
Profiling Analysis of Filter Feeder Polypedilum (Chironomidae) Gut Contents Using eDNA Metabarcoding Following Contrasting Habitat Types—Weir and Stream
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
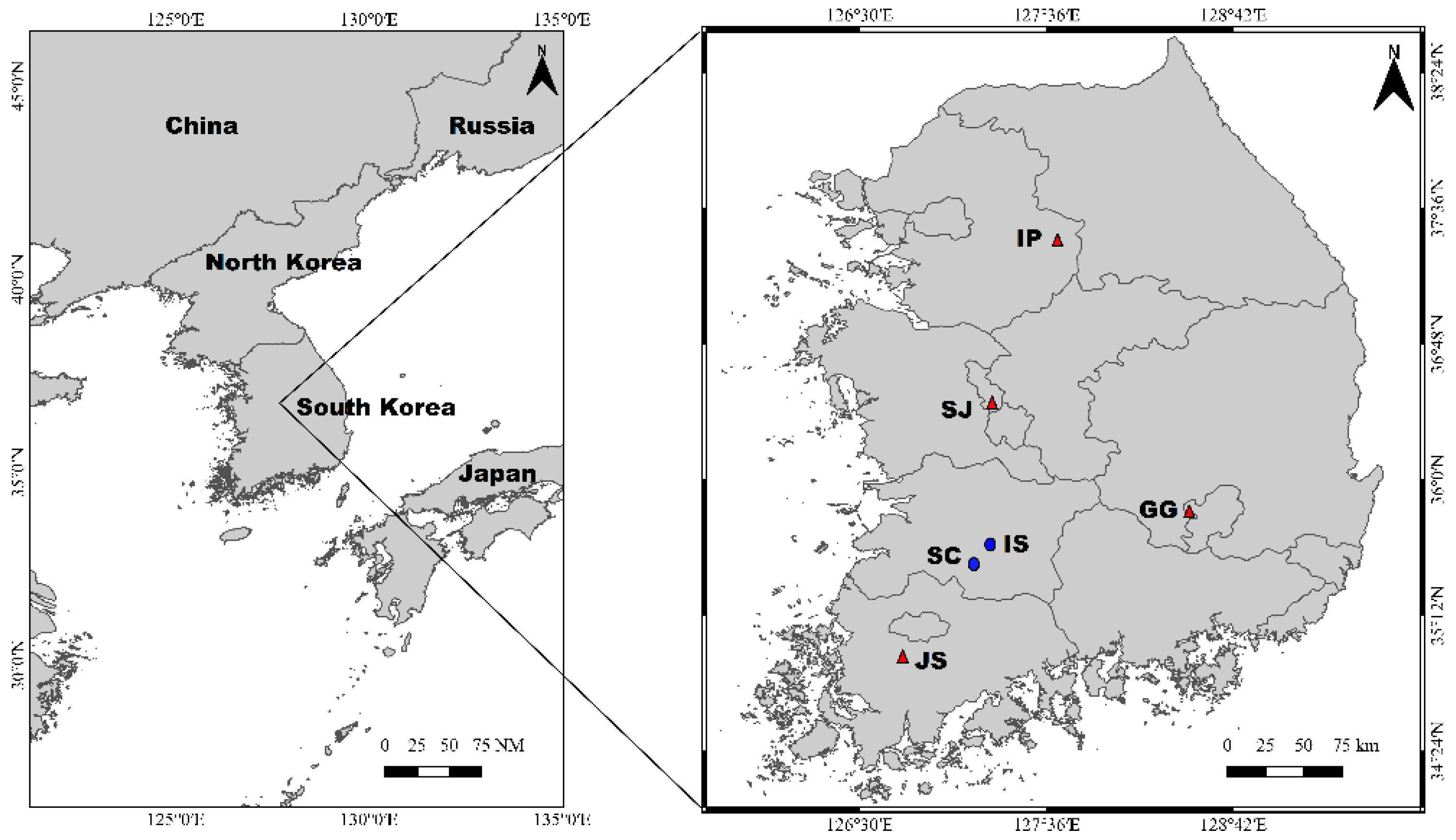
2.1. Field Sampling and Measuring Factors
2.2. DNA Extraction and Metagenomic Sequencing
2.3. Bioinformatics Analysis of the Sequence Reads
2.4. Statistical Analysis of OTUs and Environmental Factors
3. Results
3.1. Diversity of Polypedilum Larvae Gut Content Using V9 Metabarcoding
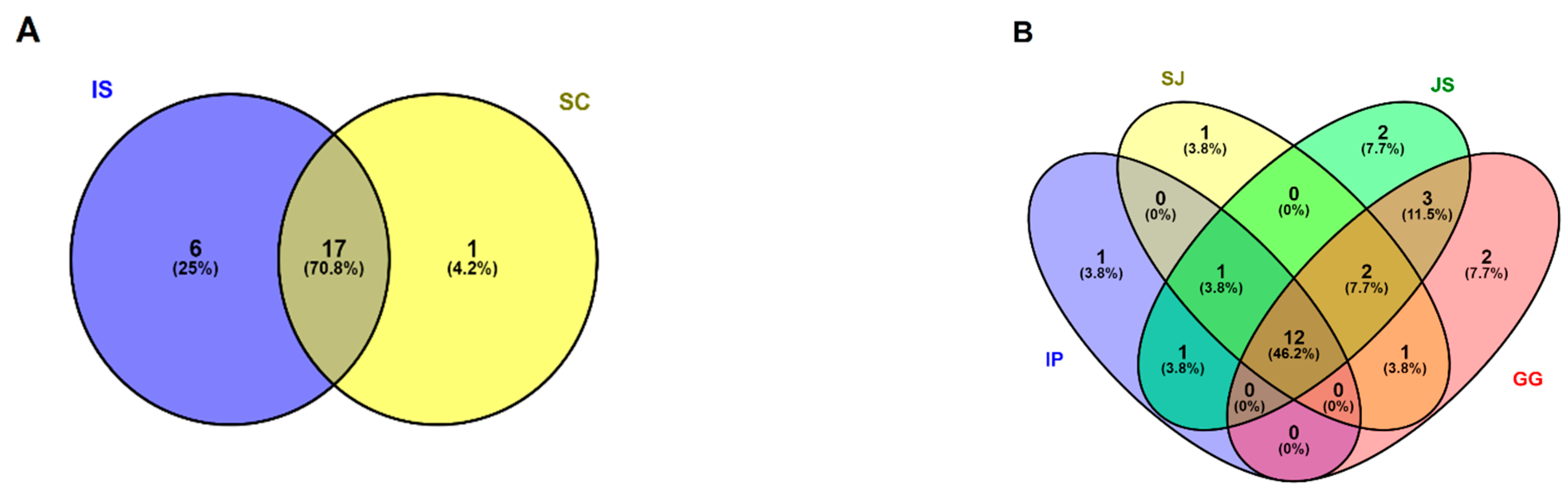
3.2. Composition of Dominant and Unique OTUs in the Gut Profile
3.3. Relationship of Environmental Factors and the Community Composition of Gut Contents
3.4. Assessment of the Habitat Sediment Condition through Fungi Communities
4. Discussion
4.1. Overview of the Gut Content and Diet Composition
4.2. Diversity of Gut Content Associated with Food Sources in Habitats
4.3. Diet Contents Exhibiting the Habitat Condition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bista, I.; Carvalho, G.R.; Walsh, K.; Seymour, M.; Hajibabaei, M.; Lallias, D.; Christmas, M.; Creer, S. Annual time-series analysis of aqueous eDNA reveals ecologically relevant dynamics of lake ecosystem biodiversity. Nat. Commun. 2017, 8, 14087. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, A.A.; Bonnici, B.L.; Yusuf, S.H.; Cruz, J.I.; Hudson, P.L.; Ram, J.L. Improved Chironomid Barcode Database Enhances Identification of Water Mite Dietary Content. Diversity 2022, 14, 65. [Google Scholar] [CrossRef]
- Floss, E.C.S.; Secretti, E.; Kotzian, C.B.; Spies, M.R.; Pires, M.M. Spatial and Temporal Distribution of Non-Biting Midge Larvae Assemblages in Streams in a Mountainous Region in Southern Brazil. J. Insect Sci. 2013, 13, 1–27. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, A.; Armitage, P.; Cranston, P.S.; Pinder, L.C.V. The Chironomidae. The Biology and Ecology of Non-Biting Midges; Chapman & Hall: London, UK, 1995; Volume 64, ISBN 9789401043083. [Google Scholar]
- Na, Y.-K.; Jo, H.; Park, J.-W.; Chang, K.-H.; Kwak, I.-S. The Gut Content Analysis of Polypedilum scalaenum in the Large-scale Weirs of 4 Major River Ecosystems. Korean J. Ecol. Environ. 2020, 53, 55–62. [Google Scholar] [CrossRef]
- Lemes-Silva, A.L.; Pagliosa, P.R.; Petrucio, M.M. Inter- and intra-guild patterns of food resource utilization by chironomid larvae in a subtropical coastal lagoon. Limnology 2014, 15, 1–12. [Google Scholar] [CrossRef]
- Butakka, C.M.M.; Ragonha, F.H.; Train, S.; Pinha, G.D.; Takeda, A.M. Chironomidae feeding habits in different habitats from a Neotropical floodplain: Exploring patterns in aquatic food webs. Braz. J. Biol. 2016, 76, 117–125. [Google Scholar] [CrossRef]
- Aydin, G.B.; Öterler, B.; Elipek, B.Ç.; Guher, H. The Comparative Gut Content Analysis of Some Chironomidae Larvae Living in the Freshwaters at Northern Thrace Region of Turkey. J. Limnol. Freshw. Fish. Res. 2020, 7, 14–23. [Google Scholar] [CrossRef]
- Öterler, B.; Aydin, G.B.; Elipek, B. Qualitative and Quantitative Examination of Algal Flora in the Gut Content of Tanypus punctipennis Meigen, 1818. J. Nat. Appl. Sci. 2018, 22, 88–94. [Google Scholar] [CrossRef]
- Tarkowska-Kukuryk, M. Periphytic algae as food source for grazing chironomids in a shallow phytoplankton-dominated lake. Limnologica 2013, 43, 254–264. [Google Scholar] [CrossRef]
- Kornijów, R.; Markiyanova, M.; Lange, E. Feeding by two closely related species of Chironomus (Diptera: Chironomidae) in south Baltic lagoons, with implications for competitive interactions and resource partitioning. Aquat. Ecol. 2019, 53, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Galizzi, M.C.; Zilli, F.L.; Marchese, M. Diet and functional feeding groups of Chironomidae (Diptera) in the Middle Paraná River floodplain (Argentina). Iheringia-Ser. Zool. 2012, 102, 117–121. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Kielgast, J.; Iversen, L.L.; Wiuf, C.; Rasmussen, M.; Gilbert, M.T.P.; Orlando, L.; Willerslev, E. Monitoring endangered freshwater biodiversity using environmental DNA. Mol. Ecol. 2012, 21, 2565–2573. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Masuda, R.; Sato, Y.; Sado, T.; Araki, H.; Kondoh, M.; Minamoto, T.; Miya, M. Environmental DNA metabarcoding reveals local fish communities in a species-rich coastal sea. Sci. Rep. 2017, 7, 40368. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Xiao, N.; Zhao, Z.; Guo, N.; Luo, Z.; Sun, G.; Li, J. eDNA metabarcoding as a promising conservation tool to monitor fish diversity in Beijing water systems compared with ground cages. Sci. Rep. 2022, 12, 11113. [Google Scholar] [CrossRef]
- Drummond, A.J.; Newcomb, R.D.; Buckley, T.R.; Xie, D.; Dopheide, A.; Potter, B.C.; Heled, J.; Ross, H.A.; Tooman, L.; Grosser, S.; et al. Evaluating a multigene environmental DNA approach for biodiversity assessment. GigaScience 2015, 4, 46. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A Method for Studying Protistan Diversity Using Massively Parallel Sequencing of V9 Hypervariable Regions of Small-Subunit Ribosomal RNA Genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Cordier, T.; Angeles, I.B.; Henry, N.; Lejzerowicz, F.; Berney, C.; Morard, R.; Brandt, A.; Cambon-Bonavita, M.-A.; Guidi, L.; Lombard, F.; et al. Patterns of eukaryotic diversity from the surface to the deep-ocean sediment. Sci. Adv. 2022, 8, eabj9309. [Google Scholar] [CrossRef]
- Sildever, S.; Laas, P.; Kolesova, N.; Lips, I.; Lips, U.; Nagai, S. Plankton biodiversity and species co-occurrence based on environmental DNA—A multiple marker study. Metabarcoding Metagenomics 2021, 5, e72371. [Google Scholar] [CrossRef]
- Minerovic, A.D.; Potapova, M.G.; Sales, C.M.; Price, J.R.; Enache, M.D. 18S-V9 DNA metabarcoding detects the effect of water-quality impairment on stream biofilm eukaryotic assemblages. Ecol. Indic. 2020, 113, 106225. [Google Scholar] [CrossRef]
- Govindarajan, A.F.; Francolini, R.D.; Jech, J.M.; Lavery, A.C.; Llopiz, J.K.; Wiebe, P.H.; Zhang, W. (Exploring the Use of Environmental DNA (eDNA) to Detect Animal Taxa in the Mesopelagic Zone. Front. Ecol. Evol. 2021, 9, 574877. [Google Scholar] [CrossRef]
- Cowart, D.A.; Matabos, M.; Brandt, M.I.; Marticorena, J.; Sarrazin, J. Exploring Environmental DNA (eDNA) to Assess Biodiversity of Hard Substratum Faunal Communities on the Lucky Strike Vent Field (Mid-Atlantic Ridge) and Investigate Recolonization Dynamics After an Induced Disturbance. Front. Mar. Sci. 2020, 6, 783. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, H. Combining Multiple Markers in Environmental DNA Metabarcoding to Assess Deep-Sea Benthic Biodiversity. Front. Mar. Sci. 2021, 8, 1188. [Google Scholar] [CrossRef]
- Kim, D.-K.; Park, K.; Jo, H.; Kwak, I.-S. Comparison of Water Sampling between Environmental DNA Metabarcoding and Conventional Microscopic Identification: A Case Study in Gwangyang Bay, South Korea. Appl. Sci. 2019, 9, 3272. [Google Scholar] [CrossRef]
- Berry, T.E.; Osterrieder, S.K.; Murray, D.C.; Coghlan, M.L.; Richardson, A.J.; Grealy, A.K.; Stat, M.; Bejder, L.; Bunce, M. DNA metabarcoding for diet analysis and biodiversity: A case study using the endangered Australian sea lion (Neophoca cinerea). Ecol. Evol. 2017, 7, 5435–5453. [Google Scholar] [CrossRef] [PubMed]
- Siegenthaler, A.; Wangensteen, O.S.; Soto, A.Z.; Benvenuto, C.; Corrigan, L.; Mariani, S. Metabarcoding of shrimp stomach content: Harnessing a natural sampler for fish biodiversity monitoring. Mol. Ecol. Resour. 2019, 19, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.D.; Questel, J.M.; Maas, K.R.; Bucklin, A. Metabarcoding analysis of regional variation in gut contents of the copepod Calanus finmarchicus in the North Atlantic Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2020, 180, 104738. [Google Scholar] [CrossRef]
- Díaz-Abad, L.; Bacco-Mannina, N.; Madeira, F.M.; Neiva, J.; Aires, T.; Serrao, E.A.; Regalla, A.; Patrício, A.R.; Frade, P.R. eDNA metabarcoding for diet analyses of green sea turtles (Chelonia mydas). Mar. Biol. 2022, 169, 1–12. [Google Scholar] [CrossRef]
- Jo, H.; Choi, B.; Park, K.; Kim, W.-S.; Kwak, I.-S. First Gut Content Analysis of 4th Instar Midge Larvae (Diptera: Chronomidae) In Large-Scale Weirs Using a DNA Meta-Barcoding Approach. Int. J. Environ. Res. Public Health 2020, 17, 2856. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 2016, e2584. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 13 June 2022).
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; ISBN 9780252725487. [Google Scholar]
- Chao, A.; Lee, S.-M. Estimating the Number of Classes via Sample Coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Levins, R. Evolution in Changing Environments; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- Hurlbert, S.H. The Measurement of Niche Overlap and Some Relatives. Ecology 1978, 59, 67–77. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the Species Traits Environment Relationships: The Fourth-Corner Problem Revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef]
- Ward, J.H., Jr. Hierarchical Grouping to Optimize an Objective Function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Beals, E.W. Bray-Curtis Ordination: An Effective Strategy for Analysis of Multivariate Ecological Data. Adv. Ecol. Res. 1984, 14, 1–55. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Albert, J.S.; Destouni, G.; Duke-Sylvester, S.M.; Magurran, A.E.; Oberdorff, T.; Reis, R.E.; Winemiller, K.O.; Ripple, W.J. Scientists’ warning to humanity on the freshwater biodiversity crisis. Ambio 2021, 50, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Lodge, D.M.; Turner, C.R.; Jerde, C.; Barnes, M.; Chadderton, L.; Egan, S.P.; Feder, J.L.; Mahon, A.R.; Pfrender, M.E. Conservation in a cup of water: Estimating biodiversity and population abundance from environmental DNA. Mol. Ecol. 2012, 21, 2555–2558. [Google Scholar] [CrossRef]
- Iacolina, L.; Lukassen, M.B.; Fløjgaard, C.; Buttenschøn, R.; Nielsen, J.L.; Pertoldi, C. eDNA and metabarcoding for rewilding projects monitoring, a dietary approach. Mamm. Biol. 2020, 100, 411–418. [Google Scholar] [CrossRef]
- Ingala, M.R.; Simmons, N.B.; Wultsch, C.; Krampis, K.; Provost, K.L.; Perkins, S.L. Molecular diet analysis of neotropical bats based on fecal DNA metabarcoding. Ecol. Evol. 2021, 11, 7474–7491. [Google Scholar] [CrossRef]
- Lejeune, B.; Mouchet, M.A.; Mehault, S.; Kopp, D. Gut content metabarcoding reveals potential importance of fisheries discards consumption in marine fauna. Can. J. Fish. Aquat. Sci. 2021, 79, 1–10. [Google Scholar] [CrossRef]
- Huryn, A. Aquatic Insects—Ecology, Feeding, and Life History. Encycl. Inl. Waters 2009, 132–143. [Google Scholar] [CrossRef]
- Rodríguez-Barreras, R.; Godoy-Vitorino, F.; Præbel, K.; Wangensteen, O.S. DNA metabarcoding unveils niche overlapping and competition among Caribbean sea urchins. Reg. Stud. Mar. Sci. 2020, 40, 101537. [Google Scholar] [CrossRef]
- Ceriani-Nakamurakare, E.; Mc Cargo, P.; Gonzalez-Audino, P.; Ramos, S.; Carmarán, C. New insights into fungal diversity associated with Megaplatypus mutatus: Gut mycobiota. Symbiosis 2020, 81, 127–137. [Google Scholar] [CrossRef]
- Gouba, N.; Raoult, D.; Drancourt, M. Plant and Fungal Diversity in Gut Microbiota as Revealed by Molecular and Culture Investigations. PLoS ONE 2013, 8, e59474. [Google Scholar] [CrossRef]
- Zhang, Y.; Pavlovska, M.; Stoica, E.; Prekrasna, I.; Yang, J.; Slobodnik, J.; Zhang, X.; Dykyi, E. Holistic pelagic biodiversity monitoring of the Black Sea via eDNA metabarcoding approach: From bacteria to marine mammals. Environ. Int. 2020, 135, 105307. [Google Scholar] [CrossRef]
- Moreno-Pino, M.; Krock, B.; De la Iglesia, R.; Echenique-Subiabre, I.; Pizarro, G.; Vásquez, M.; Trefault, N. Next Generation Sequencing and mass spectrometry reveal high taxonomic diversity and complex phytoplankton-phycotoxins patterns in Southeastern Pacific fjords. Toxicon 2018, 151, 5–14. [Google Scholar] [CrossRef]
- Gudmundsson, G.; von Schmalensee, M.; Svavarsson, J. Are foraminifers (Protozoa) important food for small isopods (Crustacea) in the deep sea? Deep Sea Res. Part I Oceanogr. Res. Pap. 2000, 47, 2093–2109. [Google Scholar] [CrossRef]
- Ptatscheck, C.; Putzki, H.; Traunspurger, W. Impact of deposit-feeding chironomid larvae (Chironomus riparius) on meiofauna and protozoans. Freshw. Sci. 2017, 36, 796–804. [Google Scholar] [CrossRef]
- Ptatscheck, C.; Kreuzinger-Janik, B.; Putzki, H.; Traunspurger, W. Insights into the importance of nematode prey for chironomid larvae. Hydrobiologia 2015, 757, 143–153. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Sabia, A.; Giovanardi, M.; Ferroni, L.; Maglie, M.; Pancaldi, S. Chlorophyta microalgae as dietary protein supplement: A comparative analysis of productivity related to photosynthesis. J. Appl. Phycol. 2022, 34, 1323–1340. [Google Scholar] [CrossRef]
- Kociolek, J.P.; Spaulding, S.A.; Lowe, R.L. Bacillariophyceae: The Raphid Diatoms; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780123858771. [Google Scholar]
- Olefeld, J.L.; Bock, C.; Jensen, M.; Vogt, J.C.; Sieber, G.; Albach, D.; Boenigk, J. Centers of endemism of freshwater protists deviate from pattern of taxon richness on a continental scale. Sci. Rep. 2020, 10, 14431. [Google Scholar] [CrossRef] [PubMed]
- Stefani, F.O.; Klimaszewski, J.; Morency, M.-J.; Bourdon, C.; Labrie, P.; Blais, M.; Venier, L.; Séguin, A. Fungal community composition in the gut of rove beetles (Coleoptera: Staphylinidae) from the Canadian boreal forest reveals possible endosymbiotic interactions for dietary needs. Fungal Ecol. 2016, 23, 164–171. [Google Scholar] [CrossRef]
- Urbina, H.; Schuster, J.; Blackwell, M. The gut of Guatemalan passalid beetles: A habitat colonized by cellobiose- and xylose-fermenting yeasts. Fungal Ecol. 2013, 6, 339–355. [Google Scholar] [CrossRef]
- Sen, K.; Sen, B.; Wang, G. Diversity, Abundance, and Ecological Roles of Planktonic Fungi in Marine Environments. J. Fungi 2022, 8, 491. [Google Scholar] [CrossRef] [PubMed]
- Limatola, N.; Chun, J.T.; Santella, L. Effects of Salinity and pH of Seawater on the Reproduction of the Sea Urchin Paracentrotus lividus. Biol. Bull. 2020, 239, 13–23. [Google Scholar] [CrossRef]
- Chen, Z.; Yuan, J.; Sun, F.; Zhang, F.; Chen, Y.; Ding, C.; Shi, J.; Li, Y.; Yao, L. Planktonic fungal community structures and their relationship to water quality in the Danjiangkou Reservoir, China. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Leray, M.; Knowlton, N. Censusing marine eukaryotic diversity in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150331. [Google Scholar] [CrossRef]
- Su, C.; Hu, W.; Hu, Z.; Zhang, Z.; Wedchaparn, O.; Zhao, L.; Liu, Q. Comparison of high-throughput sequencing analysis of gut contents between silver carp Hypophthalmichthys molitrix and bighead carp Hypophthalmichthys nobilis in mesotrophic and eutrophic lakes. Mar. Freshw. Res. 2020, 71, 761–770. [Google Scholar] [CrossRef]
- Shirey, P.D.; Cowley, D.E.; Sallenave, R. Diatoms from gut contents of museum specimens of an endangered minnow suggest long-term ecological changes in the Rio Grande (USA). J. Paleolimnol. 2008, 40, 263–272. [Google Scholar] [CrossRef]
- Blazewicz-Paszkowycz, M.; Ligowski, R. Diatoms as food source indicator for some Antarctic Cumacea and Tanaidacea (Crustacea). Antarct. Sci. 2002, 14, 11–15. [Google Scholar] [CrossRef]
- Eivarsson, M.; Schnürer, A.; Bengtson, S.; Neubeck, A. Anaerobic Fungi: A Potential Source of Biological H2 in the Oceanic Crust. Front. Microbiol. 2016, 7, 674. [Google Scholar] [CrossRef] [PubMed]
- Kagami, M.; Amano, Y.; Ishii, N. Community Structure of Planktonic Fungi and the Impact of Parasitic Chytrids on Phytoplankton in Lake Inba, Japan. Microb. Ecol. 2011, 63, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, T.; Chen, S. Ignored sediment fungal populations in water supply reservoirs are revealed by quantitative PCR and 454 pyrosequencing. BMC Microbiol. 2015, 15, 44. [Google Scholar] [CrossRef]
- Finlay, B.J.; Esteban, G.F. Freshwater Protozoa: Biodiversity and Ecological Function. Biodivers. Conserv. 1998, 7, 1163–1186. [Google Scholar] [CrossRef]
- Solomon, R.; Wein, T.; Levy, B.; Eshed, S.; Dror, R.; Reiss, V.; Zehavi, T.; Furman, O.; Mizrahi, I.; Jami, E. Protozoa populations are ecosystem engineers that shape prokaryotic community structure and function of the rumen microbial ecosystem. ISME J. 2022, 16, 1187–1197. [Google Scholar] [CrossRef]
- Chang, K.-H.; Nagata, T.; Hanazato, T. Direct and indirect impacts of predation by fish on the zooplankton community: An experimental analysis using tanks. Limnology 2004, 5, 121–124. [Google Scholar] [CrossRef]
- Chang, K.-H.; Doi, H.; Imai, H.; Gunji, F.; Nakano, S.-I. Longitudinal changes in zooplankton distribution below a reservoir outfall with reference to river planktivory. Limnology 2008, 9, 125–133. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Specificity | Primer Sequence (5′-3′) | Length (bp) | Reference |
|---|---|---|---|---|
| 1380F | eukaryotic | GCCTCCCTCGCGCCATCAGXXXXXCCCTGCCHTTTGTACACAC | 43 | [17] |
| 1510R | eukaryotic | GCCTTGCCAGCCCGCTCAGCCTTCYGCAGGTTCACCTAC | 39 |
| Samples | OTUs | Good’s Coverage (%) | Chao Index (S) | Niche Breadth (BA) | |
|---|---|---|---|---|---|
| River | SC | 4066 | 100 | 6341.05 | 0.25 |
| IS | 6361 | 100 | 8023.51 | 0.27 | |
| IP | 2004 | 100 | 2646.82 | 0.13 | |
| Weir | SJ | 2031 | 100 | 2608.67 | 0.12 |
| JS | 2014 | 100 | 2027.55 | 0.06 | |
| GG | 1372 | 100 | 3023.92 | 0.11 |
| Groups | Phylum | Class | River Samples | Weir Samples | ||||
|---|---|---|---|---|---|---|---|---|
| SC | IS | IP | SJ | JS | GG | |||
| Aerobic | Ascomycota | Dothideomycetes | 1.27 | 0.88 | 0.62 | 1 | 0.08 | 0.12 |
| Eurotiomycetes | 0.84 | 5.76 | 0.01 | 0.09 | 0.07 | 0.07 | ||
| Leotiomycetes | 0 | 0 | 0.01 | 0 | 0 | 0.08 | ||
| Saccharomycetes | 0.01 | 0.44 | 0.04 | 0.09 | 0.04 | 0.03 | ||
| Sordariomycetes | 0.12 | 0.41 | - | - | - | 0.01 | ||
| Pezizomycetes | - | - | - | 0.01 | - | - | ||
| Unclassified Ascomycota | 0.51 | 0.3 | 0.11 | 0.31 | 0.03 | 0.07 | ||
| Basidiomycota | Agaricomycetes | 0.21 | 0.82 | 0.05 | 0.04 | 0.04 | 0.16 | |
| Exobasidiomycetes | 0.02 | 0.51 | - | 0.03 | - | - | ||
| Malasseziomycetes | 0.29 | 0.48 | 0.27 | 0.32 | - | 0.3 | ||
| Microbotryomycetes | 0.31 | - | 0.05 | 0.28 | 0.05 | 0.04 | ||
| Tremellomycetes | - | - | - | 0.12 | - | 0.07 | ||
| Ustilaginomycetes | 0.01 | 0.22 | - | 0.01 | 0.01 | - | ||
| Unclassified Basidiomycota | - | 0.36 | 0.09 | 0.18 | - | 0.12 | ||
| Chytridiomycota | Chytridiomycetes | - | 0.15 | 0.08 | 0.07 | 0.01 | 0.38 | |
| Cryptomycota | Rozellidea | 0.01 | - | 0.03 | 0.03 | 0.01 | 0.04 | |
| Subtotal | 3.6 | 10.33 | 1.36 | 2.58 | 0.34 | 1.49 | ||
| Facultative Anaerobic | Peronosporomycetes | Peronosporomycetes | - | 0.58 | 2.28 | 1.31 | 0.7 | 0.13 |
| Anaerobic | Neocallimastigomycota | Neocallimastigomycetes | - | - | - | - | - | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rangaswamy, B.; Ji, C.W.; Kim, W.-S.; Park, J.-W.; Kim, Y.J.; Kwak, I.-S. Profiling Analysis of Filter Feeder Polypedilum (Chironomidae) Gut Contents Using eDNA Metabarcoding Following Contrasting Habitat Types—Weir and Stream. Int. J. Environ. Res. Public Health 2022, 19, 10945. https://doi.org/10.3390/ijerph191710945
Rangaswamy B, Ji CW, Kim W-S, Park J-W, Kim YJ, Kwak I-S. Profiling Analysis of Filter Feeder Polypedilum (Chironomidae) Gut Contents Using eDNA Metabarcoding Following Contrasting Habitat Types—Weir and Stream. International Journal of Environmental Research and Public Health. 2022; 19(17):10945. https://doi.org/10.3390/ijerph191710945
Chicago/Turabian StyleRangaswamy, Boobal, Chang Woo Ji, Won-Seok Kim, Jae-Won Park, Yong Jun Kim, and Ihn-Sil Kwak. 2022. "Profiling Analysis of Filter Feeder Polypedilum (Chironomidae) Gut Contents Using eDNA Metabarcoding Following Contrasting Habitat Types—Weir and Stream" International Journal of Environmental Research and Public Health 19, no. 17: 10945. https://doi.org/10.3390/ijerph191710945