 95, 2047–2054. [Google Scholar] [CrossRef] [PubMed]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression Models for Count Data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef]
- Chapman, E.L.; Chambers, J.Q.; Ribbeck, K.F.; Baker, D.B.; Tobler, M.A.; Zeng, H.; White, D.A. Hurricane Katrina impacts on forest trees of Louisiana’s Pearl River basin. For. Ecol. Manag. 2008, 256, 883–889. [Google Scholar] [CrossRef]
- Conner, W.H.; Duberstein, J.A.; Day, J.W.; Hutchinson, S. Impacts of Changing Hydrology and Hurricanes on Forest Structure and Growth Along a Flooding/Elevation Gradient in a South Louisiana Forested Wetland from 1986 to 2009. Wetlands 2014, 34, 803–814. [Google Scholar] [CrossRef]
- Harcombe, P.A.; Leipzig, L.E.M.; Elsik, I.S. Effects of Hurricane Rita on three long-term forest study plots in east Texas, USA. Wetlands 2009, 29, 88–100. [Google Scholar] [CrossRef]
- Pearson, D.E.; Ortega, Y.K.; Villarreal, D.; Lekberg, Y.; Cock, M.C.; Eren, Ö.; Hierro, J.L. The fluctuating resource hypothesis explains invasibility, but not exotic advantage following disturbance. Ecology 2018, 99, 1296–1305. [Google Scholar] [CrossRef]
- Aikio, S.; Duncan, R.P.; Hulme, P.E. Lag-phases in alien plant invasions: Separating the facts from the artefacts. Oikos 2010, 119, 370–378. [Google Scholar] [CrossRef]
- Scheld, H.W.; Cowles, J.R. Woody biomass potential of the Chinese tallow tree. Econ. Bot. 1981, 35, 391–397. [Google Scholar] [CrossRef]
- Warren, R.J.; Bahn, V.; Bradford, M.A. The interaction between propagule pressure, habitat suitability and density-dependent reproduction in species invasion. Oikos 2011, 121, 874–881. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Hoopes, M.F.; Marchetti, M.P. Invasion Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2013; p. 45. [Google Scholar]
- Taylor, L.A.V.; Cruzan, M.B. Propagule pressure and disturbance drive the invasion of Perennial False-Brome (Brachypodium sylvaticum). Invas. Plant Sci. Manag. 2015, 8, 169–180. [Google Scholar] [CrossRef]
- Renne, I.J.; Barrow, W.C.; Randall, L.A.J.; Bridges, W.C. Generalized avian dispersal syndrome contributes to Chinese tallow tree (Sapium sebiferum, Euphorbiaceae) invasiveness. Divers. Distrib. 2002, 8, 285–295. [Google Scholar] [CrossRef]
- Wiser, S.K.; Allen, R.B.; Clinton, P.W.; Platt, K.H. Community structure and forest invasion by an exotic herb over 23 years. Ecology 1998, 79, 2071. [Google Scholar] [CrossRef]
- Renne, I.J.; Gauthreaux, S.A.; Gresham, C.A. Seed dispersal of the Chinese Tallow Tree (Sapium sebiferum (L.) Roxb.) by birds in Coastal South Carolina. Am. Midl. Nat. 2000, 144, 202–215. [Google Scholar] [CrossRef]
- Colautti, R.I.; Grigorovich, I.A.; Macisaac, H.J. Propagule pressure: A null model for biological invasions. Biol. Invasions 2007, 9, 885. [Google Scholar] [CrossRef]
- Simberloff, D. The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Eschtruth, A.K.; Battles, J.J. The importance of quantifying propagule pressure to understand invasion: An examination of riparian forest invasibility. Ecology 2011, 92, 1314–1322. [Google Scholar] [CrossRef]
- Brooks, W.R.; Jordan, R.C. Propagule pressure and native species richness effects drive invasibility in tropical dry forest seedling layers. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 162–170. [Google Scholar] [CrossRef]
- Houseman, G.R.; Foster, B.L.; Brassil, C.E. Propagule pressure-invasibility relationships: Testing the influence of soil fertility and disturbance with Lespedeza cuneata. Oecologia 2013, 174, 511–520. [Google Scholar] [CrossRef]
- Hammond, D.H.; Varner, J.M.; Kush, J.S.; Fan, Z. Contrasting sapling bark allocation of five southeastern USA hardwood tree species in a fire prone ecosystem. Ecosphere 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Cameron, G.N.; Glumac, E.G.; Eshelman, B.D. Germination and dormancy in seeds of Sapium sebiferum (Chinese tallow tree). J. Coast. Res. 2000, 16, 391–395. [Google Scholar]
- White, D.W.; Stiles, E.W. Bird dispersal of fruits of species introduced into eastern North America. Can. J. Bot. 1992, 70, 1689–1696. [Google Scholar] [CrossRef]
- Bartuszevige, A.M.; Gorchov, D.L. Avian seed dispersal of an invasive Shrub. Biol. Invasions 2006, 8, 1013–1022. [Google Scholar] [CrossRef]
- Terraube, J.; Archaux, F.; Deconchat, M.; Halder, I.V.; Jactel, H.; Barbaro, L. Forest edges have high conservation value for bird communities in mosaic landscapes. Ecol. Evol. 2016, 6, 5178–5189. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. Error | z Value | Pr (>|z|) | |
|---|---|---|---|---|
| For the early invaded roadway (with seed trees) | ||||
| Count model: | ||||
| Intercept | 7.587 | 0.809 | 9.385 | 0.000 |
| Distance to seed trees | −0.009 | 0.004 | −2.025 | 0.043 |
| Year since last fire | 0.320 | 0.061 | 5.286 | 0.000 |
| Log(theta) | 0.849 | 0.167 | 5.076 | 0.000 |
| Binary model: | ||||
| Intercept | 7.580 | 3.431 | 2.209 | 0.027 |
| Shrub coverage | 0.021 | 0.013 | 1.647 | 0.100 |
| Distance to seed trees | 0.034 | 0.015 | 2.242 | 0.025 |
| Year since last fire | −0.662 | 0.259 | −2.553 | 0.011 |
| Distance to neighborhood | 0.468 | 1.869 | −2.502 | 0.012 |
| For the latterly invaded roadway (without seed trees) | ||||
| Count model: | ||||
| Intercept | 5.196 | 0.923 | 5.632 | 0.000 |
| Overstory basal area | 0.724 | 0.366 | 1.977 | 0.048 |
| Year since last fire | 0.924 | 0.398 | 2.324 | 0.020 |
| Log(theta) | 1.452 | 0.414 | 3.507 | 0.000 |
| Binary model: | ||||
| Intercept | −1.231 | 1.172 | −1.05 | 0.294 |
| Shrub coverage | 0.024 | 0.013 | 1.792 | 0.073 |
| Year since last fire | 1.135 | 0.505 | 2.247 | 0.025 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Fan, Z.; Liu, X.; Ezell, A.W.; Spetich, M.A.; Saucier, S.K.; Gray, S.; Hereford, S.G. Effects of Prescribed Fire, Site Factors, and Seed Sources on the Spread of Invasive Triadica sebifera in a Fire-Managed Coastal Landscape in Southeastern Mississippi, USA. Forests 2019, 10, 175. https://doi.org/10.3390/f10020175
Yang S, Fan Z, Liu X, Ezell AW, Spetich MA, Saucier SK, Gray S, Hereford SG. Effects of Prescribed Fire, Site Factors, and Seed Sources on the Spread of Invasive Triadica sebifera in a Fire-Managed Coastal Landscape in Southeastern Mississippi, USA. Forests. 2019; 10(2):175. https://doi.org/10.3390/f10020175
Chicago/Turabian StyleYang, Shaoyang, Zhaofei Fan, Xia Liu, Andrew W. Ezell, Martin A. Spetich, Scott K. Saucier, Sami Gray, and Scott G. Hereford. 2019. "Effects of Prescribed Fire, Site Factors, and Seed Sources on the Spread of Invasive Triadica sebifera in a Fire-Managed Coastal Landscape in Southeastern Mississippi, USA" Forests 10, no. 2: 175. https://doi.org/10.3390/f10020175