
Transcriptional Responses of Stress-Related Genes in Pale Chub (Zacco platypus) Inhabiting Different Aquatic Environments: Application for Biomonitoring Aquatic Ecosystems
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
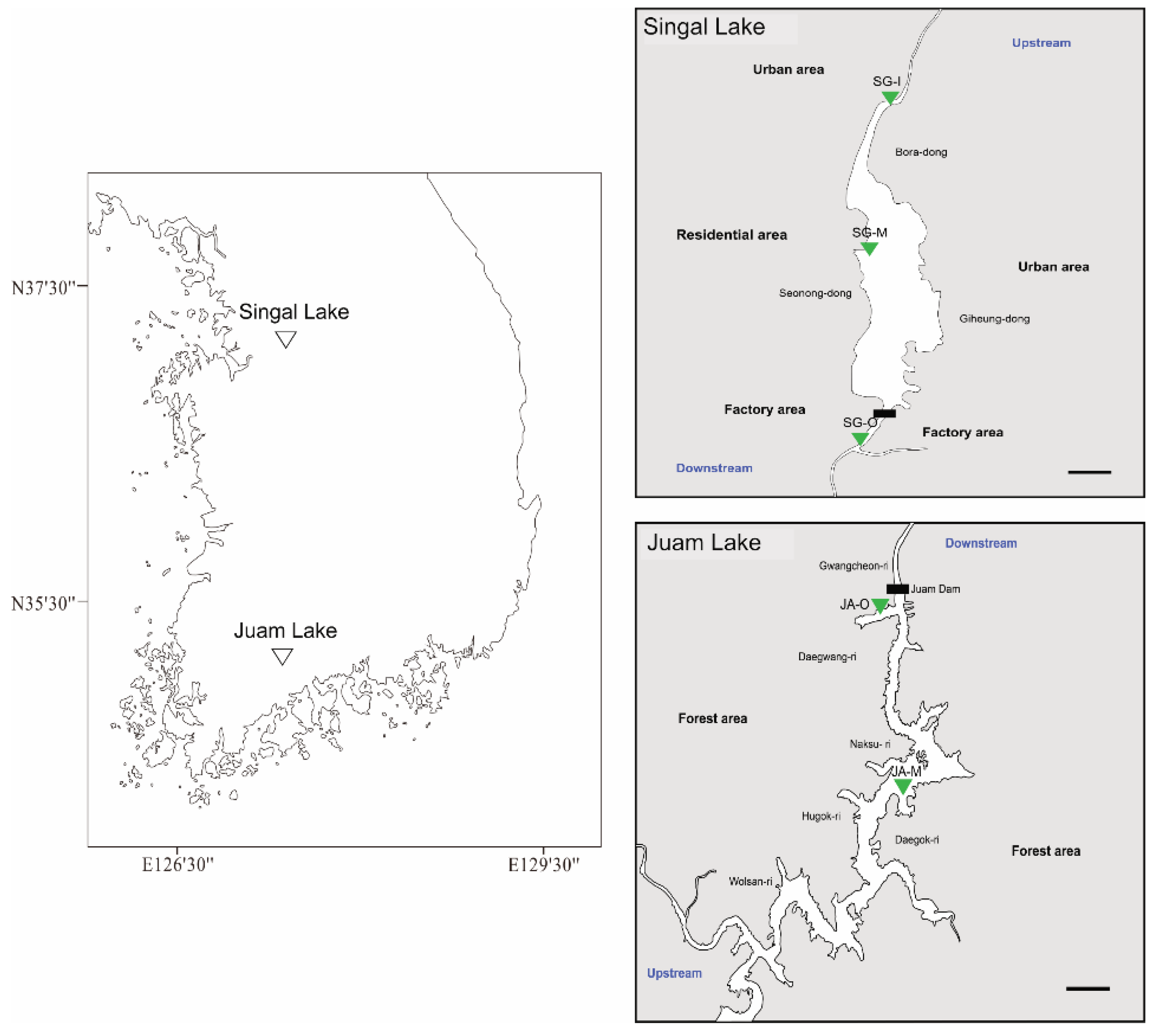
2.1. Sampling Preparation
2.2. Total RNA Extraction and cDNA Synthesis
2.3. Gene Expression Analysis Using Quantitative Real-Time PCR (RT-qPCR)
2.4. Integration of Biomarker Responses (IBR)
2.5. Statistical Analysis
3. Results
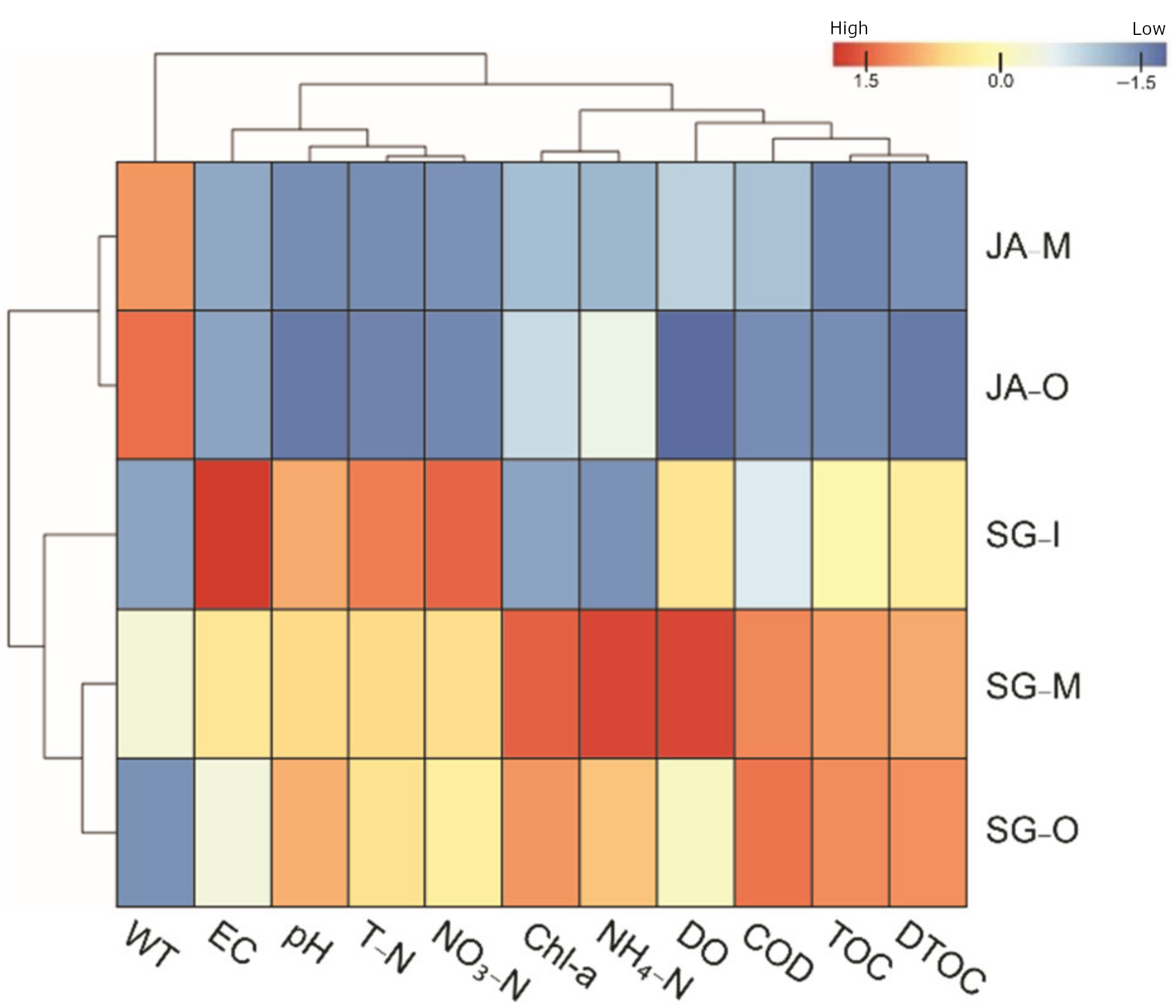
3.1. Water Quality and Hydrological Environments of Lakes
3.2. Transcriptional Responses of Stress-Related Genes in Z. platypus
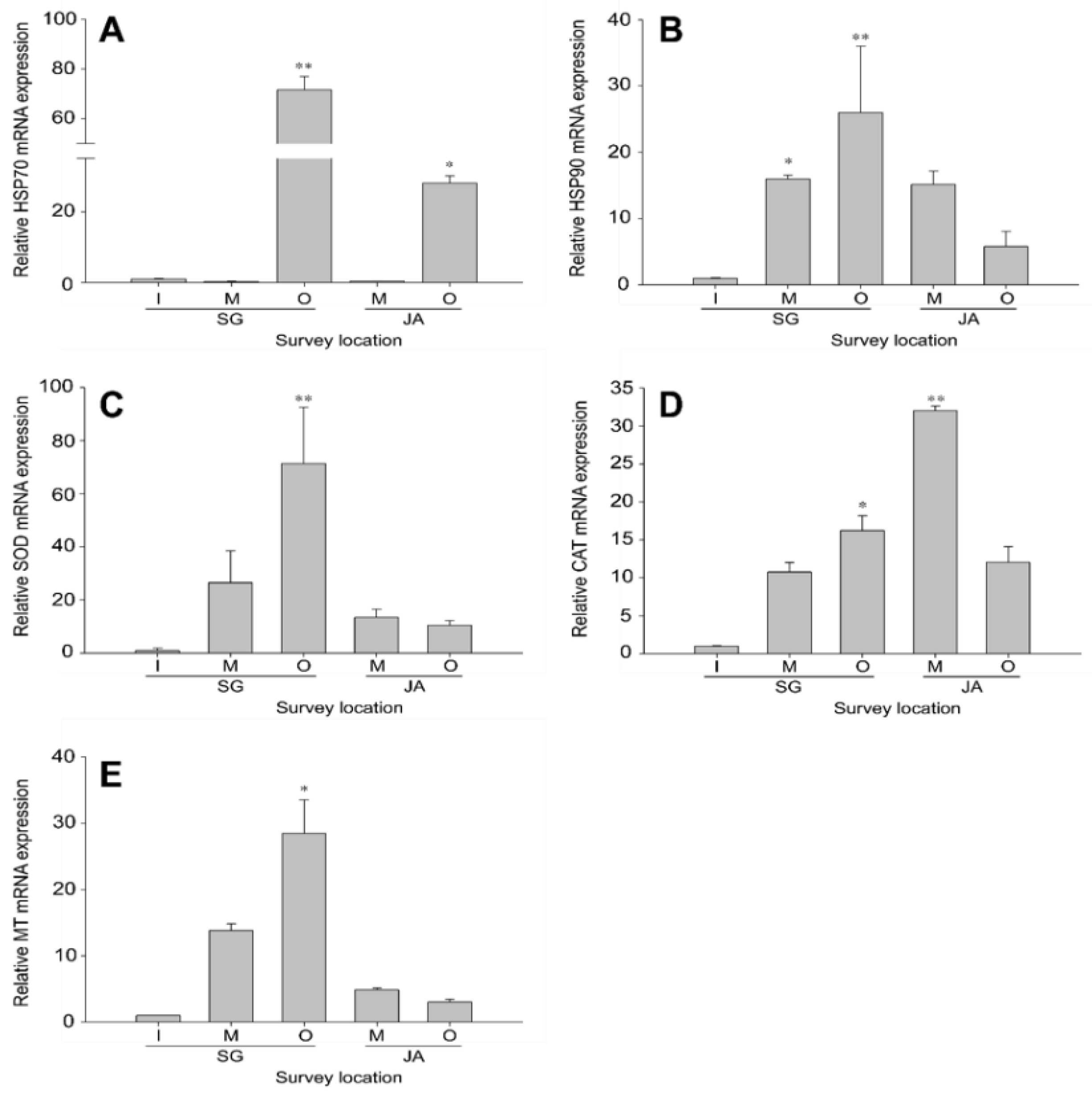
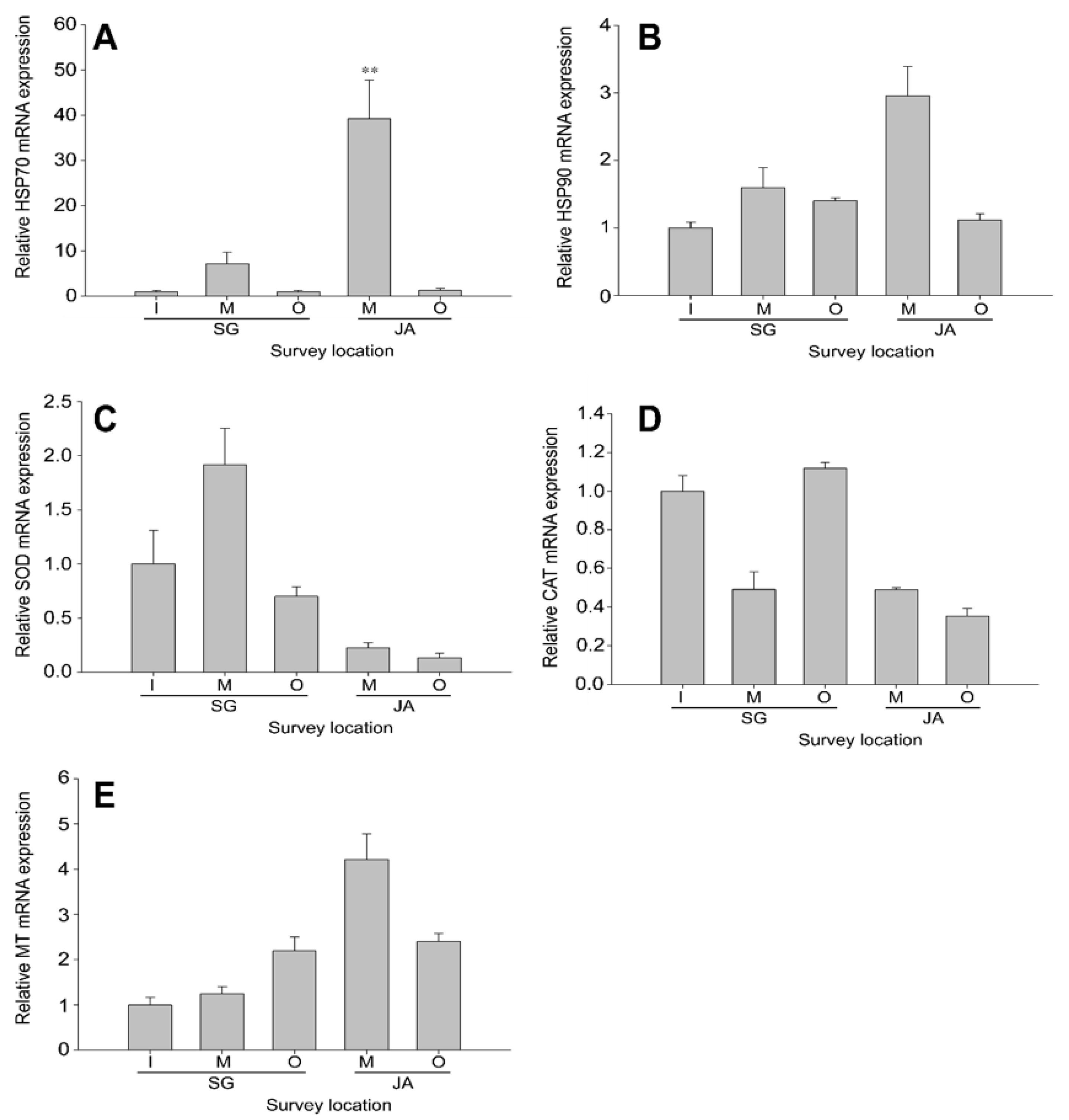
3.2.1. Cellular Homeostasis in Zacco platypus from Different Lakes
3.2.2. Antioxidant Defense in Zacco platypus from Different Lakes
3.2.3. Metal Detoxification in Zacco platypus from Different Lakes
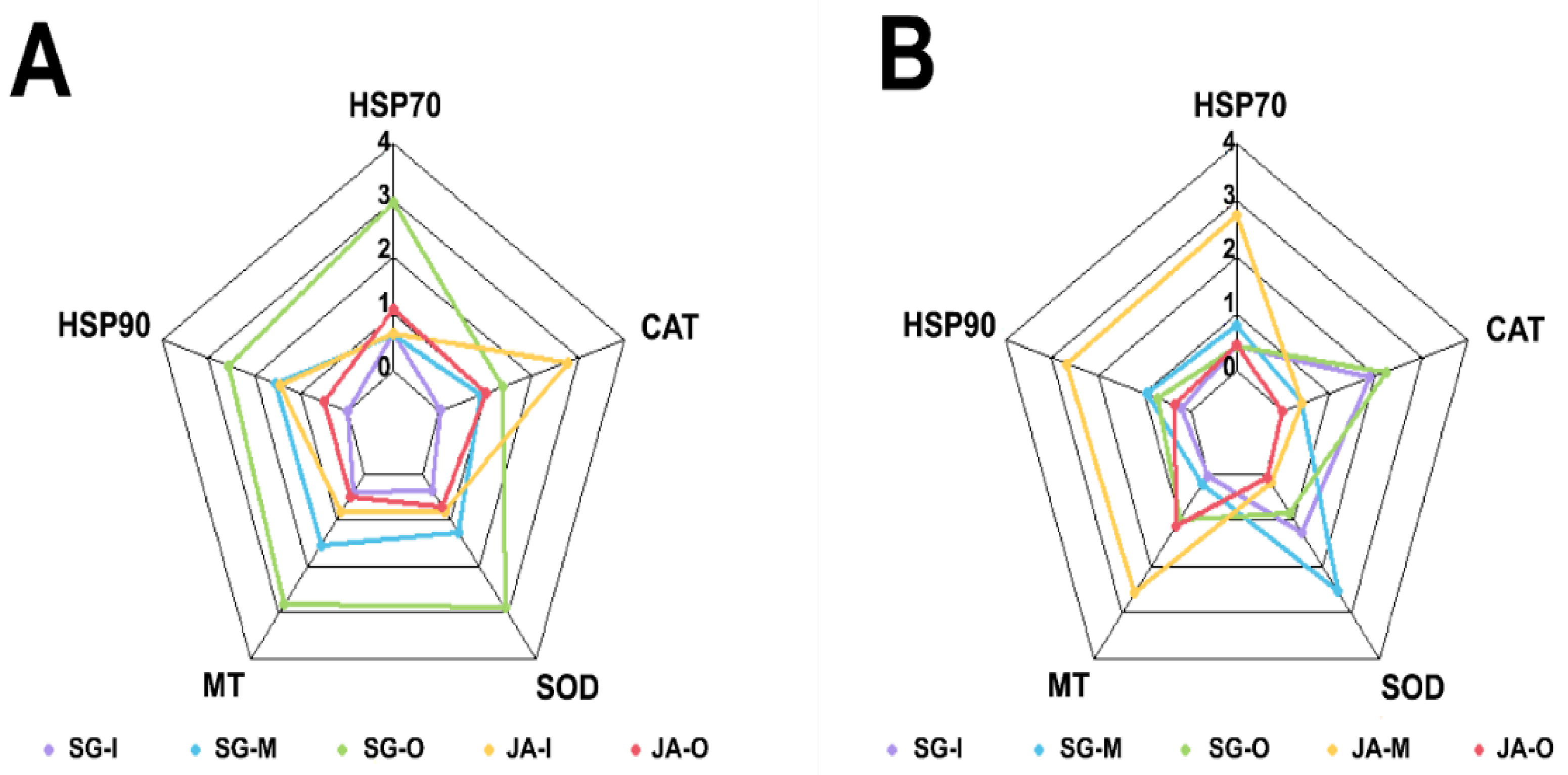
3.3. Integration of Biomarker Responses (IBR) and Heatmap Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cox, K.; Brennan, L.P.; Gerwing, T.G.; Dudas, S.E.; Juanes, F. Sound the alarm: A meta-analysis on the effect of aquatic noise on fish behavior and physiology. Glob. Change Biol. 2018, 24, 3105–3116. [Google Scholar] [CrossRef] [PubMed]
- Adeola, A.O.; Abiodun, B.A.; Adenuga, D.O.; Nomngongo, P.N. Adsorptive and photocatalytic remediation of hazardous organic chemical pollutants in aqueous medium: A review. J. Contam. Hydrol. 2022, 248, 104019. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P.E. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Lee, J.W.; Yoon, H.G.; Lee, S.K. Benzo(a)pyrene-induced cytochrome p4501A expression of four freshwater fishes (Oryzias latipes, Danio rerio, Cyprinus carpio, and Zacco platypus). Environ. Toxicol. Pharmacol. 2015, 39, 1041–1050. [Google Scholar] [CrossRef] [PubMed]
- Park, C.B.; Kim, G.E.; Kim, D.W.; Kim, S.; Yeom, D.H. Biomonitoring the effects of urban-stream waters on the health status of pale chub (Zacco platypus): A comparative analysis of biological indexes and biomarker levels. Ecotoxicol. Environ. Saf. 2021, 208, 111452. [Google Scholar] [CrossRef]
- Cazenave, J.; Bacchetta, C.; Rossi, A.; Ale, A.; Campana, M.; Parma, M.J. Deleterious effects of wastewater on the health status of fish: A field caging study. Ecol. Indic. 2014, 38, 104–112. [Google Scholar] [CrossRef]
- Kim, W.K.; Jung, J. In situ impact assessment of wastewater effluents by integrating multi-level biomarker responses in the pale chub (Zacco platypus). Ecotoxicol. Environ. Saf. 2016, 128, 246–251. [Google Scholar] [CrossRef]
- Samanta, P.; Im, H.; Yoo, J.; Lee, H.; Kim, N.Y.; Kim, W.; Hwang, S.J.; Kim, W.K.; Jung, J. Comparative assessment of the adverse outcome of wastewater effluents by integrating oxidative stress and histopathological alterations in endemic fish. J. Hazard. Mater. 2018, 344, 81–89. [Google Scholar] [CrossRef]
- Kim, W.K.; Lee, S.K.; Jung, J. Integrative assessment of biomarker responses in pale chub (Zacco platypus) exposed to copper and benzo[a]pyrene. Ecotoxicol. Environ. Saf. 2013, 92, 71–78. [Google Scholar] [CrossRef]
- Houde, M.; Giraudo, M.; Douville, M.; Bougas, B.; Couture, P.; DeSilva, A.O.; Spencer, C.; Lair, S.; Verreault, J.; Bernatchez, L.; et al. A multi-level biological approach to evaluate impacts of a major municipal effluent in wild St. Lawrence River yellow perch (Perca flavescens). Sci. Total Environ. 2014, 497–498, 307–318. [Google Scholar] [CrossRef]
- Park, K.; Kwak, I.S. Environmental co-exposure of high temperature and Cu induce hormonal disturbance of cortisol signaling and altered responses of cellular defense genes in zebrafish. Sci. Total Environ. 2022, 842, 156555. [Google Scholar] [CrossRef] [PubMed]
- Booth, L.H.; Bithell, S.L.; Wratten, S.D.; Heppelthwaite, V.J. Vineyard pesticides and their effects on invertebrate biomarkers and bioindicator species in New Zealand. Bull. Environ. Contam. Toxicol. 2003, 71, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Celander, M.C. Cocktail effects on biomarker responses in fish. Aquat. Toxicol. 2011, 105, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Denslow, N.D. Gene expression profiling in fish toxicology: A review. Rev. Environ. Contam. Toxicol. 2017, 241, 1–38. [Google Scholar] [PubMed]
- Park, K.; Kim, W.S.; Kwak, I.S. Effects of di-(2-ethylhexyl) phthalate on transcriptional expression of cellular protection-related HSP60 and HSP67B2 genes in the mud crab Macrophthalmus japonicus. Appl. Sci. 2020, 10, 2766. [Google Scholar] [CrossRef]
- Albornoz-Abud, N.A.; Canul-Marín, G.F.; Chan-Cuá, I.; Hernández-Núñez, E.; Cañizares-Martínez, M.A.; Valdés-Lozano, D.; Rodríguez-Canul, R.; Albores-Medina, A.; Colli-Dula, R.C. Gene expression analysis on growth, development and toxicity pathways of male Nile tilapia (Oreochromis niloticus), after acute and sub-chronic benzo (α) pyrene exposures. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 250, 109160. [Google Scholar] [CrossRef]
- Berrebi, P.; Boissin, E.; Fang, F.; Cattaneo-Berrebi, G. Intron polymorphism(EPIC-PCR) reveals phylogeographic structure of Zacco platypus in China: A possible target for aquaculture development. Heredity 2005, 94, 589–598. [Google Scholar] [CrossRef]
- Kim, J.H.; Yeom, D.H.; Kim, J.H.; Kim, W.K.; An, K.G. Regional ecological health or risk assessments of stream ecosystems using biomarkers and bioindicators of target species (pale chub). Water Air Soil Pollut. 2016, 227, 469. [Google Scholar] [CrossRef]
- Simmons, D.B.; Wallschläger, D. A critical review of the biogeochemistry and ecotoxicology of selenium in lotic and lentic environments. Environ. Toxicol. Chem. 2005, 24, 1331–1343. [Google Scholar] [CrossRef]
- Meena, R.A.A.; Sathishkumar, P.; Ameen, F.; Yusoff, A.R.M.; Gu, F.L. Heavy metal pollution in immobile and mobile components of lentic ecosystems-a review. Environ. Sci. Pollut. Res. Int. 2018, 25, 4134–4148. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Beliaeff, B.; Burgeot, T. Integrated biomarker response: A useful tool for ecological risk assessment. Environ. Toxicol. Chem. 2002, 21, 1316–1322. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef] [PubMed]
- Emam, M.; Caballero-Solares, A.; Xue, X.; Umasuthan, N.; Milligan, B.; Taylor, R.G.; Balder, R.; Rise, M.L. Gill and liver transcript expression changes associated with gill damage in Atlantic salmon (Salmo salar). Front. Immunol. 2022, 13, 806484. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Lin, H.R.; Xia, J.H. Differential gene expression profiles and alternative isoform regulations in gill of Nile tilapia in response to acute hypoxia. Mar. Biotechnol. 2017, 19, 551–562. [Google Scholar] [CrossRef]
- Ireland, H.E.; Harding, S.J.; Bonwick, G.A.; Jones, M.; Smith, C.J.; Williams, J.H.H. Evaluation of heat shock protein 70 as a biomarker of environmental stress in Fucus serratus and Lemna minor. Biomarkers 2004, 9, 139–155. [Google Scholar] [CrossRef]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Amiard, J.C.; Amiard-Triquet, C.; Barka, S.; Pellerin, J.; Rainbow, P.S. Metallothioneins in aquatic invertebrates: Their role in metal detoxification and their use as biomarkers. Aquat. Toxicol. 2006, 76, 160–202. [Google Scholar] [CrossRef]
- Mijošek, T.; Marijić, V.F.; Dragun, Z.; Krasnići, N.; Ivanković, D.; Erk, M. Evaluation of multi-biomarker response in fish intestine as an initial indication of anthropogenic impact in the aquatic karst environment. Sci. Total Environ. 2019, 660, 1079–1090. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, C.D.S.; Bernusso, V.A.; Araújo, H.S.S.D.; Espíndola, E.L.G.; Fernandes, M.N. Biomarker responses as indication of contaminant effects in Oreochromis niloticus. Chemosphere 2012, 89, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.K.; Lee, S.K.; Jung, J. Integrated assessment of biomarker responses in common carp (Cyprinus carpio) exposed to perfluorinated organic compounds. J. Hazard. Mater. 2010, 180, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Maulvault, A.L.; Barbosa, V.; Alves, R.; Anacleto, P.; Camacho, C.; Cunha, S.; Fernandes, J.O.; Ferreira, P.P.; Rosa, R.; Marques, A.; et al. Integrated multi-biomarker responses of juvenile seabass to diclofenac, warming and acidification co-exposure. Aquat. Toxicol. 2018, 202, 65–79. [Google Scholar] [CrossRef]
- Beghin, M.; Schmitz, M.; Betoulle, S.; Palluel, O.; Baekelandt, S.; Mandiki, S.N.M.; Gillet, E.; Nott, K.; Porcher, J.M.; Robert, C.; et al. Integrated multi-biomarker responses of juvenile rainbow trout (Oncorhynchus mykiss) to an environmentally relevant pharmaceutical mixture. Ecotoxicol. Environ. Saf. 2021, 221, 112454. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Tm (°C) | Efficiency (%) | Reference |
|---|---|---|---|---|
| GAPDH | F: ACGCGGAAGGCCATACC | 53 | 90 | Kim and Jung, 2016 [7] |
| R: GGCTGTGGGCAAAGTCATTC | ||||
| HSP70 | F: CCTCAATGGTCCTGGTGAAG | 61 | 96 | KY926431 |
| R: TCACCTTCTGCCCCAGATAA | ||||
| HSP90 | F: TGGTGTGGGCTTCTACTCTG | 60 | 88 | KM201321 |
| R: CCCTCTTCTCCTCGACGTAC | ||||
| SOD | F: GAAGGAGGATGACTTGGGTAAGG | 62 | 90 | KF515699 |
| R: CCGGCGTTGCCAGTTTTA | ||||
| CAT | F: AAATCCGCAGACTCACCTAAA | 53 | 89 | KF515698 |
| R: GGACGCAAACCCCAGAAA | ||||
| MT | F: GATTGCGCCAAGACTGGAA | 60 | 91 | KC952875 |
| R: CTGGCAGTTAGTGCACTTGCA |
| Lake | Hydrological Factors | Physicochemical Factors | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Basin Area (km2) | Reservoir Area (km2) | Maximum Water Level (EL·m) | Place | Depth (m) | WT (°C) | Do (mg·L−1) | pH | EC (µmhos/cm) | COD (mg·L−1) | TOC (mg·L−1) | TN (mg·L−1) | Chl-a (mg·m−3) | NO3-N (mg·L−1) | NH3-N (mg·L−1) | DTOC (mg·L−1) | |
| Singal (SG) | 53 | 2.3 | 46 | Inflow (I) | 0.4 | 17.4 | 12.9 | 8.3 | 723.0 | 4.3 | 3.0 | 4.3 | 5.2 | 2.9 | 0.1 | 2.8 |
| Middle (M) | 5.0 | 19.4 | 15.1 | 8.1 | 439.3 | 5.9 | 3.4 | 3.5 | 21.3 | 2.1 | 0.4 | 3.1 | ||||
| Outflow (O) | 0.5 | 17.1 | 11.8 | 8.3 | 273.0 | 6.0 | 3.5 | 3.4 | 19.1 | 2.0 | 0.3 | 3.3 | ||||
| Average | 5.9 | 18.0 | 13.3 | 8.2 | 478.4 | 5.4 | 3.3 | 3.7 | 15.2 | 2.3 | 0.2 | 3.1 | ||||
| Juam (JA) | 1010 | 33 | 108.5 | Middle (M) | 36.6 | 22.2 | 10.3 | 6.2 | 78.3 | 3.9 | 3.9 | 0.9 | 6.4 | 0.4 | 0.1 | 1.7 |
| Outflow (O) | 38.5 | 22.8 | 8.8 | 6.1 | 70.5 | 3.5 | 2.2 | 0.9 | 8.2 | 0.4 | 0.2 | 1.6 | ||||
| Average | 37.6 | 22.5 | 9.6 | 6.1 | 74.4 | 3.7 | 2.1 | 0.9 | 7.3 | 0.4 | 0.1 | 1.6 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.-S.; Park, K.; Park, J.-W.; Lee, S.-H.; Kim, J.-H.; Kim, Y.-J.; Oh, G.-H.; Ko, B.-S.; Park, J.-W.; Hong, C.; et al. Transcriptional Responses of Stress-Related Genes in Pale Chub (Zacco platypus) Inhabiting Different Aquatic Environments: Application for Biomonitoring Aquatic Ecosystems. Int. J. Environ. Res. Public Health 2022, 19, 11471. https://doi.org/10.3390/ijerph191811471
Kim W-S, Park K, Park J-W, Lee S-H, Kim J-H, Kim Y-J, Oh G-H, Ko B-S, Park J-W, Hong C, et al. Transcriptional Responses of Stress-Related Genes in Pale Chub (Zacco platypus) Inhabiting Different Aquatic Environments: Application for Biomonitoring Aquatic Ecosystems. International Journal of Environmental Research and Public Health. 2022; 19(18):11471. https://doi.org/10.3390/ijerph191811471
Chicago/Turabian StyleKim, Won-Seok, Kiyun Park, Jae-Won Park, Sun-Ho Lee, Ji-Hoon Kim, Yong-Jun Kim, Gun-Hee Oh, Bong-Soon Ko, Ji-Won Park, Cheol Hong, and et al. 2022. "Transcriptional Responses of Stress-Related Genes in Pale Chub (Zacco platypus) Inhabiting Different Aquatic Environments: Application for Biomonitoring Aquatic Ecosystems" International Journal of Environmental Research and Public Health 19, no. 18: 11471. https://doi.org/10.3390/ijerph191811471