
Genome Analysis of a Novel Polysaccharide-Degrading Bacterium Paenibacillus algicola and Determination of Alginate Lyases

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Screening and Identification of Strain HB172198T
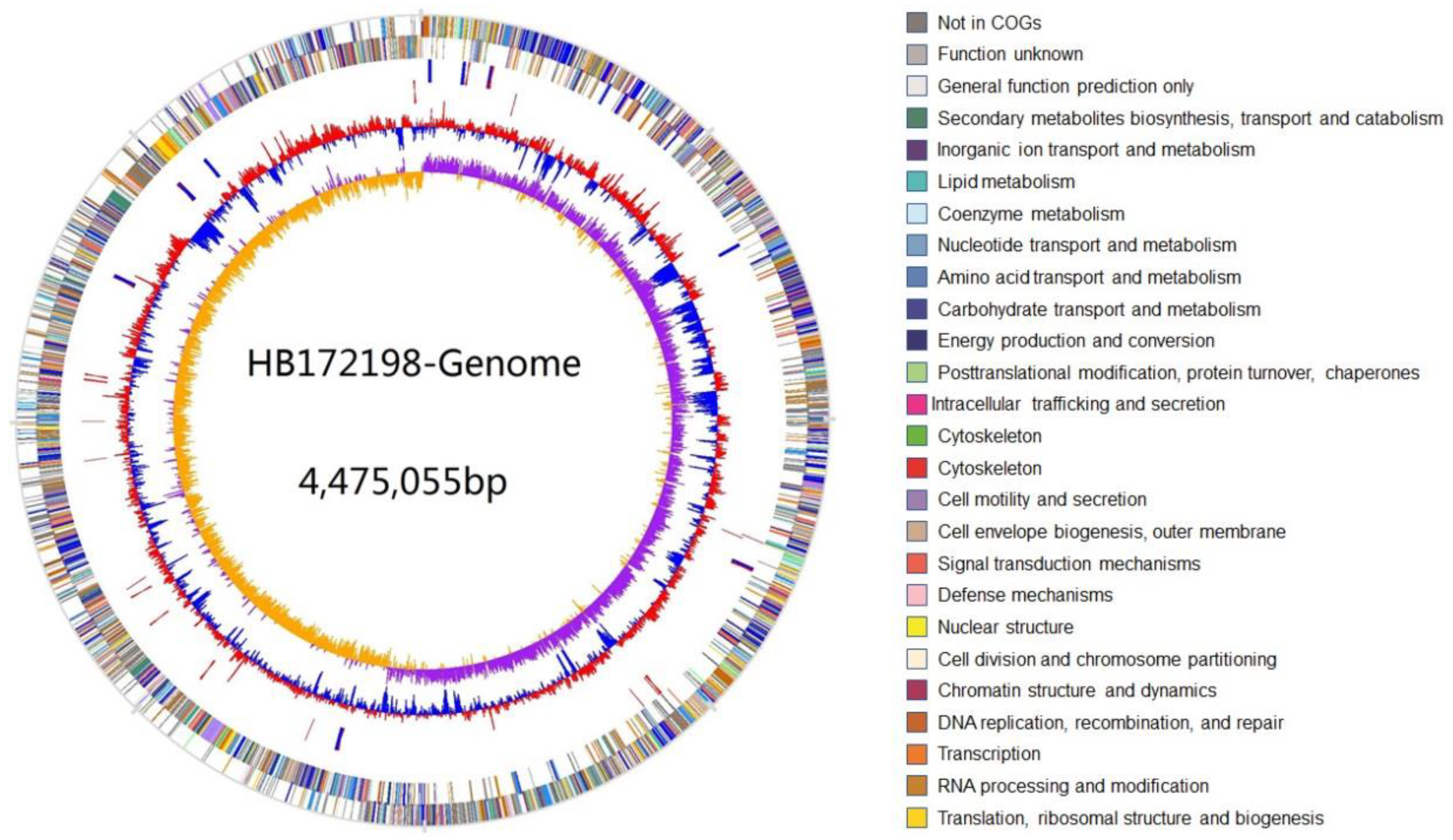
2.2. Genome Specifics
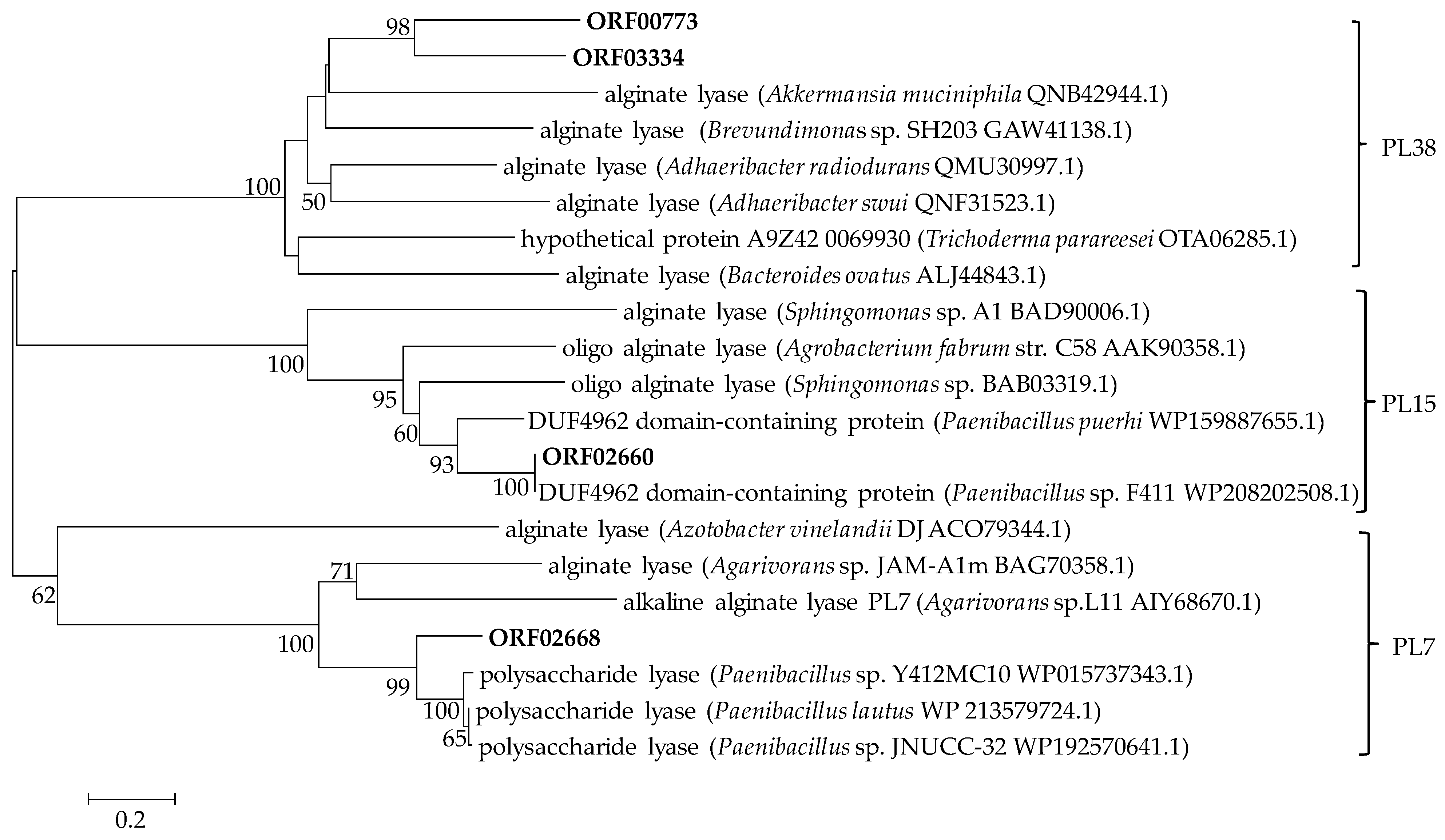
2.3. Genetic Basis of Polysaccharide Degradation
2.4. Test of Carbohydrate Utilization
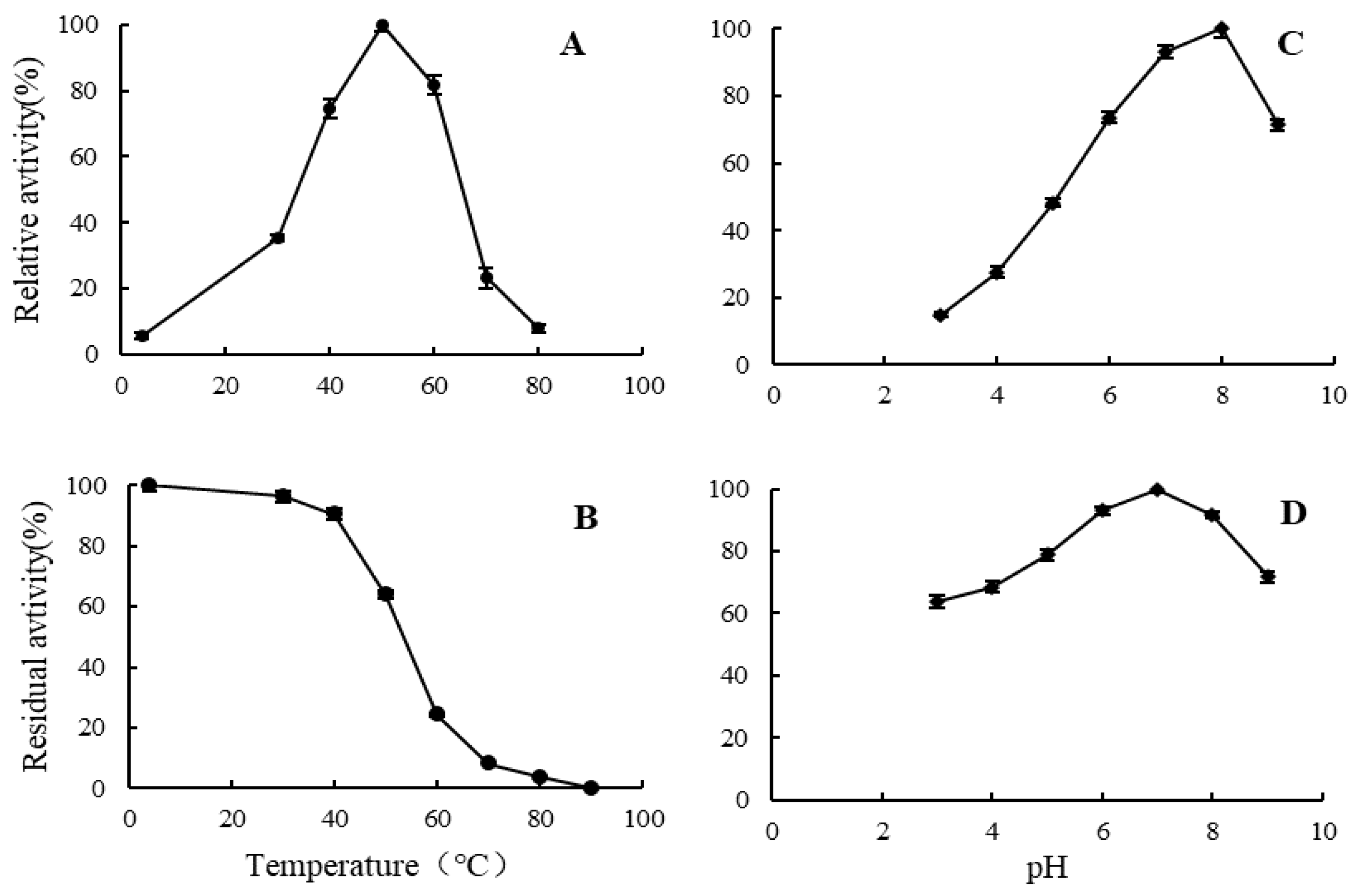
2.5. Enzymatic Properties of Alginate Lyase
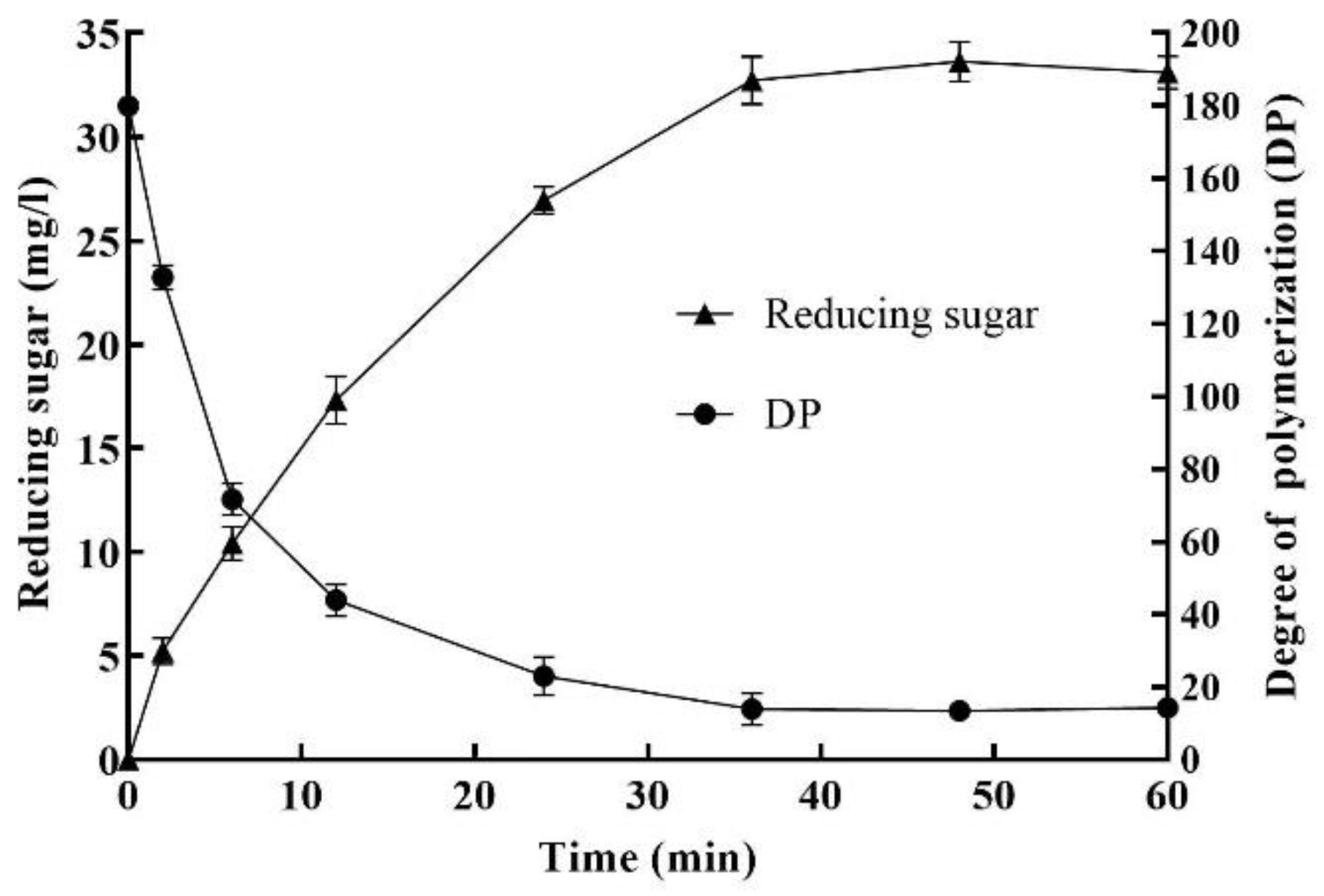
2.6. Enzymatic Degradation of Sodium Alginate
3. Materials and Methods
3.1. Materials and Strains
3.2. Screening and Identification of Strain HB172198T
3.3. Genome Sequencing and Annotation
3.4. Utilizing Abilities of Carbohydrate
3.5. Detection of Alginate Lyase Activity and Enzymatic Properties
3.6. Preparation and Detection of the Enzymatic Degradation of Alginate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, B.L.; Jin, M.; Li, L.; Qu, W.; Zeng, R.Y. Genome sequencing reveals the complex polysaccharide-degrading ability of novel deep-sea bacterium Flammeovirga pacifica WPAGA1. Front. Microbiol. 2017, 8, 600–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martone, P.T.; Estevez, J.M.; Lu, F.; Ruel, K.; Denny, M.W.; Somerville, C.; Ralph, J. Discovery of lignin in seaweed reveals convergent evolution of cell-wall architecture. Curr. Biol. 2009, 19, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.P.; Chen, L.; Zhang, Z.J.; Wang, X.J.; Qin, S.; Yan, P.S. Screening of alginate lyase-excreting microorganisms from the surface of brown algae. AMB Express 2017, 7, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, T.; Preston, L.; Schiller, N. Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar] [CrossRef]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [Green Version]
- Rhein-Knudsen, N.; Meyer, A.S. Chemistry, gelation, and enzymatic modification of seaweed food hydrocolloids. Trends Food Sci. Technol. 2021, 109, 608–621. [Google Scholar] [CrossRef]
- Chen, X.L.; Dong, S.; Xu, F.; Dong, F.; Li, P.Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a new cold-adapted and salt-activated polysaccharide lyase family 7 alginate lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120–1128. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.H.; Yin, H.Y.; Zhao, X.M.; Wang, W.X.; Du, Y.G.; He, A.L.; Sun, K.G. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr. Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef]
- Yang, J.H.; Bang, M.A.; Jang, C.H.; Jo, G.H.; Jung, S.K.; Ki, S.H. Alginate oligosaccharide enhances LDL uptake via regulation of LDLR and PCSK9 expression. J. Nutr. Biochem. 2015, 26, 1393–1400. [Google Scholar] [CrossRef]
- Ghadam, P.; Akhlaghi, F.; Ali, A.A. One-step purification and characterization of alginate lyase from a clinical Pseudomonas aeruginosa with destructive activity on bacterial biofilm. Iran J. Basic Med. Sci. 2017, 20, 467–473. [Google Scholar]
- Vidal-Melgosa, S.; Lagatora, M.; Sichert, A.; Priest, T.; Pätzold, J.; Hehemann, J.H. Not digested: Algal glycans move carbon dioxide into the deep-sea. bioRxiv 2022. [Google Scholar] [CrossRef]
- Neumann, A.M.; Balmonte, J.P.; Berger, M.; Giebel, H.A.; Arnosti, C.; Voget, S.; Simon, M.; Brinkhoff, T.; Wietz, M. Different utilization of alginate and other algal polysaccharides by marine Alteromonas macleodii ecotypes. Environ. Microbiol. 2015, 17, 3857–3868. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.K.; Gong, Y.; Shang, D.D.; Liu, B.T.; Du, Z.J.; Chen, G.J. Degradation of alginate by a newly isolated marine bacterium Agarivorans sp. B2Z047. Mar. Drug. 2022, 20, 254. [Google Scholar] [CrossRef]
- Zhu, J.; Mo, K.L.; Zheng, Z.G.; Wang, Z.X.; Hu, Y.H.; Zou, X.X.; Gu, H.J.; Wang, H.Y.; Bao, S.X.; Huang, H.Q. Paenibacillus algicola sp. nov., a novel marine bacterium producing alginate lyase. Int. J. Syst. Evol. Microbiol. 2020, 70, 5087–5092. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Barbeyron, T.; Martin, R.; Portetelle, D.; Michel, G.; Vandenbol, M. The cultivable surface microbiota of the brown alga Ascophyllum nodosum is enriched in macroalgal-polysaccharide-degrading bacteria. Front. Microbiol. 2015, 24, 1487–1500. [Google Scholar] [CrossRef]
- Chernysheva, N.; Bystritskaya, E.; Likhatskaya, G.; Nedashkovskaya, O.; Isaeva, M. Genome-wide analysis of PL7 alginate lyases in the genus Zobellia. Molecules 2021, 26, 2387. [Google Scholar] [CrossRef]
- Das, S.N.; Wagenknecht, M.; Nareddy, P.K.; Bhuvanachandra, B.; Niddana, R.; Balamurugan, R.; Swamy, M.J.; Moerschbacher, B.M.; Podile, A.R. Amino groups of chitosan are crucial for binding to a family 32 carbohydrate binding module of a chitosanase from Paenibacillus elgii. J. Biol. Chem. 2016, 36, 18977–18990. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.J.; Yang, Y.J.; Tang, X.H.; Shen, J.D.; Xu, B.; Li, J.J.; Wu, Q.; Zhou, J.P.; Ding, J.M.; Han, N.Y.; et al. NaCl-, protease-tolerant and cold-active endoglucanase from Paenibacillus sp. YD236 isolated from the feces of Bos frontalis. Springerplus 2016, 5, 746–757. [Google Scholar] [CrossRef] [Green Version]
- Phakeenuya, V.; Ratanakhanokchai, K.; Kosugi, A.; Tachaapaikoon, C. A novel multifunctional GH9 enzyme from Paenibacillus curdlanolyticus B-6 exhibiting endo/exo functions of cellulase, mannanase and xylanase activities. Appl. Microbiol. Biotechnol. 2020, 104, 2079–2096. [Google Scholar] [CrossRef]
- Ashraf, S.; Soudi, M.R.; Ghadam, P. Production of xanthanases by Paenibacillus spp.: Complete xanthan segradation and possible applications. Iran J. Biotechnol. 2017, 15, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Itoh, T. Structures and functions of carbohydrate-active enzymes of chitinolytic bacteria Paenibacillus sp. str. FPU-7. Biosci. Biotechnol. Biochem. 2021, 85, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.P.; Chen, L.; Lou, Z.Y.; Yuan, X.T.; Pan, G.P.; Ren, X.Y.; Wang, P.Y. Cloning and characterization of a novel alginate lyase from Paenibacillus sp. LJ-23. Mar. Drugs 2022, 20, 66. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Cheng, Y.Y.; Gu, J.Y.; Wang, Y.H.; Li, J.G.; Li, F.C.; Han, W.J. Draft genome sequence of Paenibacillus sp. strain MY03, a terrestrial bacterium capable of degrading multiple marine-derived polysaccharides. Genome Announc. 2017, 5, e00678-17. [Google Scholar] [CrossRef] [Green Version]
- Kurakake, M.; Kitagawa, Y.H.; Okazaki, A.; Shimizu, K. Enzymatic properties of alginate lyase from Paenibacillus sp. S29. Appl. Biochem. Biotechnol. 2010, 183, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Nakagawa, E.; Yoda, M.; Nakaichi, A.; Hibi, T.; Kimoto, H. Structural and biochemical characterisation of a novel alginate lyase from Paenibacillus sp. str. FPU-7. Sci. Rep. 2019, 9, 14870. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.Z.; Zhao, X.; Liu, L.L.; Yu, G.L. Applications of mass spectrometry to structural analysis of marine oligosaccharides. Mar. Drugs 2014, 12, 4005. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Jiang, C.; Xu, J.; Liu, Z.; Mao, X. Characteristics and applications of alginate lyases. Int. J. Biol. Macromol. 2020, 164, 1304–1320. [Google Scholar] [CrossRef]
- Kam, N.; Park, Y.J.; Lee, E.Y.; Kim, H.S. Molecular identification of a polyM-specific alginate lyase from Pseudomonas sp. strain KS-408 for degradation of glycosidic linkages between two mannuronates or mannuronate and guluronate in alginate. Can. J. Microbiol. 2011, 57, 1032–1041. [Google Scholar] [CrossRef]
- Sun, C.; Chen, Y.J.; Zhang, X.Q.; Pan, J.; Cheng, H.; Wu, M. Draft genome sequence of Microbulbifer elongatus strain HZ11, a brown seaweed-degrading bacterium with potential ability to produce bioethanol from alginate. Mar. Genomics 2014, 18, 83–85. [Google Scholar] [CrossRef]
- Hisano, T.; Nishimura, M.; Yamashita, T.; Imanaka, T.; Muramatsu, T.; Kimura, A.; Murata, K. A simple method for determination of substrate specificity of alginate lyases. J. Fermen. Bioeng. 1994, 78, 182–184. [Google Scholar] [CrossRef]
- Mann, A.J.; Hahnke, R.L.; Huang, S.X.; Werner, J.; Xing, P.; Barbeyron, T.; Huettel, B.; Stuber, K.; Reinhardt, R.; Harder, J.; et al. The genome of the alga-associated marine flavobacterium Formosa agariphila KMM 3901T reveals a broad potential for degradation of algal polysaccharides. Appl. Environ. Microbiol. 2013, 79, 6813–6822. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, M.; Konno, N.; Suzuki, T.; Fujii, Y.; Kodama, Y.; Isogai, A.; Habu, N. A bacterial endo-β-1,4-glucuronan lyase, CUL-I from Brevundimonas sp. SH203, belonging to a novel polysaccharide lyase family. Protein Expr. Purif. 2020, 166, 105502–105506. [Google Scholar] [CrossRef] [PubMed]
- Pilgaard, B.; Vuillemin, M.; Munk, L.; Holck, J.; Meier, S.; Wilkens, C.; Meyer, A.S. Discovery of a novel glucuronan lyase system in Trichoderma parareesei. Appl. Environ. Microbiol. 2022, 88, e01819-21. [Google Scholar] [CrossRef] [PubMed]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.J.; Chang, Y.K.; Hong, S.K. Agar degradation by microorganisms and agar-degrading exzymes. Appl. Microbiol. Biotechnol. 2012, 94, 917–930. [Google Scholar] [CrossRef]
- Ebringerová, A.; Heinze, T. Xylan and xylan derivatives-biopolymers with valuable properties, 1. Naturally occurring xylans structures, isolation procedures and properties. Macromol. Rapid Commun. 2000, 21, 542–556. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Tatusov, R.L.; Natale, D.A.; Garkavtsev, I.V.; Tatusova, T.A.; Shankavaram, U.T.; Rao, B.S.; Kiryutin, B.; Galperin, M.Y.; Fedorova, N.D.; Koonin, E.V. The COG database: New developments in phylogenetic classification of proteins from complete genomes. Nucleic Acids Res. 2001, 29, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Hattori, M.; Aoki-Kinoshita, K.F.; Itoh, M.; Kawashima, S.; Katayama, T.; Araki, M.; Hirakawa, M. From genomics to chemical genomics: New developments in KEGG. Nucleic Acids Res. 2006, 34, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H. Predicting secretory proteins with SignalP. Methods Mol. Biol. 2017, 1611, 59–73. [Google Scholar] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Budi, S.W.; van Tuinen, D.; Arnould, C.; Dumas-Gaudot, E.; Gianinazzi-Pearson, V.; Gianinazzi, S. Hydrolytic enzyme activity of Paenibacillus sp. strain B2 and effects of the antagonistic bacterium on cell integrity of two soil-borne pathogenic fungi. Appl. Soil Ecol. 2000, 15, 191–199. [Google Scholar] [CrossRef]
- Huang, H.Q.; Li, S.; Bao, S.X.; Mo, K.L.; Sun, D.M.; Hu, Y.H. Expression and characterization of a cold-adapted alginate lyase with exo/endo-type activity from a novel marine bacterium Alteromonas portus HB161718T. Mar. Drugs 2021, 19, 155. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid for determina tion of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Albalasmeh, A.A.; Berhe, A.A.; Ghezzehei, T.A. A new method for rapid determination of carbohydrate and total carbon concentrations using UV spectrophotometry. Carbohydr. Polym. 2013, 97, 253–261. [Google Scholar] [CrossRef]
- Kim, D.E.; Lee, E.Y.; Kim, H.S. Cloning and characterization of alginate lyase from a marine bacterium Streptomyces sp. ALG-5. Marine Biotechnol. 2009, 11, 10–16. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Number |
|---|---|
| Genome size (bp) | 4,475,055 |
| G+C content (%) | 51.2% |
| Total genes predicted | 4182 |
| Protein-coding genes | 4001 |
| tRNA genes | 80 |
| rRNA genes | 27 |
| 5S rRNA | 9 |
| 16S rRNA | 9 |
| 23S rRNA | 9 |
| ncRNAs | 4 |
| Pseudo genes (total) | 70 |
| Catabolic Enzymes | Enzyme Family | No. of Enymes |
|---|---|---|
| Alginate lyase | PL7 | 1 |
| PL15 | 1 | |
| PL38 | 2 | |
| β-Agarase | GH50 | 1 |
| GH86 | 1 | |
| ι-Carrageenase | GH82 | 1 |
| β-1,4-Endo-glucanase | GH9 | 1 |
| β-Glucosidase | GH3 | 4 |
| α-Amylase | GH2 | 1 |
| GH13 | 3 | |
| GH13|CBM34 | 1 | |
| Pullulanase | CBM48|GH13|CBM41 | 1 |
| CBM41|CBM41|CBM48|GH13|CBM41|GH13 | 1 | |
| Lichenase | GH16 | 1 |
| Endo-1,4-β-xylanase | CBM22|GH10|CBM9 | 1 |
| CBM22|GH10|CBM9|CBM9 | 1 | |
| GH11 | 1 | |
| Xylan 1,4-β-xylosidase | GH43 | 3 |
| GH52 | 1 | |
| α-Glucosidase | GH4 | 1 |
| GH13 | 1 | |
| CBM34|GH13 | 1 | |
| Heparinase | PL12 | 1 |
| Chitinase | GH18 | 1 |
| α-L-fucosidase | GH29 | 1 |
| Glucan endo-1,3-β-D-glucosidase | CBM54|GH16|CBM4|CBM4|CBM4|CBM4 | 1 |
| Glycosaminoglycan polysaccharide lyase | PL8 | 1 |
| α-Galactosidase | GH4 | 1 |
| β-Galactosidase | GH36 | 1 |
| GH2 | 4 | |
| GHnc|CBM66 | 1 | |
| α-L-rhamnosidase | GH78 | 3 |
| α-Mannosidase | GH38 | 2 |
| GH125 | 1 | |
| α-Phosphotrehalase | GH13 | 1 |
| Arabinanase | GH117 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, H.; Zheng, Z.; Zou, X.; Wang, Z.; Gao, R.; Zhu, J.; Hu, Y.; Bao, S. Genome Analysis of a Novel Polysaccharide-Degrading Bacterium Paenibacillus algicola and Determination of Alginate Lyases. Mar. Drugs 2022, 20, 388. https://doi.org/10.3390/md20060388
Huang H, Zheng Z, Zou X, Wang Z, Gao R, Zhu J, Hu Y, Bao S. Genome Analysis of a Novel Polysaccharide-Degrading Bacterium Paenibacillus algicola and Determination of Alginate Lyases. Marine Drugs. 2022; 20(6):388. https://doi.org/10.3390/md20060388
Chicago/Turabian StyleHuang, Huiqin, Zhiguo Zheng, Xiaoxiao Zou, Zixu Wang, Rong Gao, Jun Zhu, Yonghua Hu, and Shixiang Bao. 2022. "Genome Analysis of a Novel Polysaccharide-Degrading Bacterium Paenibacillus algicola and Determination of Alginate Lyases" Marine Drugs 20, no. 6: 388. https://doi.org/10.3390/md20060388