Torreya jackii (Taxaceae): A Special Species that is Genetically Admixed, Morphologically Distinct, and Geographically Sympatric with Parent Species

Abstract
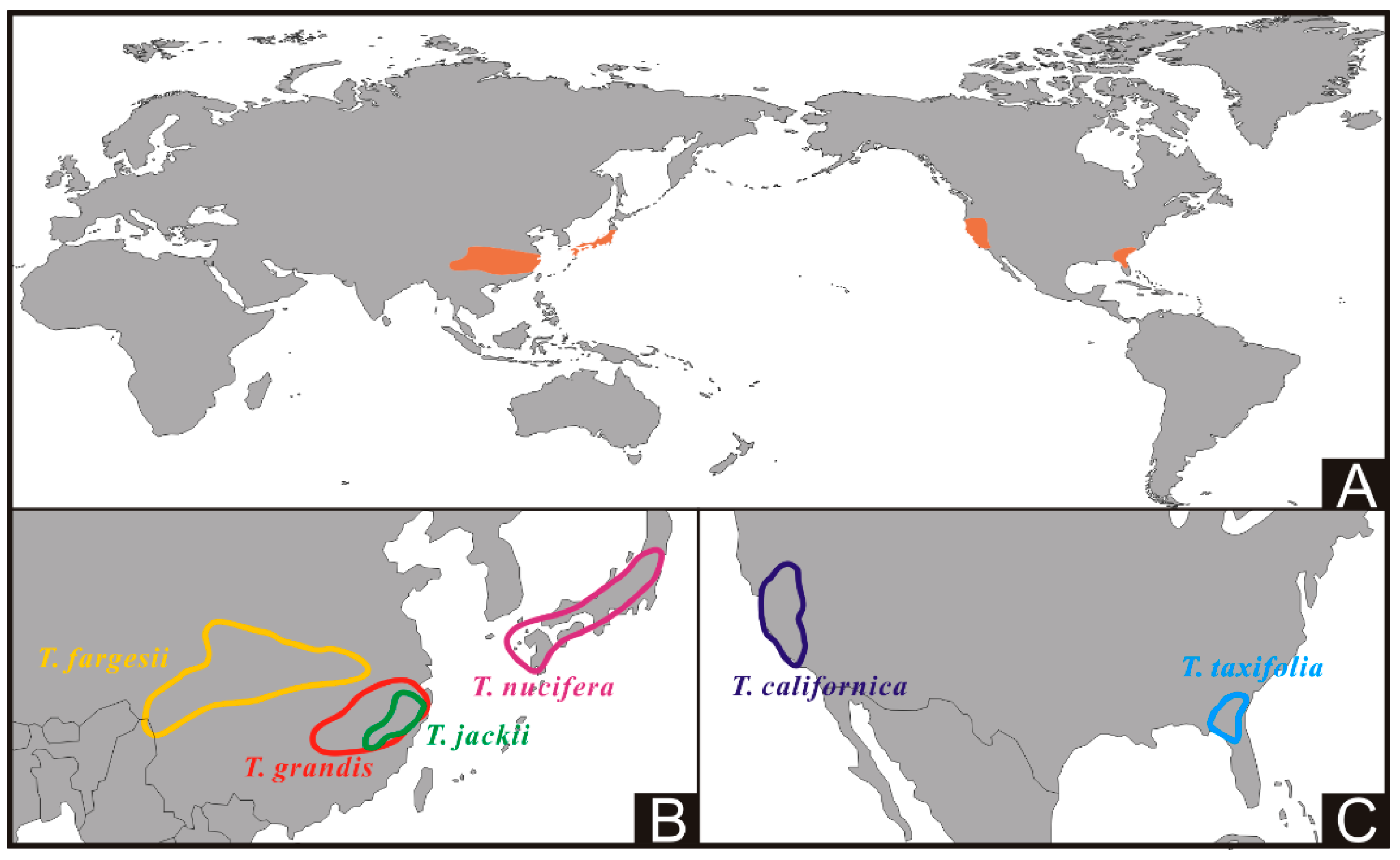
:1. Introduction
2. Materials and Methods
2.1. Sample Collections
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Data Analyses
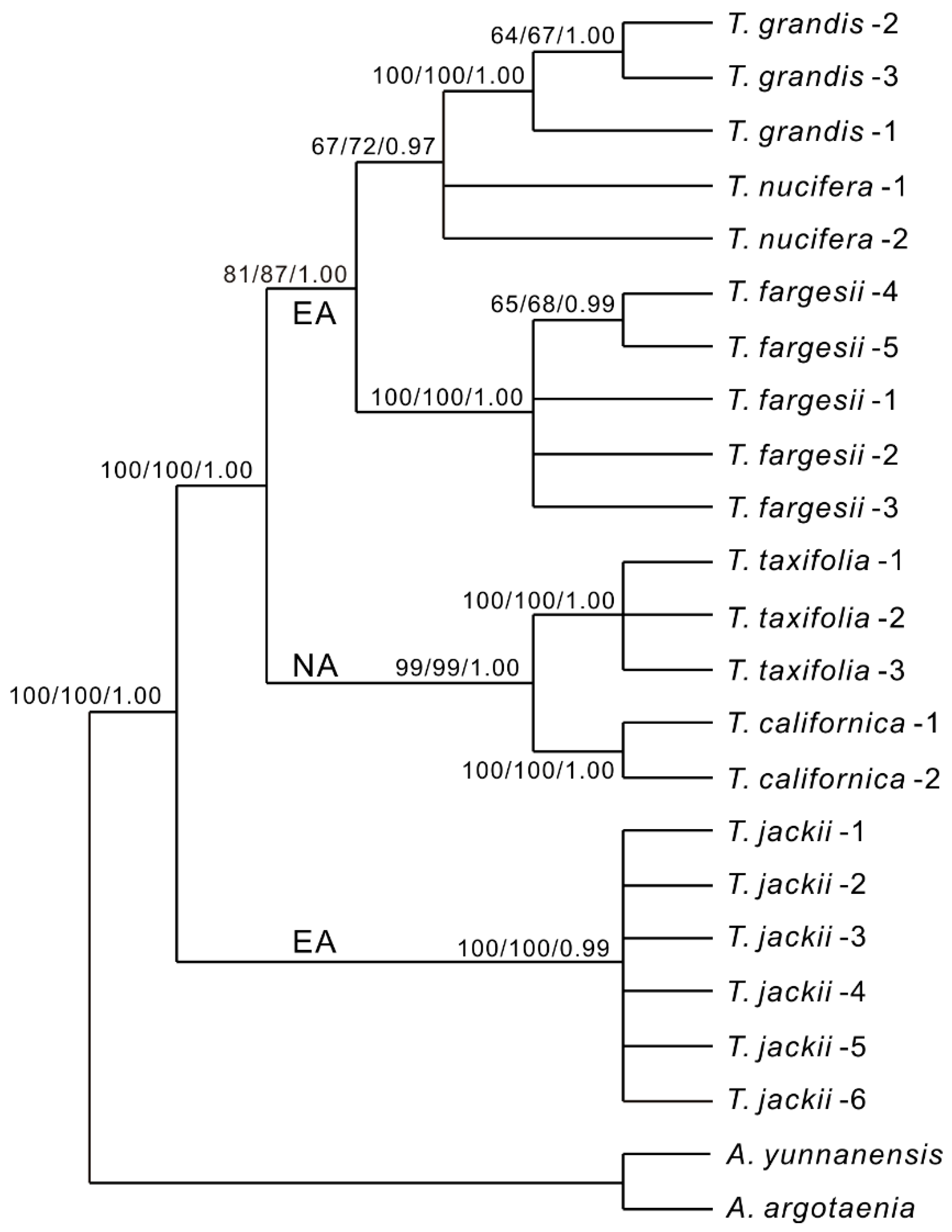
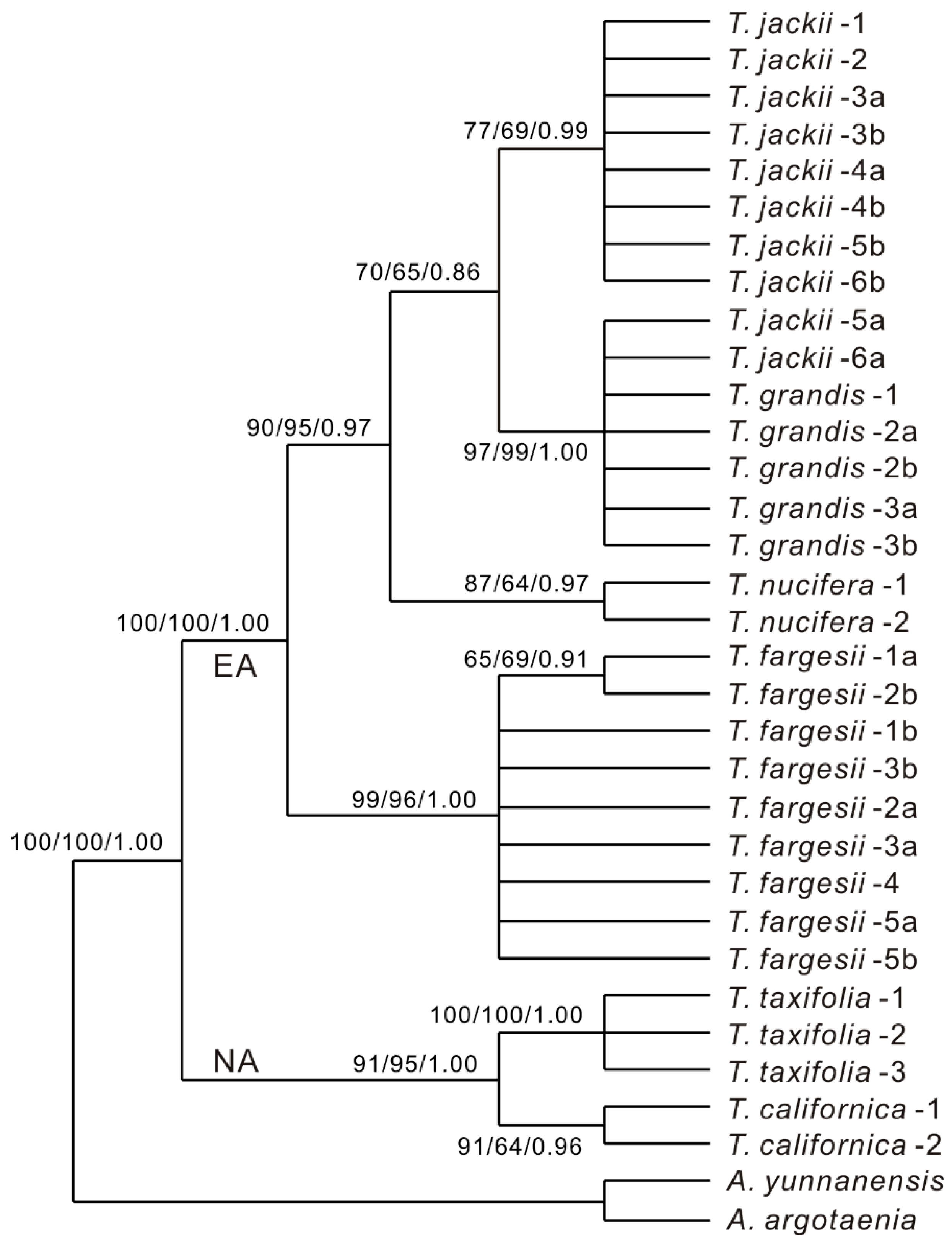
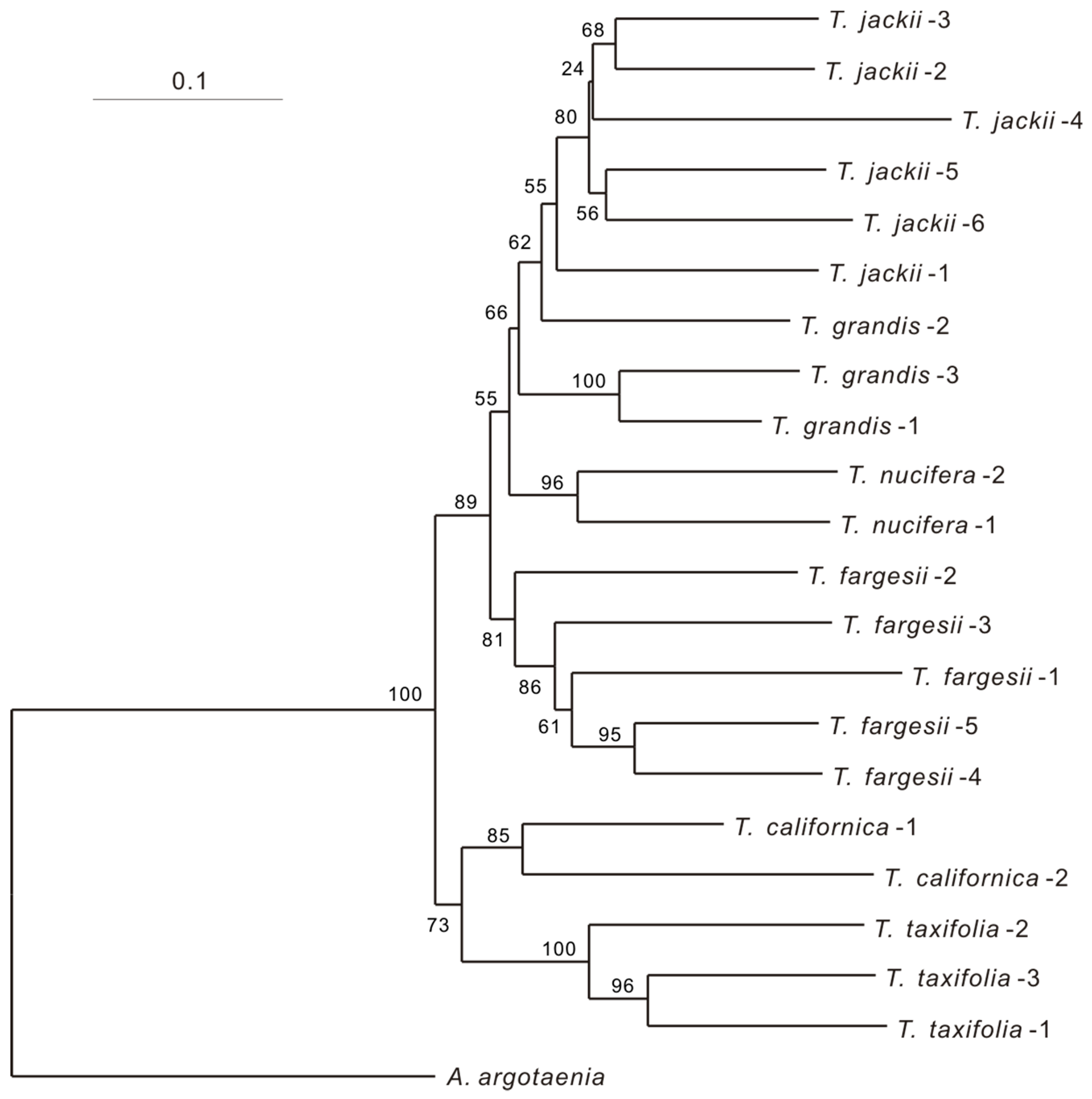
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fu, L.K.; Li, N.; Robert, R.M. Taxaceae. In Flora of China; Hong, D.Y., Raven, P.H., Eds.; Science Press: Beijing, China, 1999; pp. 89–96. (In Chinese) [Google Scholar]
- Farjon, A. A Handbook of the World’s Conifers; Brill Academic Publishers: Leiden, The Netherlands, 2010. [Google Scholar]
- Christenhusz, M.J.M.; Reveal, J.L.; Farjon, A.; Gardner, M.F.; Mill, R.R.; Chase, M.W. New classification and linear sequence of extant gymnosperms. Phytotaxa 2011, 19, 55–70. [Google Scholar] [CrossRef]
- Florin, R. The distribution of conifer and taxad genera in time and space. Acta Horti Bergiani 1963, 20, 122–312. [Google Scholar]
- Hu, H.H. Synoptical study of Chinese Torreya. Contr. Biol. Lab. Sci. Soc. China 1927, 3, 1–9. [Google Scholar]
- Kang, N.; Tang, Z.X. Studies on the taxonomy of the genus Torreya. Bullet. Bot. Res. 1995, 15, 349–362. (In Chinese) [Google Scholar]
- Li, J.H.; Davis, C.C.; Donoghue, M.J.; Kelley, S.; Tredici, P.D. Phylogenetic relationships of Torreya (Taxaceae) inferred from sequences of nuclear ribosomal DNA ITS region. Harv. Pap. Bot. 2001, 6, 275–281. [Google Scholar]
- Hao, D.C.; Xiao, P.G.; Huang, B.L.; Ge, G.B.; Yang, L. Interspecific relationships and origins of Taxaceae and Cephalotaxaceae revealed by partitioned Bayesian analyses of chloroplast and nuclear DNA sequences. Plant Syst. Evol. 2008, 276, 89–104. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Nicolson, R.G.; Tripp, K.; Chaw, S.M. Phylogeny of Taxaceae and Cephalotaxaceae genera inferred from chloroplast matK gene and nuclear rDNA ITS region. Mol. Phylogenet. Evol. 2000, 14, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small amounts of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Grivet, D.; Heinze, B.; Vendramin, G.G.; Petit, R.J. Genome walking with consensus primers: Application to the large single copy region of chloroplast DNA. Mol. Ecol. Notes 2001, 1, 345–349. [Google Scholar] [CrossRef]
- Little, D.P.; Schwarzbach, A.E.; Adams, R.P.; Hsieh, C.F. The circumscription and phylogenetic relationships of Callitropsis and the newly described genus Xanthocyparis (Cupressaceae). Am. J. Bot. 2004, 91, 1872–1881. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, J.; Lickey, E.B.; Beck, J.T.; Farmer, S.B.; Liu, W.; Miller, J.; Siripun, K.S.; Winder, C.T.; Schilling, E.E.; Small, R.L. The tortoise and the hare II: Relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Susanna, A.; Von Raab-Straube, E.; Milne, R.; Liu, J.Q. Island-like radiation of Saussurea (Asteraceae: Cardueae) triggered by uplifts of the Qinghai-Tibetan Plateau. Biol. J. Linn. Soc. 2009, 97, 893–903. [Google Scholar] [CrossRef]
- Kou, Y.X.; Zeng, J.; Liu, J.Q.; Zhao, C.M. Germplasm diversity and differentiation of Helianthus tuberosus L. revealed by AFLP marker and phenotypic traits. J. Agr. Sci. 2014, 152, 779–789. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Li, W.H. Mathematical model of studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Van de Peer, Y.; De Wachter, R. TREECON for Windows: A software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment. Comput. Appl. Biosci. 1994, 10, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Som, A. Causes, consequences and solutions of phylogenetic incongruence. Brief. Bioinform. 2015, 16, 536–548. [Google Scholar] [CrossRef] [PubMed]
- McBreen, K.; Lockhart, P.J. Reconstructing reticulate evolutionary histories of plants. Trends Plant Sci. 2006, 11, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todesco, M.; Pascual, M.A.; Owens, G.L.; Ostevik, K.L.; Moyers, B.T.; Hübner, S.; Heredia, S.M.; Hahn, K.L.; Caseys, C.; Bock, D.G.; et al. Hybridization and extinction. Evol. Appl. 2016, 9, 892–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.W. Study on the biological character and conservation of endemic species Torreya jackii. Chin. Biodivers. 1997, 5, 206–209. (In Chinese) [Google Scholar]
- Wang, X.M.; Zhou, B.Z.; Cao, Y.H.; Wang, K.; Lin, G.X. A GIS-based study on the habitat characteristics of Chinese Torreya in Kuaiji mountains. Acta Agric. Uni. Jiangxiensis 2010, 32, 523–527. (In Chinese) [Google Scholar]
- Dong, L.M.; Shen, D.F.; Yu, W.W.; Zeng, Y.R.; Wu, Z.M.; Dai, W.S. Trait variations of male Torreya grandis trees. J. Zhejiang A F Uni. 2012, 29, 715–721. (In Chinese) [Google Scholar]
- Guan, Q.; Yuan, M.; Yu, Z. Early discrimination of karyotype and sexuality for Chinese Torreya. Sci. Silvae Sinicae 1993, 29, 389–392. (In Chinese) [Google Scholar]
- Buchholz, J.T. The embryogeny of Torreya, with a note on Austrotaxus. Bullet. Torrey Bot. Club 1940, 67, 731–754. [Google Scholar] [CrossRef]
- Vallejo-Marín, M.; Cooley, A.M.; Lee, M.Y.; Folmer, M.; McKain, M.R.; Puzey, J.R. Strongly asymmetric hybridization barriers shape the origin of a new polyploid species and its hybrid ancestor. Am. J. Bot. 2016, 103, 1–17. [Google Scholar] [CrossRef]
- Mallet, J. Hybridization as an invasion of the genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Currat, M.; Ruedi, M.; Petit, R.J.; Excoffier, L. The hidden side of invasions: Massive introgression by local genes. Evolution 2008, 62, 1908–1920. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.C.; Fu, L.K. Torreya Arn. In Flora Reipublicae Popularis Sinica; Cheng, W.C., Fu, L.K., Eds.; Science Press: Beijing, China, 1978; pp. 457–467. (In Chinese) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Geographic Origin/Voucher No. | GenBank No. (matK, rbcL, trnL-trnF, trnD-trnT, rpl16, psbB1-psbB2, trnS-trnG, ITS) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Torreya | |||||||||
| T. jackii -1 | Xianju, Zhejiang/Liu and Noshiro 005 | KJ588955 | KJ589001 | KJ589070 | KJ589047 | KJ589024 | KJ588978 | KJ589093 | KJ588928 |
| T. jackii -2 | Shaowu, Fujian/Kou 0104 | KJ588956 | KJ589002 | KJ589071 | KJ589048 | KJ589025 | KJ588979 | KJ589094 | KJ588929 |
| T. jackii -3 | Jinyun, Zhejiang/Kou 1202 | KJ588957 | KJ589003 | KJ589072 | KJ589049 | KJ589026 | KJ588980 | KJ589095 | KJ588930-31 |
| T. jackii -4 | Xianju, Zhejiang/Kou 1506 | KJ588958 | KJ589004 | KJ589073 | KJ589050 | KJ589027 | KJ588981 | KJ589096 | KJ588932-32 |
| T. jackii -5 | Lishui, Zhejiang/Kou 1101 | KJ588959 | KJ589005 | KJ589074 | KJ589051 | KJ589028 | KJ588982 | KJ589097 | KJ588934-35 |
| T. jackii -6 | Lishui, Zhejiang/Kou 1102 | KJ588960 | KJ589006 | KJ589075 | KJ589052 | KJ589029 | KJ588983 | KJ589098 | KJ588936-37 |
| T. grandis -1 | Lin’an, Zhejiang/Liu and Noshiro 010 | KJ588940 | KJ588986 | KJ589055 | KJ589032 | KJ589009 | KJ588963 | KJ589078 | KJ588907 |
| T. grandis -2 | Zhuji, Zhejiang/Liu and Noshiro 013 | KJ588941 | KJ588987 | KJ589056 | KJ589033 | KJ589010 | KJ588964 | KJ589079 | KJ588908-09 |
| T. grandis -3 | Jinyun, Zhejiang/Kou 1208 | KJ588942 | KJ588988 | KJ589057 | KJ589034 | KJ589011 | KJ588965 | KJ589080 | KJ588910-11 |
| T. fargesii -1 | Jinfoshan, Chongqing/Liu and Noshiro 014 | KJ588947 | KJ588993 | KJ589062 | KJ589039 | KJ589016 | KJ588970 | KJ589085 | KJ588917-18 |
| T. fargesii -2 | Jinfoshan, Chongqing/Liu and Noshiro 016 | KJ588948 | KJ588994 | KJ589063 | KJ589040 | KJ589017 | KJ588971 | KJ589086 | KJ588919-20 |
| T. fargesii -3 | Shenlongjia, Hubei/Liu 200807-2 | KJ588949 | KJ588995 | KJ589064 | KJ589041 | KJ589018 | KJ588972 | KJ589087 | KJ588921-22 |
| T. fargesii -4 | Lijiang, Yunnan/Liu and Noshiro 018 | KJ588945 | KJ588991 | KJ589060 | KJ589037 | KJ589014 | KJ588968 | KJ589083 | KJ588914 |
| T. fargesii -5 | Weixi, Yunnan/Liu and Noshiro 019 | KJ588946 | KJ588992 | KJ589061 | KJ589038 | KJ589015 | KJ588969 | KJ589084 | KJ588915-16 |
| T. nucifera -1 | Forestry and Forest Products Research Institute, Japan/Nakais 200203 | KJ588943 | KJ588989 | KJ589058 | KJ589035 | KJ589012 | KJ588966 | KJ589081 | KJ588912 |
| T. nucifera -2 | Kunming Bot. Gard. (from Japan)/8 | KJ588944 | KJ588990 | KJ589059 | KJ589036 | KJ589013 | KJ588967 | KJ589082 | KJ588913 |
| T. taxifolia -1 | Kunming Bot. Gard./1062-89-c (Arnold Arborefum) | KJ588950 | KJ588996 | KJ589065 | KJ589042 | KJ589019 | KJ588973 | KJ589088 | KJ588923 |
| T. taxifolia -2 | Kunming Bot. Gard./857-87*a | KJ588951 | KJ588997 | KJ589066 | KJ589043 | KJ589020 | KJ588974 | KJ589089 | KJ588924 |
| T. taxifolia -3 | Kunming Bot. Gard./1054-89-I | KJ588952 | KJ588998 | KJ589067 | KJ589044 | KJ589021 | KJ588975 | KJ589090 | KJ588925 |
| T. californica -1 | Philipps-Universität Marburg Bot. Gard./7 | KJ588953 | KJ588999 | KJ589068 | KJ589045 | KJ589022 | KJ588976 | KJ589091 | KJ588926 |
| T. californica -2 | San Francisco, California/Bruce Bartholomew, s.n. SFBG# XY-226 | KJ588954 | KJ589000 | KJ589069 | KJ589046 | KJ589023 | KJ588977 | KJ589092 | KJ588927 |
| Amentotaxus | |||||||||
| A. argotaenia | Jinfoshan, Chongqing/3025 | KJ588962 | KJ589008 | KJ589077 | KJ589054 | KJ589031 | KJ588985 | KJ589100 | KJ588939 |
| A. yunnanensis | Heilongtan, Kunming/3007 | KJ588961 | KJ589007 | KJ589076 | KJ589053 | KJ589030 | KJ588984 | KJ589099 | KJ588938 |
| Loci | Length | Variable Sites | PI | CI | RI |
|---|---|---|---|---|---|
| ITS | 1281 | 52 | 46 | 0.839 | 0.960 |
| matK | 1434 | 14 | 13 | 1.000 | 1.000 |
| psbB1-psbB2 | 1434 | 4 | 3 | 1.000 | 1.000 |
| rbcL | 1299 | 12 | 11 | 0.944 | 0.969 |
| rpl16 | 962 | 13 | 11 | 1.000 | 1.000 |
| trnD-trnT | 1519 | 21 | 14 | 0.980 | 0.986 |
| trnL-trnF | 914 | 15 | 13 | 0.984 | 0.987 |
| trnS-trnG | 679 | 8 | 4 | 0.905 | 0.918 |
| Plastid/ITS | T. fargesii | T. nucifera | T. taxifolia | T. californica | T. grandis | T. jackii |
|---|---|---|---|---|---|---|
| T. fargesii | 0.20 | 0.42 | 0.40 | 0.27 | 0.33 | |
| T. nucifera | 0.92 | 0.28 | 0.26 | 0.12 | 0.21 | |
| T. taxifolia | 2.29 | 2.10 | 0.39 | 0.37 | 0.39 | |
| T. californica | 1.90 | 1.72 | 0.68 | 0.35 | 0.36 | |
| T. grandis | 1.35 | 0.74 | 2.54 | 2.16 | 0.33 | |
| T. jackii | 1.20 | 0.60 | 2.39 | 2.01 | 0.57 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-J.; Xiao, K.; Kou, Y.-X. Torreya jackii (Taxaceae): A Special Species that is Genetically Admixed, Morphologically Distinct, and Geographically Sympatric with Parent Species. Forests 2019, 10, 174. https://doi.org/10.3390/f10020174
Wang Y-J, Xiao K, Kou Y-X. Torreya jackii (Taxaceae): A Special Species that is Genetically Admixed, Morphologically Distinct, and Geographically Sympatric with Parent Species. Forests. 2019; 10(2):174. https://doi.org/10.3390/f10020174
Chicago/Turabian StyleWang, Yu-Jin, Kun Xiao, and Yi-Xuan Kou. 2019. "Torreya jackii (Taxaceae): A Special Species that is Genetically Admixed, Morphologically Distinct, and Geographically Sympatric with Parent Species" Forests 10, no. 2: 174. https://doi.org/10.3390/f10020174