
Effect of Obstructive Sleep Apnea and CPAP Treatment on the Bioavailability of Erythrocyte and Plasma Nitric Oxide
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioethics Approval
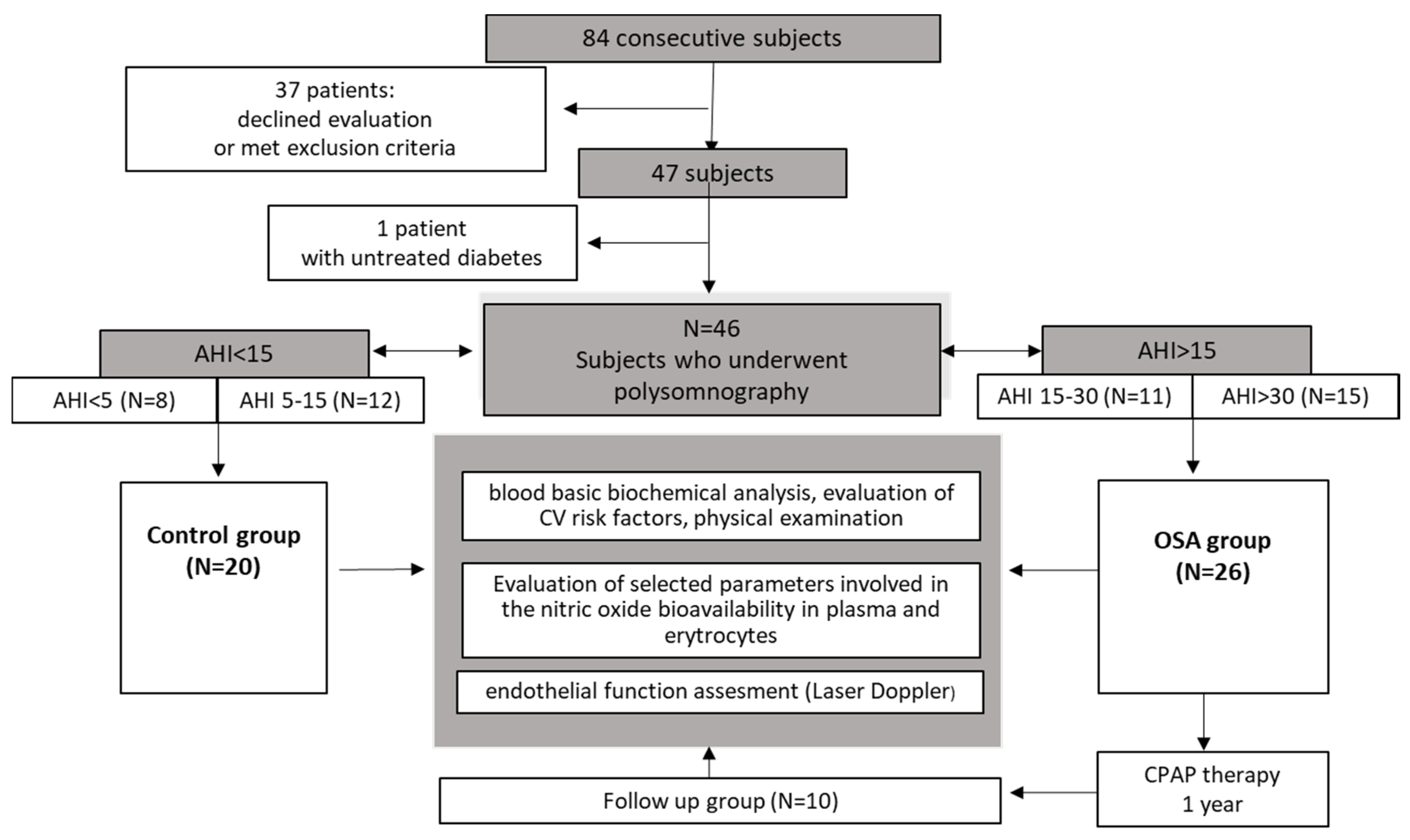
2.2. Subject Recruitment
2.3. Measurements
- -
- apneas—defined as the absence of airflow for ≥10 s,
- -
- hypopneas (a reduction in the amplitude of breathing by ≥30% for ≥10 s with a ≥3% decline in blood oxygen saturation SpO2) or followed by arousal.
2.4. Subgroups
2.5. Biochemical Analysis
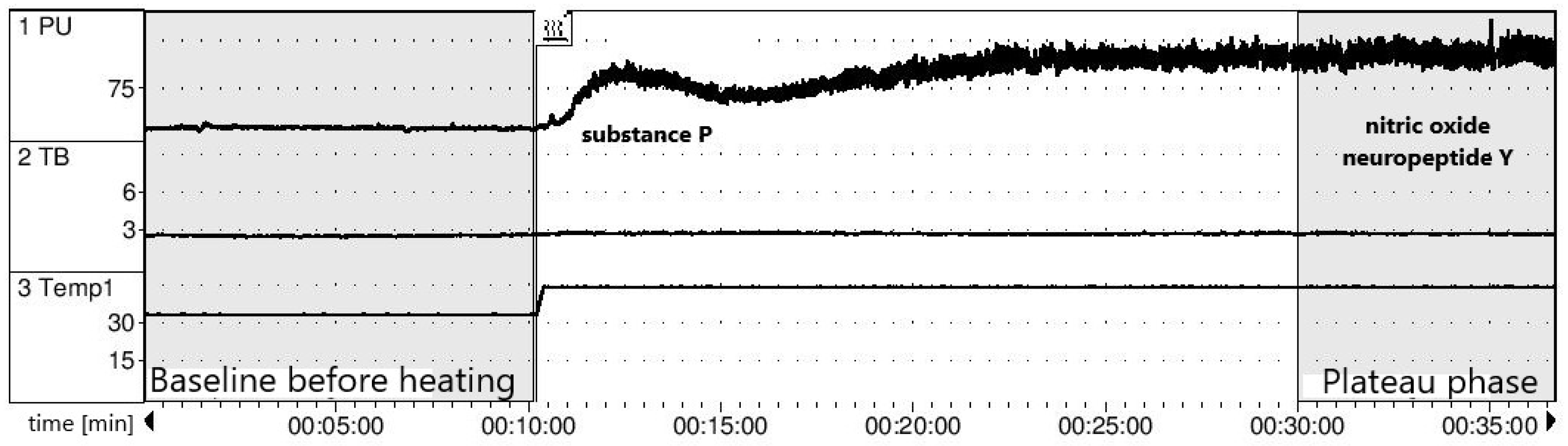
2.6. Endothelial Function Assessment
2.7. Statistical Analysis
3. Results
3.1. Baseline Characteristics
3.2. Assessment of Baseline Endothelial Function
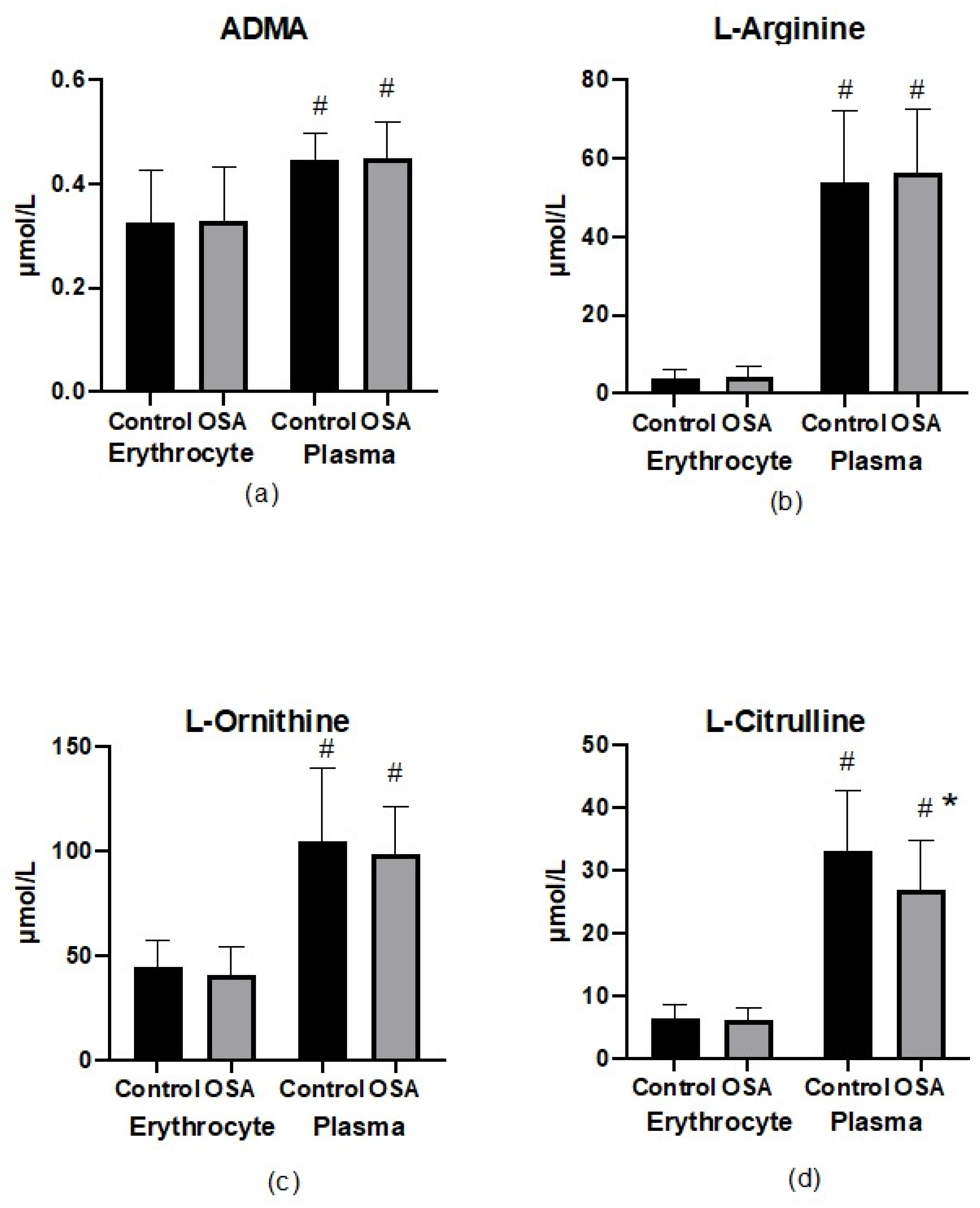
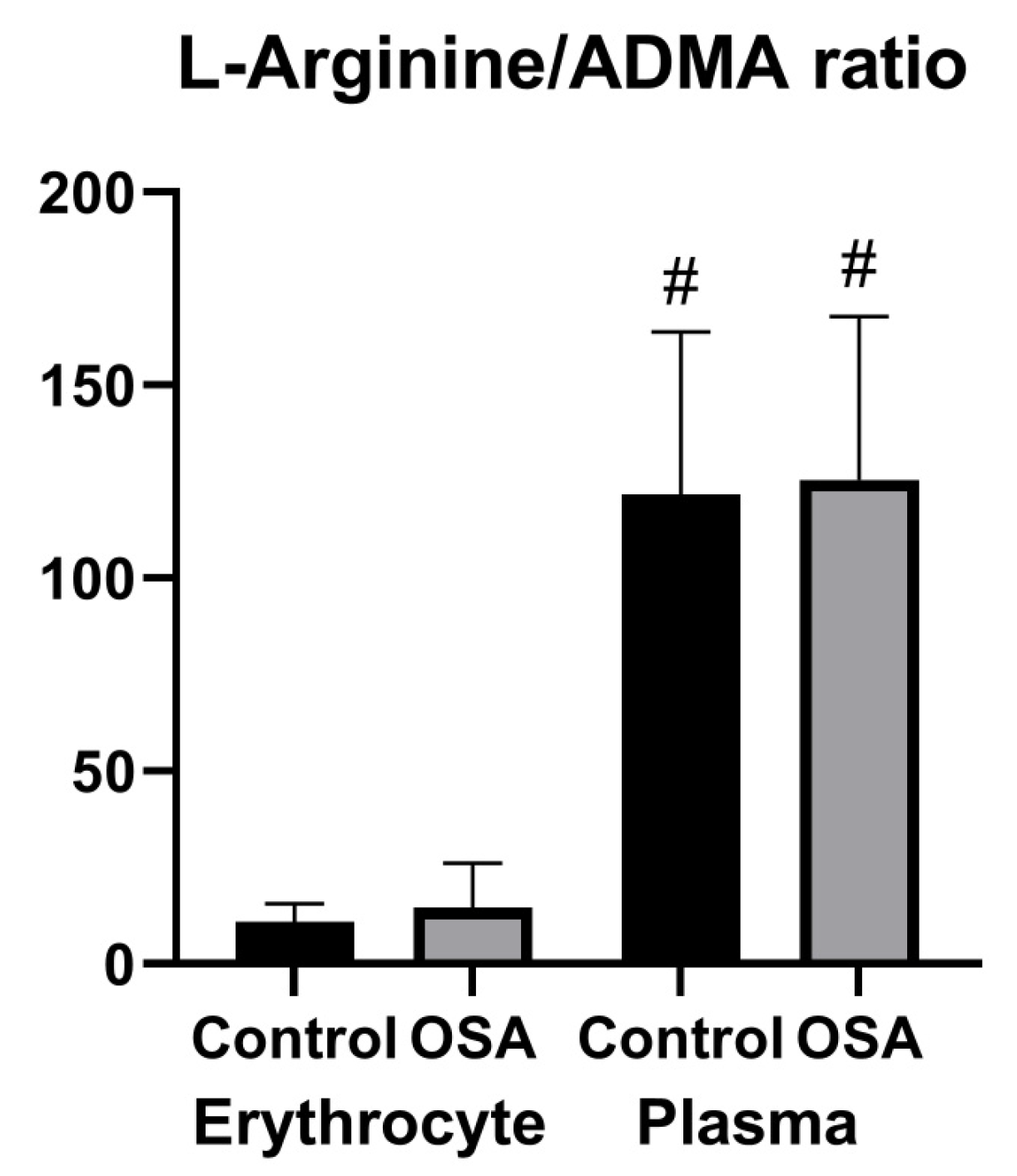
3.3. Parameters of Nitric Oxide Bioavailability in Erythrocytes and Plasma
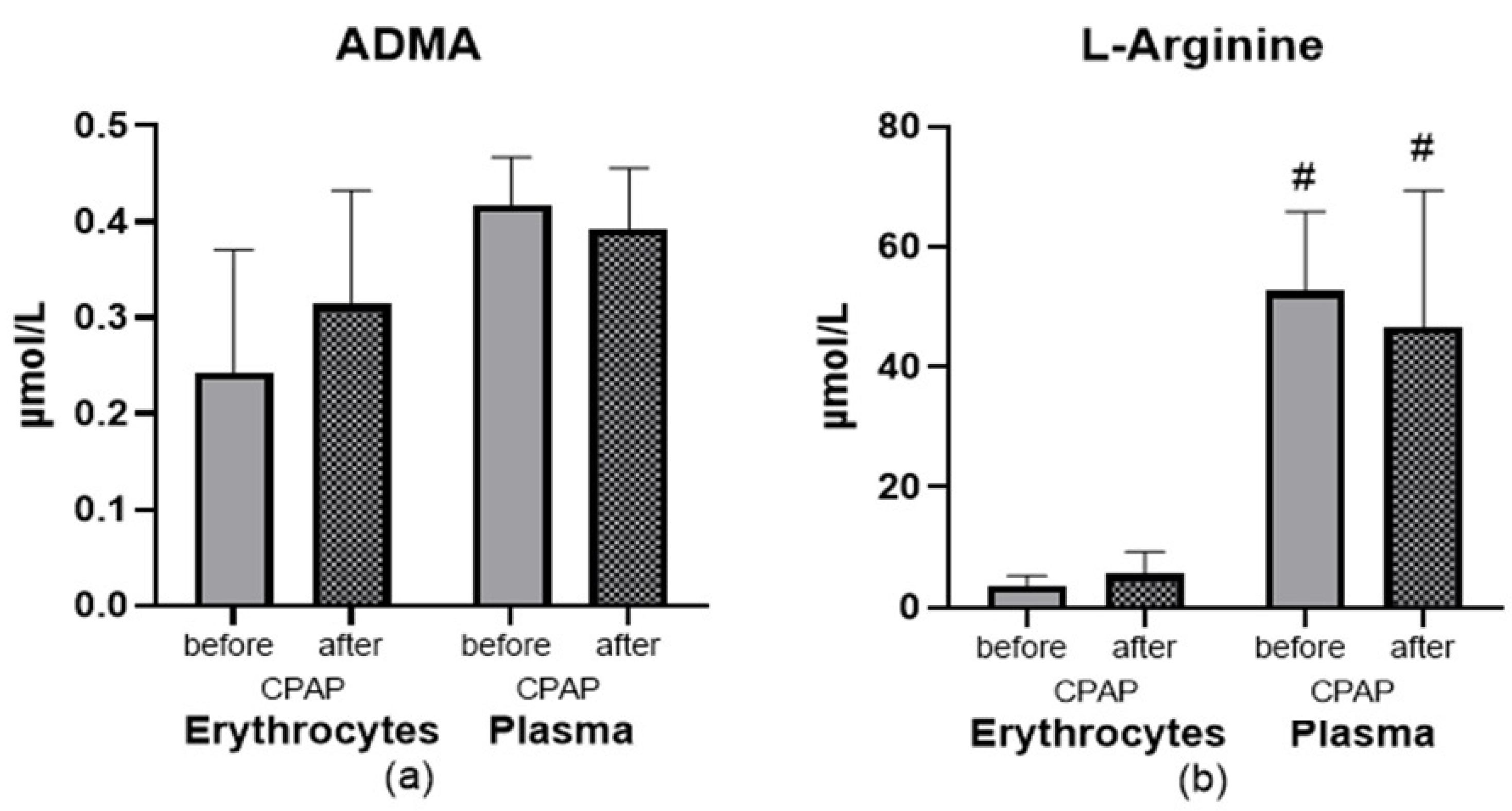
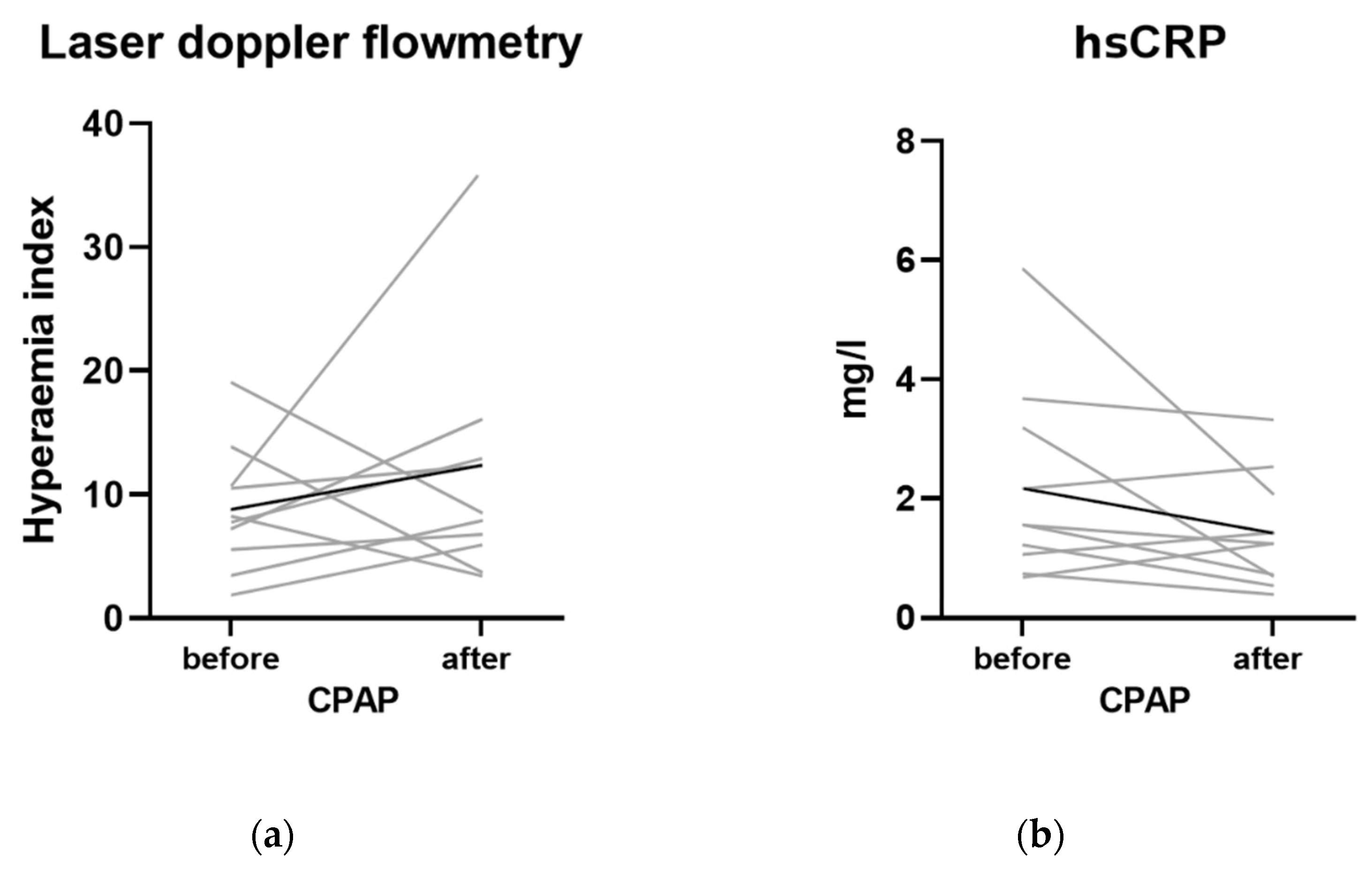
3.4. Effect of CPAP
4. Discussion
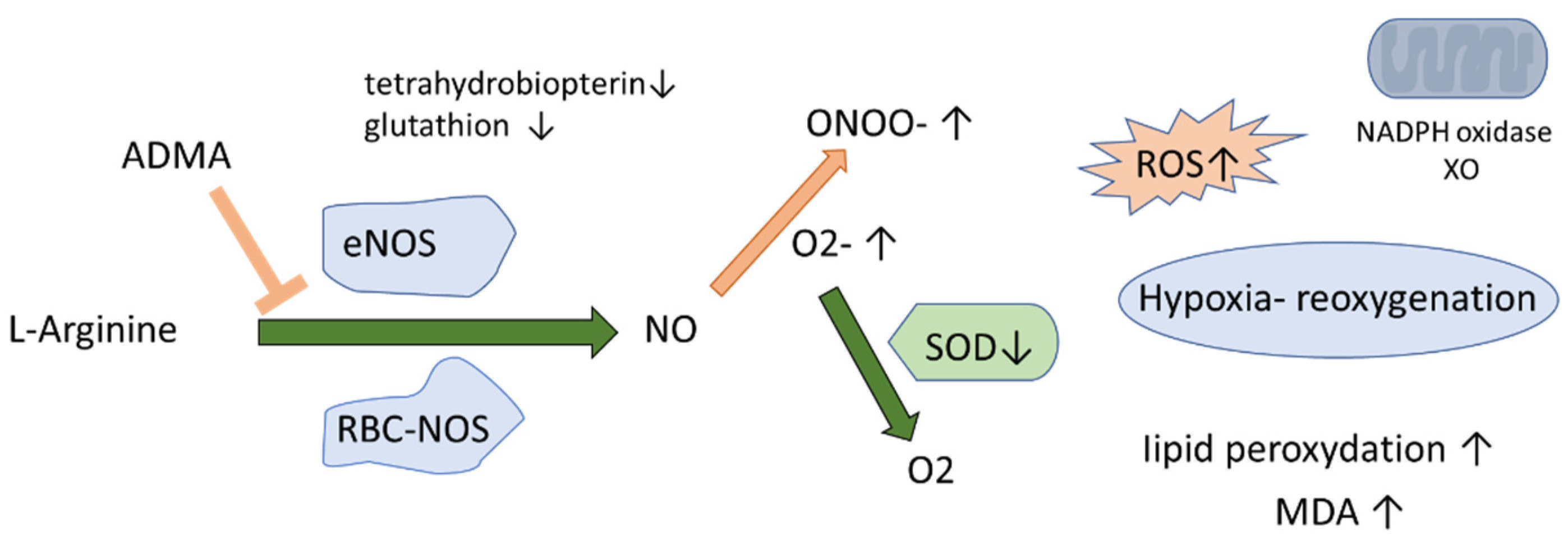
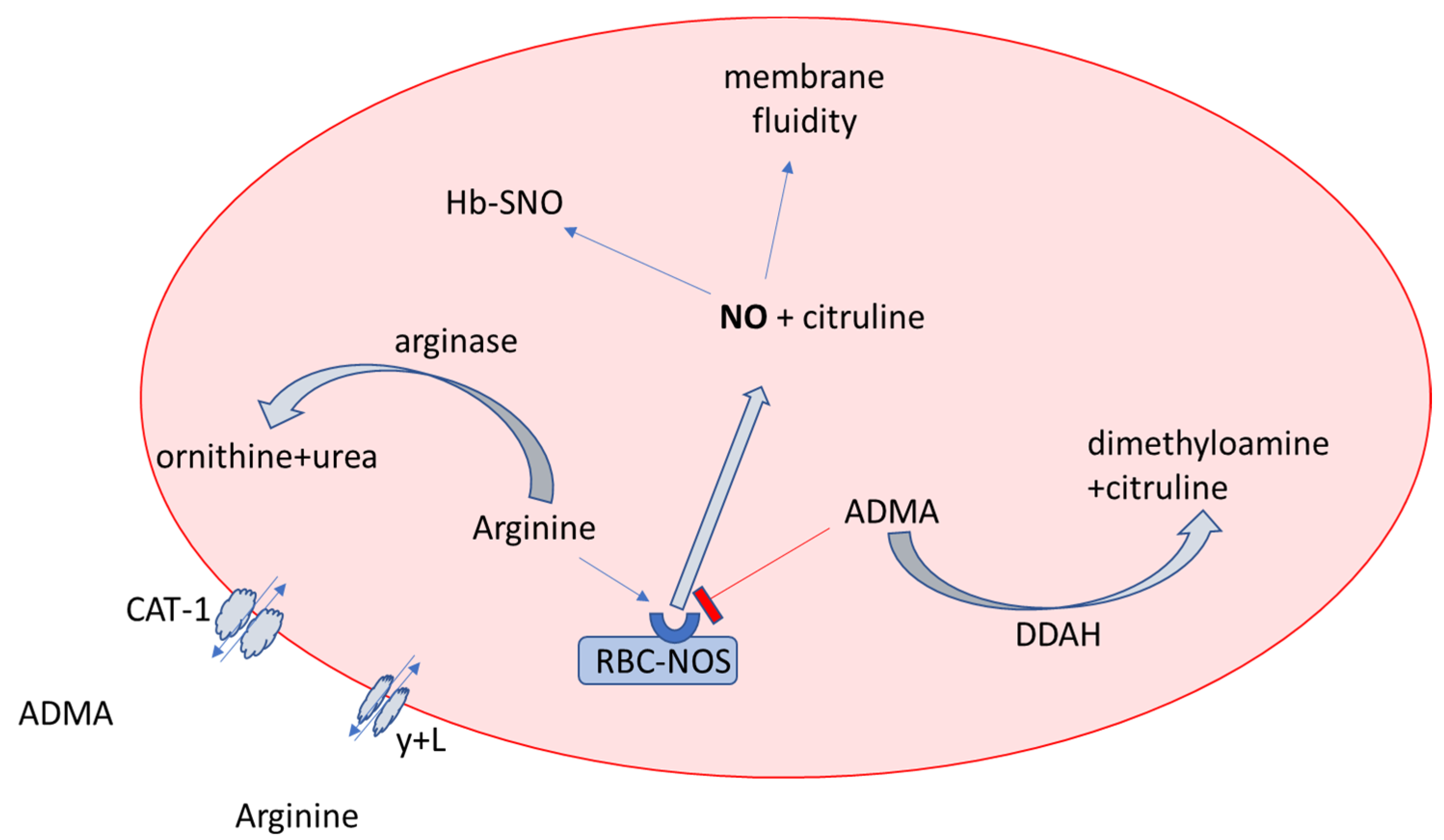
4.1. NO Biotranformation in Erythrocytes and Plasma in OSA and Non-OSA Subjects
4.2. Effect of CPAP
4.3. L-Citrulline and NO Production
4.4. Other Differences in the Demographic and Biochemical Characteristics between the Groups
5. Study Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benjafield, A.V.; Ayas, N.T.; Eastwood, P.R.; Heinzer, R.; Ip, M.S.M.; Morrell, M.J.; Nunez, C.M.; Patel, S.R.; Penzel, T.; Pépin, J.L.D.; et al. Estimation of the global prevalence and burden of obstructive sleep apnoea: A literature-based analysis. Lancet Respir. Med. 2019, 7, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Stanek, A.; Fazeli, B.; Bartuv, S.; Sutkowska, E. Editorial The Role of Endothelium in Physiological and Pathological States: New Data. BioMed Res. Int. 2018, 2018, 1098039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanek, A.; Brożyna-Tkaczyk, K.; Myśliński, W. Oxidative Stress Markers among Obstructive Sleep Apnea Patients. Oxidative Med. Cell. Longev. 2021, 2021, 9681595. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Gomez, J.-J.; Singh, I.; Khan, M. S-Nitrosylation in Regulation of Inflammation and Cell Damage. Curr. Drug Targets 2018, 19, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Pau, M.C.; Mangoni, A.A.; Zinellu, E.; Pintus, G.; Carru, C.; Fois, A.G.; Pirina, P.; Zinellu, A. Circulating Superoxide Dismutase Concentrations in Obstructive Sleep Apnoea (OSA): A Systematic Review and Meta-Analysis. Antioxidants 2021, 10, 1764. [Google Scholar] [CrossRef]
- Pau, M.C.; Zinellu, E.; Fois, S.S.; Piras, B.; Pintus, G.; Carru, C.; Mangoni, A.A.; Fois, A.G.; Zinellu, A.; Pirina, P. Circulating Malondialdehyde Concentrations in Obstructive Sleep Apnea (OSA): A Systematic Review and Meta-Analysis with Meta-Regression. Antioxidants 2021, 10, 1053. [Google Scholar] [CrossRef]
- Gao, F.; Lucke-Wold, B.P.; Li, X.; Logsdon, A.F.; Xu, L.C.; Xu, S.; LaPenna, K.B.; Wang, H.; Talukder, M.A.H.; Siedlecki, C.A.; et al. Reduction of Endothelial Nitric Oxide Increases the Adhesiveness of Constitutive Endothelial Membrane ICAM-1 through Src-Mediated Phosphorylation. Front. Physiol. 2018, 8, 1124. [Google Scholar] [CrossRef] [Green Version]
- Böger, R.H.; Maas, R.; Schulze, F.; Schwedhelm, E. Asymmetric dimethylarginine (ADMA) as a prospective marker of cardiovascular disease and mortality—An update on patient populations with a wide range of cardiovascular risk. Pharmacol. Res. 2009, 60, 481–487. [Google Scholar] [CrossRef]
- Billecke, S.S.; Kitzmiller, L.A.; Northrup, J.J.; Whitesall, S.E.; Kimoto, M.; Hinz, A.V.; D’Alecy, L.G. Contribution of whole blood to the control of plasma asymmetrical dimethylarginine. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1788–H1796. [Google Scholar] [CrossRef]
- Davids, M.; van Hell, A.J.; Visser, M.; Nijveldt, R.J.; van Leeuwen, P.A.M.; Teerlink, T. Role of the human erythrocyte in generation and storage of asymmetric dimethylarginine. Am. J. Physiol. Circ. Physiol. 2012, 302, H1762–H1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoro, M.; Suzuki, M.; Murota, K.; Otsuka, C.; Yamashita, H.; Takahashi, Y.; Tsuji, H.; Kimoto, M. Asymmetric dimethylarginine, an endogenous NOS inhibitor, is actively metabolized in rat erythrocytes. Biosci. Biotechnol. Biochem. 2012, 76, 1334–1342. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, A.O.; Bulau, P.; Zakrzewicz, D.; Kitowska, K.E.; Weissmann, N.; Grimminger, F.; Morty, R.E.; Eickelberg, O. Increased protein arginine methylation in chronic hypoxia: Role of protein arginine methyltransferases. Am. J. Respir. Cell Mol. Biol. 2006, 35, 436–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwemer, C.F.; Davenport, R.D.; Gomez-Espina, J.; Blanco-Gonzalez, E.; Whitesall, S.E.; D’Alecy, L.G. Packed red blood cells are an abundant and proximate potential source of Nitric oxide synthase inhibition. PLoS ONE 2015, 10, e0119991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsikas, D.; Böhmer, A.; Großkopf, H.; Beckmann, B.; Dreißigacker, U.; Jordan, J.; Maassen, N. Clinical-chemistry laboratory relevant hemolysis is unlikely to compromise human plasma concentration of free asymmetric dimethylarginine (ADMA). Clin. Biochem. 2012, 45, 1536–1538. [Google Scholar] [CrossRef]
- Hannemann, J.; Zummack, J.; Hillig, J.; Böger, R. Metabolism of asymmetric dimethylarginine in hypoxia: From bench to bedside. Pulm. Circ. 2020, 10, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Kang, E.S.; Cates, T.B.; Harper, D.N.; Chiang, T.M.; Myers, L.K.; Acchiardo, S.R.; Kimoto, M. An enzyme hydrolyzing methylated inhibitors of nitric oxide synthase is present in circulating human red blood cells. Free Radic. Res. 2001, 35, 693–707. [Google Scholar] [CrossRef]
- Bollenbach, A.; Tsikas, D. Pharmacological activation of dimethylarginine dimethylaminohydrolase (DDAH) activity by inorganic nitrate and DDAH inhibition by NG-hydroxy-l-arginine, Nω,Nω-dimethyl-l-citrulline and Nω,Nω-dimethyl-Nδ-hydroxy-l-citrulline: Results and overview. Amino Acids 2019, 51, 483–494. [Google Scholar] [CrossRef]
- Banjarnahor, S.; Rodionov, R.N.; König, J.; Maas, R. Transport of l-arginine related cardiovascular risk markers. J. Clin. Med. 2020, 9, 3975. [Google Scholar] [CrossRef]
- Shima, Y.; Maeda, T.; Aizawa, S.; Tsuboi, I.; Kobayashi, D.; Kato, R.; Tamai, I. l-arginine import via cationic amino acid transporter CAT1 is essential for both differentiation and proliferation of erythrocytes. Blood 2006, 107, 1352–1356. [Google Scholar] [CrossRef]
- Badran, M.; Golbidi, S.; Ayas, N.; Laher, I. Nitric Oxide Bioavailability in Obstructive Sleep Apnea: Interplay of Asymmetric Dimethylarginine and Free Radicals. Sleep Disord. 2015, 2015, 387801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, J.; Mieth, M.; Endreß, B.; Auge, D.; König, J.; Fromm, M.F.; Maas, R. Interaction of the cardiovascular risk marker asymmetric dimethylarginine (ADMA) with the human cationic amino acid transporter 1 (CAT1). J. Mol. Cell. Cardiol. 2012, 53, 392–400. [Google Scholar] [CrossRef]
- Peled, N.; Kassirer, M.; Kramer, M.R.; Rogowski, O.; Shlomi, D.; Fox, B.; Berliner, A.S.; Shitrit, D. Increased erythrocyte adhesiveness and aggregation in obstructive sleep apnea syndrome. Thromb. Res. 2008, 121, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.; Pauly, S.; Ali, J.; Walpurgis, K.; Thevis, M.; Bloch, W.; Suhr, F. RBC-NOS-Dependent S-Nitrosylation of Cytoskeletal Proteins Improves RBC Deformability. PLoS ONE 2013, 8, e56759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, K.; Nishio, I. An association between plasma asymmetric dimethylarginine and membrane fluidity of erythrocytes in hypertensive and normotensive men: An electron paramagnetic resonance investigation. Am. J. Hypertens. 2005, 18, 1243–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, R.B.; Brooks, R.; Gamaldo, C.; Harding, S.M.; Lloyd, R.M.; Quan, S.F.; Troester, M.T.; Vaughn, B.V. AASM Scoring Manual Updates for 2017 (Version 2.4). J. Clin. Sleep Med. 2017, 13, 665. [Google Scholar] [CrossRef]
- Qiu, Z.H.; Luo, Y.M.; McEvoy, R.D. The Sleep Apnea Cardiovascular Endpoints (SAVE) study: Implications for health services and sleep research in China and elsewhere. J. Thorac. Dis. 2017, 9, 2217–2220. [Google Scholar] [CrossRef] [Green Version]
- Fleszar, M.G.; Wiśniewski, J.; Krzystek-Korpacka, M.; Misiak, B.; Frydecka, D.; Piechowicz, J.; Lorenc-Kukuła, K.; Gamian, A. Quantitative Analysis of l-Arginine, Dimethylated Arginine Derivatives, l-Citrulline, and Dimethylamine in Human Serum Using Liquid Chromatography–Mass Spectrometric Method. Chromatographia 2018, 81, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Minson, C.T. Mechanisms and Modulators of Temperature Regulation: Thermal provocation to evaluate microvascular reactivity in human skin. J. Appl. Physiol. 2010, 109, 1239. [Google Scholar] [CrossRef] [Green Version]
- Bironneau, V.; Tamisier, R.; Trzepizur, W.; Andriantsitohaina, R.; Berger, M.; Goupil, F.; Joyeux-Faure, M.; Jullian-Desayes, I.; Launois, S.; Le Vaillant, M.; et al. Sleep apnoea and endothelial dysfunction: An individual patient data meta-analysis. Sleep Med. Rev. 2020, 52, 101309. [Google Scholar] [CrossRef]
- Arlouskaya, Y.; Sawicka, A.; Głowala, M.; Giebułtowicz, J.; Korytowska, N.; Tałałaj, M.; Nowicka, G.; Wrzosek, M. Asymmetric Dimethylarginine (ADMA) and Symmetric Dimethylarginine (SDMA) Concentrations in Patients with Obesity and the Risk of Obstructive Sleep Apnea (OSA). J. Clin. Med. 2019, 8, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barceló, A.; De La Peña, M.; Ayllón, O.; Vega-Agapito, M.V.; Piérola, J.; Pérez, G.; González, C.; Alonso, A.; Agustí, A.G.N. Increased plasma levels of asymmetric dimethylarginine and soluble CD40 ligand in patients with sleep apnea. Respiration. 2009, 77, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, Y.; Fırat, H.; Şimşek, B.; Torun, M.; Yardim-Akaydin, S. Circulating nitric oxide (NO), asymmetric dimethylarginine (ADMA), homocysteine, and oxidative status in obstructive sleep apnea-hypopnea syndrome (OSAHS). Sleep Breath. 2008, 12, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Teerlink, T.; Luo, Z.; Palm, F.; Wilcox, C.S. CELLULAR ADMA: REGULATION AND ACTION. Pharmacol. Res. 2009, 60, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Q.; Gao, J.; Lu, R.; Jin, Y.; Zou, Y.; Yu, C.; Yan, Y. Asymmetric dimethylarginine compartmental behavior during high-flux hemodialysis. Ren. Fail. 2020, 42, 760–766. [Google Scholar] [CrossRef]
- Yang, J.; Gonon, A.T.; Sjöquist, P.O.; Lundberg, J.O.; Pernow, J. Arginase regulates red blood cell nitric oxide synthase and export of cardioprotective nitric oxide bioactivity. Proc. Natl. Acad. Sci. USA 2013, 110, 15049–15054. [Google Scholar] [CrossRef] [Green Version]
- Mahdi, A.; Tengbom, J.; Alvarsson, M.; Wernly, B.; Zhou, Z.; Pernow, J. Red Blood Cell Peroxynitrite Causes Endothelial Dysfunction in Type 2 Diabetes Mellitus via Arginase. Cells 2020, 9, 1712. [Google Scholar] [CrossRef]
- Premont, R.T.; Reynolds, J.D.; Zhang, R.; Stamler, J.S. Role of Nitric Oxide Carried by Hemoglobin in Cardiovascular Physiology: Developments on a Three-Gas Respiratory Cycle. Circ. Res. 2020, 126, 129–158. [Google Scholar] [CrossRef]
- Msaad, S.; Chaabouni, A.; Marrakchi, R.; Boudaya, M.; Kotti, A.; Feki, W.; Jamoussi, K.; Kammoun, S. Nocturnal Continuous Positive Airway Pressure (nCPAP) Decreases High-Sensitivity C-Reactive Protein (hs-CRP) in Obstructive Sleep Apnea-Hypopnea Syndrome. Sleep Disord. 2020, 2020, 8913247. [Google Scholar] [CrossRef]
- Papadia, C.; Osowska, S.; Cynober, L.; Forbes, A. Citrulline in health and disease. Review on human studies. Clin. Nutr. 2018, 37, 1823–1828. [Google Scholar] [CrossRef]
- Ambrosetti, M.; Lucioni, A.M.; Conti, S.; Pedretti, R.F.; Neri, M. Metabolic syndrome in obstructive sleep apnea and related cardiovascular risk. J. Cardiovasc. Med. 2006, 7, 826–829. [Google Scholar] [CrossRef] [PubMed]
- Stanek, A.; Brożyna-Tkaczyk, K.; Myśliński, W. The role of obesity-induced perivascular adipose tissue (Pvat) dysfunction in vascular homeostasis. Nutrients 2021, 13, 3843. [Google Scholar] [CrossRef] [PubMed]
- Stanek, A.; Wielkoszyński, T.; Bartuś, S.; Cholewka, A. Whole-body cryostimulation improves inflammatory endothelium parameters and decreases oxidative stress in healthy subjects. Antioxidants 2020, 9, 1308. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control Group AHI < 15 n = 20 | OSA Group AHI > 15 n = 26 | p-Value |
|---|---|---|---|
| (Mean ± SD) or Median(Q1–Q3) | (Mean ± SD) or Median (Q1–Q3) | ||
| Men (n, %) | 13 65% | 24 92% | NS |
| Women (n, %) | 7 35% | 2 8% | NS |
| Age (years) | 50.6 ± 7.41 | 50.6 ± 9.95 | NS |
| Height (cm) | 173.6 ± 11.7 | 174.9 ± 6.93 | NS |
| Weight (kg) | 79.2 ± 17.4 | 92.0 ± 16.2 | <0.05 |
| Hypertension (n, %) | 4.0 20% | 11.0 42% | NS |
| BMI (kg/m2) median(Q1–Q3) | 25.4 (23.7–28.2) | 29.1 (26.4–33.1) | <0.05 |
| RBC (mln/µL) | 5.04 ± 0.36 | 5.05 ± 0.34 | NS |
| WBC (k/µL) median(Q1–Q3) | 5.66 (4.41–6.91) | 6.66 (5.22–8.98) | <0.05 |
| Hb (g/dL) | 15.2 ± 1.13 | 15.3 ± 1.00 | NS |
| PLT (k/µL) | 233.7 ± 45.0 | 243.0 ± 62.6 | NS |
| Ht (%) | 44.8 ± 3.14 | 45.5 ± 2.61 | NS |
| MCV (fL) | 88.9 ± 2.53 | 90.2 ± 4.01 | NS |
| MCH (pg) | 30.2 ± 1.11 | 30.3 ± 1.33 | NS |
| MCHC (g/dL) | 34.0 ± 0.83 | 33.6 ± 0.91 | NS |
| HbA1c (%) | 5.52 ± 0.36 | 5.58 ± 0.38 | NS |
| ALT (U/L) median(Q1–Q3) | 25.0 (19.5–41.5) | 33.5 (22–39) | NS |
| LDL (mg/dL) | 129.3 ± 35.7 | 140.0 ± 27.6 | NS |
| Total cholesterol (mg/dL) | 212.1 ± 36.9 | 224.6 ± 39.9 | NS |
| HDL (mg/dL) | 57.9 ± 11.3 | 51.6 ± 12.7 | NS |
| Triglycerides (mg/dL) median(Q1–Q3) | 124.6 (86–145) | 164.9 (116–208) | NS |
| TSH (µlU/mL) median(Q1–Q3) | 1.46 (1.04–1.64) | 1.32 (0.9–1.8) | NS |
| Creatinine (mg/dL) median(Q1–Q3) | 0.97 (0.86–1.04) | 0.99 (0.89–1.08) | NS |
| eGFR (mL/min/1.73 m2) | 80.5 ± 11.8 | 86.5 ± 14.8 | NS |
| Uric acid (mg/dL) | 4.95 ± 1.05 | 6.38 ± 0.92 | <0.05 |
| Urea (mg/dL) | 30.3 ± 6.76 | 31.5 ± 7.53 | NS |
| Mg (mmol/L) | 2.13 ± 0.11 | 2.12 ± 0.14 | NS |
| K (mmol/L) | 4.35 ± 0.29 | 4.28 ± 0.28 | NS |
| Na (mmol/L) | 140.4 ± 1.83 | 141.5 ± 1.86 | NS |
| hsCRP (mg/L) median(Q1–Q3) | 0.46 (0.28–0.73) | 1.43 (0.80–3.45) | <0.05 |
| Ca (mmol/L) | 9.37 ± 0.35 | 9.34 ± 0.27 | NS |
| Glucose (mg/dL) | 94.9 ± 10.4 | 100.5 ± 9.80 | NS |
| Insulin (µU/mL) median(Q1–Q3) | 6.15 (5.0–7.3) | 11.9 (6.4–14.4) | <0.05 |
| HOMA-IR median(Q1–Q3) | 1.48 (1.16–1.64) | 2.93 (1.36–3.78) | <0.05 |
| QUICKI median | 0.36 ± 0.03 | 0.33 ± 0.03 | <0.05 |
| AHI (events/h) median(Q1–Q3) | 6.1 (3.0–8.3) | 36.4 (20–37) | <0.05 |
| ODI (events/h) median(Q1–Q3) | 5.0 (2.2–9.0) | 34.4 (20–39) | <0.05 |
| Mean saturation (%) | 94.3 ± 1.2 | 92.5 ± 1.7 | <0.05 |
| Duration of desaturation < 90% (% of total sleep time) median(Q1–Q3) | 0.1 (0.0–0.9) | 5.1 (1.6–11.0) | <0.05 |
| Hyperemia Index | |
|---|---|
| Mean ± SD | |
| Control group | 11.2 ± 5.9 |
| OSA group before CPAP | 9.6 ± 5.3 |
| OSA after 1 year of CPAP | 11.6 ± 4.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mochol, J.; Gawryś, J.; Szahidewicz-Krupska, E.; Wiśniewski, J.; Fortuna, P.; Rola, P.; Martynowicz, H.; Doroszko, A. Effect of Obstructive Sleep Apnea and CPAP Treatment on the Bioavailability of Erythrocyte and Plasma Nitric Oxide. Int. J. Environ. Res. Public Health 2022, 19, 14719. https://doi.org/10.3390/ijerph192214719
Mochol J, Gawryś J, Szahidewicz-Krupska E, Wiśniewski J, Fortuna P, Rola P, Martynowicz H, Doroszko A. Effect of Obstructive Sleep Apnea and CPAP Treatment on the Bioavailability of Erythrocyte and Plasma Nitric Oxide. International Journal of Environmental Research and Public Health. 2022; 19(22):14719. https://doi.org/10.3390/ijerph192214719
Chicago/Turabian StyleMochol, Jakub, Jakub Gawryś, Ewa Szahidewicz-Krupska, Jerzy Wiśniewski, Paulina Fortuna, Piotr Rola, Helena Martynowicz, and Adrian Doroszko. 2022. "Effect of Obstructive Sleep Apnea and CPAP Treatment on the Bioavailability of Erythrocyte and Plasma Nitric Oxide" International Journal of Environmental Research and Public Health 19, no. 22: 14719. https://doi.org/10.3390/ijerph192214719