
Call the Eckols: Present and Future Potential Cancer Therapies


 ,
,  and
and 
Abstract
:1. Introduction
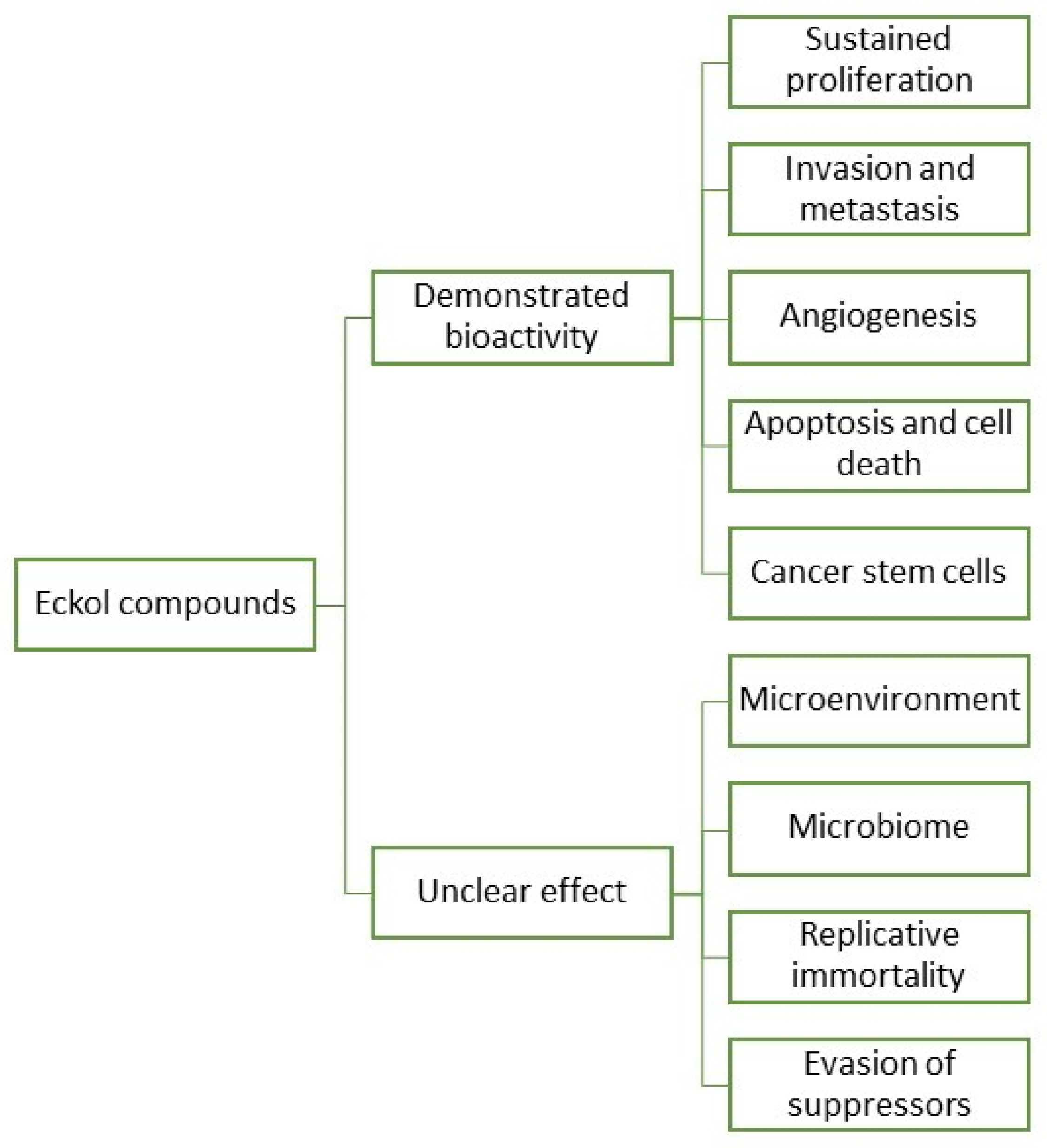
2. Eckols as Potential Agents against Cancer Hallmarks
2.1. Eckols in Sustained Proliferation and Signaling
2.2. Eckols in Invasion and Metastasis
2.3. Eckols in Angiogenesis
2.4. Eckols in Resisting Programmed Cell Death (Apoptosis)
2.5. Eckols in Cancer Cell Stemness
3. Cancer Hallmarks in Which Eckols Can Demonstrate Potential Bioactivities
3.1. The Stromal Role of the Microenvironment
3.2. The Accessory Tumoral Microbiome
3.3. The Characteristic Replicative Immortality of Telomeres
3.4. Evasion of Cellular Suppression Mechanisms
4. Future of Eckols as a Therapeutic
Radioprotective Ability of Eckols
5. Bottlenecks in Eckols Implementation as a Therapeutic Alternative
5.1. Seasonal Variability of Macroalgae Bioactives
5.2. Eckol Extraction and Isolation
5.3. Bioavailability
5.4. In Vitro vs. In Vivo Questions on Eckols
5.5. Eckols Toxicity on Non-Cancerous Cells
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The Human Tumor Microbiome Is Composed of Tumor Type–Specific Intracellular Bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, T.; Steyger, P.S. An Integrated View of Cisplatin-Induced Nephrotoxicity and Ototoxicity. Toxicol. Lett. 2015, 237, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, S.; Zhang, W.; Smid, S.D. Phlorotannins: A Review on Biosynthesis, Chemistry and Bioactivity. Food Biosci. 2021, 39, 100832. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A View on Drug Resistance in Cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [Green Version]
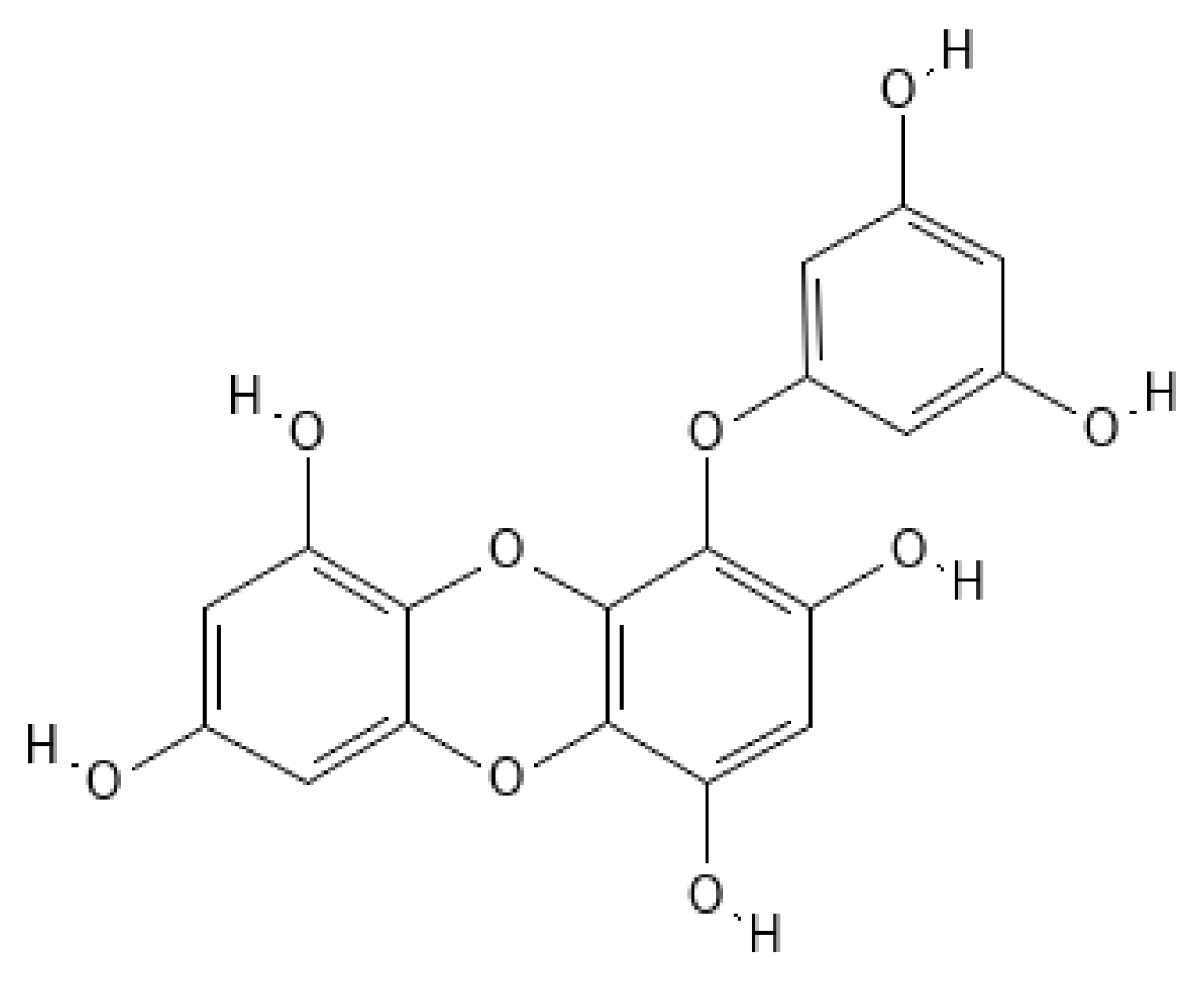
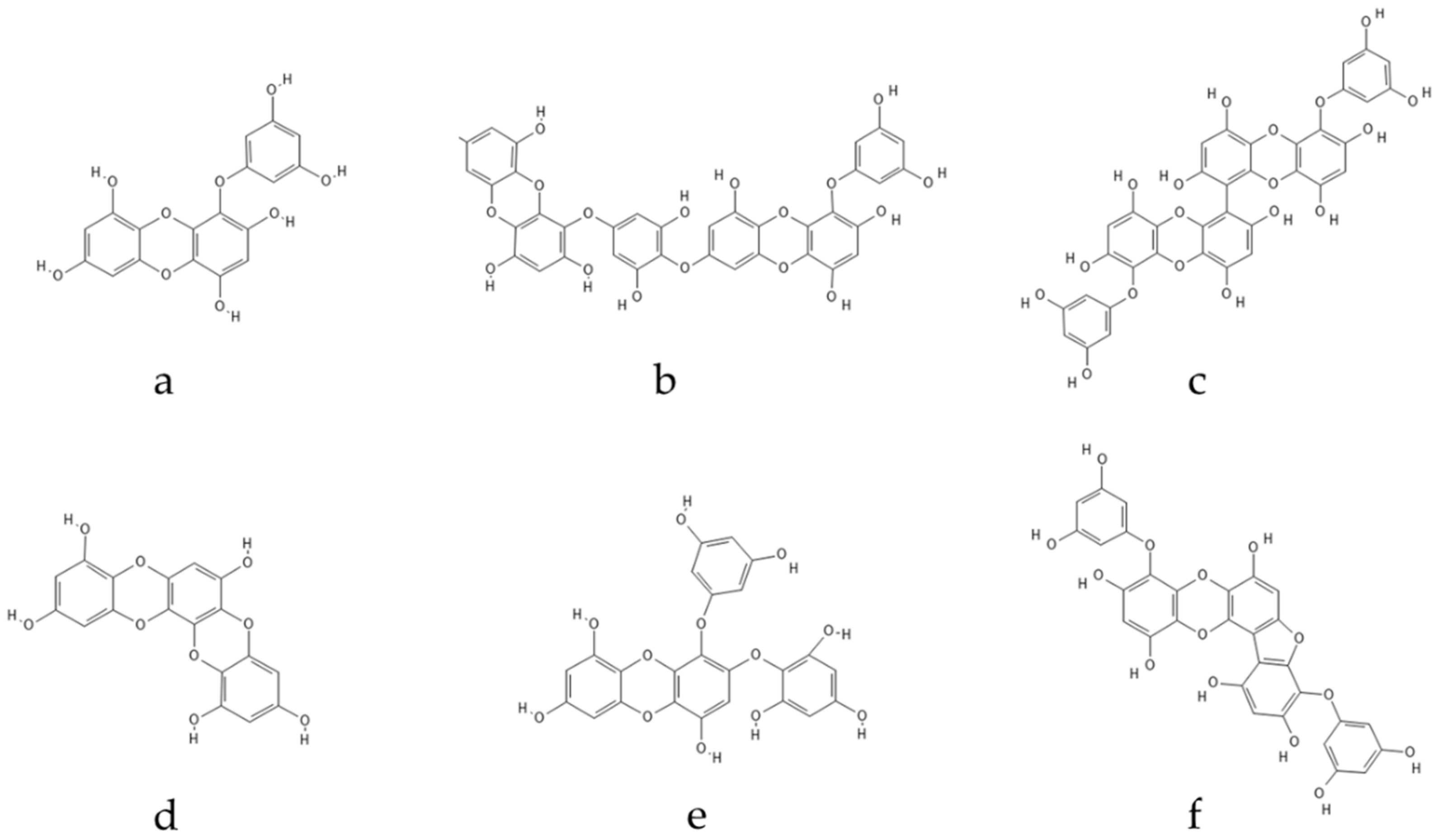
- Manandhar, B.; Paudel, P.; Seong, S.H.; Jung, H.A.; Choi, J.S. Characterizing Eckol as a Therapeutic Aid: A Systematic Review. Mar. Drugs 2019, 17, 361. [Google Scholar] [CrossRef] [Green Version]
- Karadeniz, F.; Kim, S.-K. Antitumor and Antimetastatic Effects of Marine Algal Polyphenols. In Handbook of Anticancer Drugs from Marine Origin; Springer International Publishing: Cham, Switzerland, 2015; pp. 177–183. [Google Scholar]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP Kinase Signalling Pathways in Cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [Green Version]
- Noorolyai, S.; Shajari, N.; Baghbani, E.; Sadreddini, S.; Baradaran, B. The Relation between PI3K/AKT Signalling Pathway and Cancer. Gene 2019, 698, 120–128. [Google Scholar] [CrossRef]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained Proliferation in Cancer: Mechanisms and Novel Therapeutic Targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, W.; Zhao, S.; Li, S.; Yan, D.; Wang, J. Eckol Inhibits Reg3A-induced Proliferation of Human SW1990 Pancreatic Cancer Cells. Exp. Ther. Med. 2019, 18, 2825–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwangi, H.M.; Njue, W.M.; Onani, M.O.; Thovhoghi, N.; Mabusela, W.T. Phlorotannins and a Sterol Isolated from a Brown Alga Ecklonia Maxima, and Their Cytotoxic Activity against Selected Cancer Cell Lines HeLa, H157 and MCF7. Interdiscip. J. Chem. 2017, 2, 1–6. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of Apoptosis by Phloroglucinol Derivative from Ecklonia Cava in MCF-7 Human Breast Cancer Cells. Food Chem. Toxicol. 2009, 47, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-M. Dieckol Inhibits 12-O-Tetradecanoylphorbol-13-Acetate-Induced SK-Hep1 Human Hepatoma Cell Motility through Suppression of Matrix Metalloproteinase-9 Activity. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 376–381. [Google Scholar] [CrossRef]
- Wang, C.H.; Li, X.; Jin, L.; Zhao, Y.; Zhu, G.; Shen, W. Dieckol Inhibits Non-Small-Cell Lung Cancer Cell Proliferation and Migration by Regulating the PI3K/AKT Signaling Pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22346. [Google Scholar] [CrossRef] [Green Version]
- Efferth, T. Stem Cells, Cancer Stem-like Cells, and Natural Products. Planta Med. 2012, 78, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Hyun, K.-H.; Yoon, C.-H.; Kim, R.-K.; Lim, E.-J.; An, S.; Park, M.-J.; Hyun, J.-W.; Suh, Y.; Kim, M.-J.; Lee, S.-J. Eckol Suppresses Maintenance of Stemness and Malignancies in Glioma Stem-like Cells. Toxicol. Appl. Pharmacol. 2011, 254, 32–40. [Google Scholar] [CrossRef]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-MTOR Pathways: Cross-Talk and Compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, M.; Yang, K.; Tian, J. 6,60-Bieckol Induces Apoptosis and Suppresses TGF-b-Induced Epithelial- Mesenchymal Transition in Non-Small Lung Cancer Cells. Chinese Herb. Med. 2021, 14, 254–262. [Google Scholar] [CrossRef]
- Lee, Y.; Park, J.; Park, S.; Joo, N.; Lee, B.H.; Lee, K.B.; Oh, S. Dieckol or Phlorofucofuroeckol Extracted from Ecklonia Cava Suppresses Lipopolysaccharide-Mediated Human Breast Cancer Cell Migration and Invasion. J. Appl. Phycol. 2019, 32, 631–640. [Google Scholar] [CrossRef]
- Jeong, S.-H.; Jeon, Y.-J.; Park, S.J. Inhibitory Effects of Dieckol on Hypoxia-Induced Epithelial-Mesenchymal Transition of HT29 Human Colorectal Cancer Cells. Mol. Med. Rep. 2016, 14, 5148–5154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, W.; Liu, H.; Lei, Y.; Gao, H.; Alahmadi, T.A.; Peng, H.; Chen, W. Chemopreventive Effect of Dieckol against 7,12-dimethylbenz(a)Anthracene Induced Skin Carcinogenesis Model by Modulatory Influence on Biochemical and Antioxidant Biomarkers. Environ. Toxicol. 2021, 36, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Kim, Y.T.; Jeon, Y.J. Antioxidant Dieckol Downregulates the Rac1/ROS Signaling Pathway and Inhibits Wiskott-Aldrich Syndrome Protein (WASP)-Family Verprolin-Homologous Protein 2 (WAVE2)-Mediated Invasive Migration of B16 Mouse Melanoma Cells. Mol. Cells 2012, 33, 363–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Jeon, Y.J. Dieckol from Ecklonia Cava Suppresses the Migration and Invasion of HT1080 Cells by Inhibiting the Focal Adhesion Kinase Pathway Downstream of Rac1-ROS Signaling. Mol. Cells 2012, 33, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, M.; Zhao, S.; Liu, J.; Hu, X. In Vivo Anti-Tumor Effect of Eckol, a Phlorotannin Component Isolated from Brown Algae, Associated with Regulating Dendritic Cells in Sarcoma 180 (S180) Xenografts-Bearing Mice. Proc. Annu. Meet. Jpn. Pharmacol. Soc. 2018, WCP2018, PO1-8-21. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Guo, J.; Hu, X.M.; Zhao, S.Q.; Li, S.L.; Wang, J. An in Vivo Anti-Tumor Effect of Eckol from Marine Brown Algae by Improving the Immune Response. Food Funct. 2019, 10, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-S.; Yadunandam, A.K.; Kim, S.-J.; Woo, H.-C.; Kim, H.-R.; Kim, G.-D. Dieckol, Isolated from Ecklonia Stolonifera, Induces Apoptosis in Human Hepatocellular Carcinoma Hep3B Cells. J. Nat. Med. 2012, 67, 519–527. [Google Scholar] [CrossRef]
- Ahn, J.-H.; Yang, Y.-I.; Lee, K.-T.; Choi, J.-H. Dieckol, Isolated from the Edible Brown Algae Ecklonia Cava, Induces Apoptosis of Ovarian Cancer Cells and Inhibits Tumor Xenograft Growth. J. Cancer Res. Clin. Oncol. 2015, 141, 255–268. [Google Scholar] [CrossRef]
- Sadeeshkumar, V.; Duraikannu, A.; Ravichandran, S.; Kodisundaram, P.; Fredrick, W.S.; Gobalakrishnan, R. Modulatory Efficacy of Dieckol on Xenobiotic-Metabolizing Enzymes, Cell Proliferation, Apoptosis, Invasion and Angiogenesis during NDEA-Induced Rat Hepatocarcinogenesis. Mol. Cell. Biochem. 2017, 433, 195–204. [Google Scholar] [CrossRef]
- Eo, H.J.; Kwon, T.; Park, G.H.; Song, H.M.; Lee, S.; Park, N.; Jeong, J.B. In Vitro Anticancer Activity of Phlorofucofuroeckol A via Upregulation of Activating Transcription Factor 3 against Human Colorectal Cancer Cells. Mar. Drugs 2016, 14, 69. [Google Scholar] [CrossRef] [Green Version]
- Berx, G.; van Roy, F. Involvement of Members of the Cadherin Superfamily in Cancer. Cold Spring Harb. Perspect. Biol. 2009, 1, a003129. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, D.S.; Farabaugh, S.M.; Ford, H.L. Epithelial-Mesenchymal Transition in Cancer: Parallels Between Normal Development and Tumor Progression. J. Mammary Gland Biol. Neoplasia 2010, 15, 117–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shlyakhtina, Y.; Moran, K.L.; Portal, M.M. Genetic and Non-Genetic Mechanisms Underlying Cancer Evolution. Cancers 2021, 13, 1380. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Li, M.; Tan, Y.; Cao, L.; Gu, Q.; Yang, H.; Hu, C. Snail1-Expressing Cancer-Associated Fibroblasts Induce Lung Cancer Cell Epithelial-Mesenchymal Transition through MiR-33b. Oncotarget 2017, 8, 114769–114786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talmadge, J.E.; Fidler, I.J. AACR Centennial Series: The Biology of Cancer Metastasis: Historical Perspective. Cancer Res. 2010, 70, 5649–5669. [Google Scholar] [CrossRef] [Green Version]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The Intestinal Microbiota Modulates the Anticancer Immune Effects of Cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Li, Y.; Qian, Z.-J.; Lee, S.-H.; Li, Y.-X.; Kim, S. Dieckol from Ecklonia Cava Regulates Invasion of Human Fibrosarcoma Cells and Modulates MMP-2 and MMP-9 Expression via NF-ΚB Pathway. Evid.-Based Complement. Altern. Med. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Baeriswyl, V.; Christofori, G. The Angiogenic Switch in Carcinogenesis. Semin. Cancer Biol. 2009, 19, 329–337. [Google Scholar] [CrossRef]
- Ferrara, N. Vascular Endothelial Growth Factor. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 789–791. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, Y.; Xiao, Z.; Tang, Y.; Hong, P.; Sun, S.; Zhou, C.; Qian, Z.-J. Inhibition Effects of 7-Phloro-Eckol from Ecklonia Cava on Metastasis and Angiogenesis Induced by Hypoxia through Regulation of AKT/MTOR and ERK Signaling Pathways. Arab. J. Chem. 2021, 14, 103187. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 Apoptotic Switch in Cancer Development and Therapy. Oncogene 2007, 26, 1324–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junttila, M.R.; Evan, G.I. P53—A Jack of All Trades but Master of None. Nat. Rev. Cancer 2009, 9, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Panchision, D.M.; McKay, R.D.G. The Control of Neural Stem Cells by Morphogenic Signals. Curr. Opin. Genet. Dev. 2002, 12, 478–487. [Google Scholar] [CrossRef]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a Cancer Stem Cell in Human Brain Tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar]
- Galli, R.; Binda, E.; Orfanelli, U.; Cipelletti, B.; Gritti, A.; De Vitis, S.; Fiocco, R.; Foroni, C.; Dimeco, F.; Vescovi, A. Erratum: Isolation and Characterization of Tumorigenic, Stem-like Neural Precursors from Human Glioblastoma (Cancer Research (October 2004) 64 (7011–7021). Cancer Res. 2004, 64, 8130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Garcia, A.J.; Wu, M.; Lawson, D.A.; Witte, O.N.; Wu, H. Pten Deletion Leads to the Expansion of a Prostatic Stem/Progenitor Cell Subpopulation and Tumor Initiation. Proc. Natl. Acad. Sci. USA 2006, 103, 1480–1485. [Google Scholar] [CrossRef] [Green Version]
- Bhowmick, N.A.; Neilson, E.G.; Moses, H.L. Stromal Fibroblasts in Cancer Initiation and Progression. Nature 2004, 432, 332–337. [Google Scholar] [CrossRef]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential Role of Intratumor Bacteria in Mediating Tumor Resistance to the Chemotherapeutic Drug Gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The Fungal Mycobiome Promotes Pancreatic Oncogenesis via Activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Kim, H.J.; Dasagrandhi, C.; Kim, S.H.; Kim, B.G.; Eom, S.H.; Kim, Y.M. In Vitro Antibacterial Activity of Phlorotannins from Edible Brown Algae, Eisenia Bicyclis Against Streptomycin-Resistant Listeria Monocytogenes. Indian J. Microbiol. 2018, 58, 105–108. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial Action of Compounds from Marine Seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasco, M.A. Telomeres and Human Disease: Ageing, Cancer and Beyond. Nat. Rev. Genet. 2005, 6, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, D.L.; Sage, J. Cellular Mechanisms of Tumour Suppression by the Retinoblastoma Gene. Nat. Rev. Cancer 2008, 8, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, B.J.; Strasser, A.; Kelly, G.L. Tumor-Suppressor Functions of the TP53 Pathway. Cold Spring Harb. Perspect. Med. 2016, 6, a026062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curto, M.; Cole, B.K.; Lallemand, D.; Liu, C.-H.; McClatchey, A.I. Contact-Dependent Inhibition of EGFR Signaling by Nf2/Merlin. J. Cell Biol. 2007, 177, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Partanen, J.I.; Nieminen, A.I.; Klefstrom, J. 3D View to Tumor Suppression: Lkb1, Polarity and the Arrest of Oncogenic c-Myc. Cell Cycle 2009, 8, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Ikushima, H.; Miyazono, K. TGFβ Signalling: A Complex Web in Cancer Progression. Nat. Rev. Cancer 2010, 10, 415–424. [Google Scholar] [CrossRef]
- Mohan, G.; Ayisha Hamna, T.P.; Jijo, A.J.; Saradha Devi, K.M.; Narayanasamy, A.; Vellingiri, B. Recent Advances in Radiotherapy and Its Associated Side Effects in Cancer—a Review. J. Basic Appl. Zool. 2019, 80, 14. [Google Scholar] [CrossRef]
- Park, E.; Ahn, G.; Lee, N.H.; Kim, J.M.; Yun, J.S.; Hyun, J.W.; Jeon, Y.J.; Wie, M.B.; Lee, Y.J.; Park, J.W.; et al. Radioprotective Properties of Eckol against Ionizing Radiation in Mice. FEBS Lett. 2008, 582, 925–930. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Kang, K.A.; Piao, M.J.; Ko, D.O.; Wang, Z.H.; Lee, I.K.; Kim, B.J.; Jeong, I.Y.; Shin, T.; Park, J.W.; et al. Eckol Protects V79-4 Lung Fibroblast Cells against γ-Ray Radiation-Induced Apoptosis via the Scavenging of Reactive Oxygen Species and Inhibiting of the c-Jun NH2-Terminal Kinase Pathway. Eur. J. Pharmacol. 2008, 591, 114–123. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Lee, N.H.; Joo, H.G.; Jee, Y. Modulation of Apoptosis of Eckol against Ionizing Radiation in Mice. Biochem. Biophys. Res. Commun. 2008, 372, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Sadeeshkumar, V.; Duraikannu, A.; Aishwarya, T.; Jayaram, P.; Ravichandran, S.; Ganeshamurthy, R. Radioprotective Efficacy of Dieckol against Gamma Radiation-Induced Cellular Damage in Hepatocyte Cells. Naunyn. Schmiedebergs Arch. Pharmacol. 2019, 392, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Kang, K.A.; Hyun, J.W. Effect of Dieckol on Y-Ray Radiation-Induced V79-4 Lung Fibroblast Damage Involved in Modulation of Reactive Oxygen Species. J. Med. Life Sci. 2009, 6, 368–372. [Google Scholar] [CrossRef]
- Moon, C.; Kim, S.-H.; Kim, J.-C.; Hyun, J.W.; Lee, N.H.; Park, J.W.; Shin, T. Protective Effect of Phlorotannin Components Phloroglucinol and Eckol on Radiation-Induced Intestinal Injury in Mice. Phyther. Res. 2008, 22, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Lomartire, S.; Cotas, J.; Pacheco, D.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Environmental Impact on Seaweed Phenolic Production and Activity: An Important Step for Compound Exploitation. Mar. Drugs 2021, 19, 245. [Google Scholar] [CrossRef]
- Domínguez, H. Algae as a Source of Biologically Active Ingredients for the Formulation of Functional Foods and Nutraceuticals. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 1–19. [Google Scholar]
- Liu, M.; Hansen, P.E.; Lin, X. Bromophenols in Marine Algae and Their Bioactivities. Mar. Drugs 2011, 9, 1273–1292. [Google Scholar] [CrossRef] [Green Version]
- Singh, I.P.; Sidana, J. Phlorotannins. In Functional Ingredients from Algae for Foods and Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2013; pp. 181–204. [Google Scholar]
- Wei, R.; Lee, M.-S.; Lee, B.; Oh, C.-W.; Choi, C.-G.; Kim, H.-R. Isolation and Identification of Anti-Inflammatory Compounds from Ethyl Acetate Fraction of Ecklonia Stolonifera and Their Anti-Inflammatory Action. J. Appl. Phycol. 2016, 28, 3535–3545. [Google Scholar] [CrossRef]
- Ahn, G.-N.; Kim, K.-N.; Cha, S.-H.; Song, C.-B.; Lee, J.; Heo, M.-S.; Yeo, I.-K.; Lee, N.-H.; Jee, Y.-H.; Kim, J.-S.; et al. Antioxidant Activities of Phlorotannins Purified from Ecklonia Cava on Free Radical Scavenging Using ESR and H2O2-Mediated DNA Damage. Eur. Food Res. Technol. 2007, 226, 71–79. [Google Scholar] [CrossRef]
- Kim, A.-R.; Shin, T.-S.; Lee, M.-S.; Park, J.-Y.; Park, K.-E.; Yoon, N.-Y.; Kim, J.-S.; Choi, J.-S.; Jang, B.-C.; Byun, D.-S.; et al. Isolation and Identification of Phlorotannins from Ecklonia Stolonifera with Antioxidant and Anti-Inflammatory Properties. J. Agric. Food Chem. 2009, 57, 3483–3489. [Google Scholar] [CrossRef]
- Jung, H.A.; Roy, A.; Jung, J.H.; Choi, J.S. Evaluation of the Inhibitory Effects of Eckol and Dieckol Isolated from Edible Brown Alga Eisenia Bicyclis on Human Monoamine Oxidases A and B. Arch. Pharm. Res. 2017, 40, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-A.; Lee, S.-H.; Lee, J.-H.; Kang, N.; Oh, J.-Y.; Seun-heui, S.; Ahn, G.; Ko, S.C.; Fernando, S.P.; Kim, S.-Y.; et al. A Marine Algal Polyphenol, Dieckol, Attenuates Blood Glucose Levels by Akt Pathway in Alloxan Induced Hyperglycemia Zebrafish Model. RSC Adv. 2016, 6, 78570–78575. [Google Scholar] [CrossRef]
- Ha, J.W.; Song, H.; Hong, S.S.; Boo, Y.C. Marine Alga Ecklonia Cava Extract and Dieckol Attenuate Prostaglandin E2 Production in HaCaT Keratinocytes Exposed to Airborne Particulate Matter. Antioxidants 2019, 8, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, G.; Andrade, P.; Valentão, P. Phlorotannins: Towards New Pharmacological Interventions for Diabetes Mellitus Type 2. Molecules 2016, 22, 56. [Google Scholar] [CrossRef] [Green Version]
- González-Colunga, D.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; Cruz-Suárez, L.E. Bioactivity-Guided Identification of Anti-AHPND (Acute Hepatopancreatic Necrosis Disease) Metabolites of Ecklonia Arborea. J. Appl. Phycol. 2019, 31, 3189–3199. [Google Scholar] [CrossRef]
- Kim, J.; Yoon, M.; Yang, H.; Jo, J.; Han, D.; Jeon, Y.-J.; Cho, S. Enrichment and Purification of Marine Polyphenol Phlorotannins Using Macroporous Adsorption Resins. Food Chem. 2014, 162, 135–142. [Google Scholar] [CrossRef]
- Catarino, M.; Silva, A.; Cardoso, S. Fucaceae: A Source of Bioactive Phlorotannins. Int. J. Mol. Sci. 2017, 18, 1327. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-C.; Kang, S.-M.; Ahn, G.; Kim, K.-N.; Kang, N.; Samarakoon, K.W.; Oh, M.-C.; Lee, J.-S.; Jeon, Y.-J. Protective Effect of a Marine Polyphenol, Dieckol against Carbon Tetrachloride-Induced Acute Liver Damage in Mouse. Environ. Toxicol. Pharmacol. 2013, 35, 517–523. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Kondo, S.; Segawa, S.; Lin, Y.-C.; Toyohara, H.; Ito, H.; Konoki, K.; Cho, Y.; Uchida, T. Isolation and Structural Determination of Two Novel Phlorotannins from the Brown Alga Ecklonia Kurome Okamura, and Their Radical Scavenging Activities. Mar. Drugs 2013, 11, 165–183. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Kim, S.-B.; Hwang, H.-J.; Kim, Y.-M.; Lee, M.-S. Antibacterial Property of Ecklonia Cava Extract against Marine Bacterial Pathogens. J. Food Hyg. Saf. 2016, 31, 380–385. [Google Scholar] [CrossRef]
- Munin, A.; Edwards-Lévy, F. Encapsulation of Natural Polyphenolic Compounds; a Review. Pharmaceutics 2011, 3, 793–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-X.; Wijesekara, I.; Li, Y.; Kim, S.-K. Phlorotannins as Bioactive Agents from Brown Algae. Process. Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Sallam, I.E.; Abdelwareth, A.; Attia, H.; Aziz, R.K.; Homsi, M.N.; von Bergen, M.; Farag, M.A. Effect of Gut Microbiota Biotransformation on Dietary Tannins and Human Health Implications. Microorganisms 2021, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.; Mu, T.; Sun, H.; Garcia-Vaquero, M. Phlorotannins: A Review of Extraction Methods, Structural Characteristics, Bioactivities, Bioavailability, and Future Trends. Algal Res. 2021, 60, 102484. [Google Scholar] [CrossRef]
- Grgić, J.; Šelo, G.; Planinić, M.; Tišma, M.; Bucić-Kojić, A. Role of the Encapsulation in Bioavailability of Phenolic Compounds. Antioxidants 2020, 9, 923. [Google Scholar] [CrossRef]
- Rahaiee, S.; Assadpour, E.; Faridi Esfanjani, A.; Silva, A.S.; Jafari, S.M. Application of Nano/Microencapsulated Phenolic Compounds against Cancer. Adv. Colloid Interface Sci. 2020, 279, 102153. [Google Scholar] [CrossRef]
- Wijesekara, I.; Yoon, N.Y.; Kim, S.-K. Phlorotannins from Ecklonia Cava (Phaeophyceae): Biological Activities and Potential Health Benefits. BioFactors 2010, 36, 408–414. [Google Scholar] [CrossRef]
- Cotas, J.; Pacheco, D.; Gonçalves, A.M.M.; Silva, P.; Carvalho, L.G.; Pereira, L. Seaweeds’ Nutraceutical and Biomedical Potential in Cancer Therapy: A Concise Review. J. Cancer Metastasis Treat. 2021, 2021. [Google Scholar] [CrossRef]
- Catarino, M.; Silva, A.; Mateus, N.; Cardoso, S. Optimization of Phlorotannins Extraction from Fucus Vesiculosus and Evaluation of Their Potential to Prevent Metabolic Disorders. Mar. Drugs 2019, 17, 162. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.P.; Hofseth, L.J.; Harris, C.C. Radical Causes of Cancer. Nat. Rev. Cancer 2003, 3, 276–285. [Google Scholar] [CrossRef]
- Sepantafar, M.; Maheronnaghsh, R.; Mohammadi, H.; Radmanesh, F.; Hasani-sadrabadi, M.M.; Ebrahimi, M.; Baharvand, H. Engineered Hydrogels in Cancer Therapy and Diagnosis. Trends Biotechnol. 2017, 35, 1074–1087. [Google Scholar] [CrossRef] [PubMed]
- Negara, B.F.S.P.; Sohn, J.H.; Kim, J.-S.; Choi, J.-S. Effects of Phlorotannins on Organisms: Focus on the Safety, Toxicity, and Availability of Phlorotannins. Foods 2021, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.-C.; Kim, S.H.; Park, Y.; Lee, B.H.; Hwang, H.J. Effects of 12-Week Oral Supplementation of Ecklonia Cava Polyphenols on Anthropometric and Blood Lipid Parameters in Overweight Korean Individuals: A Double-Blind Randomized Clinical Trial. Phyther. Res. 2012, 26, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Um, M.Y.; Kim, J.Y.; Han, J.K.; Kim, J.; Yang, H.; Yoon, M.; Kim, J.; Kang, S.W.; Cho, S. Phlorotannin Supplement Decreases Wake after Sleep Onset in Adults with Self-Reported Sleep Disturbance: A Randomized, Controlled, Double-Blind Clinical and Polysomnographic Study. Phyther. Res. 2018, 32, 698–704. [Google Scholar] [CrossRef]
- Baldrick, F.R.; McFadden, K.; Ibars, M.; Sung, C.; Moffatt, T.; Megarry, K.; Thomas, K.; Mitchell, P.; Wallace, J.M.W.; Pourshahidi, L.K.; et al. Impact of a (Poly)Phenol-Rich Extract from the Brown Algae Ascophyllum Nodosum on DNA Damage and Antioxidant Activity in an Overweight or Obese Population: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2018, 108, 688–700. [Google Scholar] [CrossRef]
- Paradis, M.-E.; Couture, P.; Lamarche, B. A Randomised Crossover Placebo-Controlled Trial Investigating the Effect of Brown Seaweed (Ascophyllum Nodosum and Fucus Vesiculosus) on Postchallenge Plasma Glucose and Insulin Levels in Men and Women. Appl. Physiol. Nutr. Metab. 2011, 36, 913–919. [Google Scholar] [CrossRef]
- Ko, S.-C.; Cha, S.-H.; Heo, S.-J.; Lee, S.-H.; Kang, S.-M.; Jeon, Y.-J. Protective Effect of Ecklonia Cava on UVB-Induced Oxidative Stress: In Vitro and in Vivo Zebrafish Model. J. Appl. Phycol. 2011, 23, 697–708. [Google Scholar] [CrossRef]
- Le, Q.-T.; Li, Y.; Qian, Z.-J.; Kim, M.-M.; Kim, S.-K. Inhibitory Effects of Polyphenols Isolated from Marine Alga Ecklonia Cava on Histamine Release. Process. Biochem. 2009, 44, 168–176. [Google Scholar] [CrossRef]
- Okeke, E.S.; Nweze, E.J.; Chibuogwu, C.C.; Anaduaka, E.G.; Chukwudozie, K.I.; Ezeorba, T.P.C. Aquatic Phlorotannins and Human Health: Bioavailability, Toxicity, and Future Prospects. Nat. Prod. Commun. 2021, 16. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Phlorotannin | Mechanism of Action | Anticancer Activity | Reference |
|---|---|---|---|
| Eckol | Interfere with Reg3A-mediated upregulation of JAK2, STAT3, NF-κB, cyclin D1 proteins | Antiproliferative action in pancreatic cancer cells | [12] |
| Eckol | - | Inhibitory activity against metastasis and reduced induced cell damages in cancer cell lines HeLa, H157 and MCF7 | [13] |
| Dioxinodehydroeckol | Reduced expression of Bcl-2 and NF-κB proteins | Antiproliferative activity in human breast cancer cells | [14] |
| Dieckol | Inhibition of TPA-induced matrix metalloproteinase-9 (MMP-9) activity in SK-Hep1 cells | Control and regulation of cancer cell motility | [15] |
| Dieckol | Interfere with Pi3K/AKT/mTOR signaling and caspases level; increased expression of E-cadherin | Inhibition and apoptosis of non-small–cell lung cancer cell line A549 | [16] |
| 6,6-bieckol | Increase the expression of E-cadherin and down-regulated Snail1 and Twist1 transcriptional levels—associated with lower survival rates in patients with cancer | Inhibition of non-small–cell lung cancer cells migration and proliferation | [20] |
| Dieckol, phlorofucofuroeckol | Decrease in the expression of receptor4 (TLR-4) and NF-κB promoter-driven transcriptional activity | Reduced proliferation, migration, tumor growth and inflammation | [21] |
| Dieckol | EMT marker protein expression and intracellular localization, cell motility, and cell invasion suppress hypoxia-induced EMT in HT29 cells via modulating cellular ROS and protein expression levels downstream of the HIF1 signaling pathway | Reduced cell motility of colorectal cancer cells | [22] |
| Dieckol | Inhibition of Pi3K/AKT/mTOR signaling and activation of the tumor suppressor protein E-cadherin | Inhibition of Non-small–cell lung cancer A549 migration | [16] |
| Dieckol | Increase antioxidants (SOD, CAT, GPx, and GSH) while decreasing phase-I enzymes Cyt-p450 and Cyt-b5; reduce pro-inflammatory modulators such as IL-6, IL-1, and TNF-α | Inhibition of DMBA-induced skin cancer in mice | [23] |
| Dieckol | Inhibition of ROS-mediated Rac1 activation and reduce WAVE2 expression, which interacts with NADPH oxidase component p47phox | Reduction of B16 melanoma cell motility and blocked invasive migration | [24] |
| Dieckol | Inhibitor of MMP-2, -9 expressions by the downregulation of NF-κB pathway | Suppression of cell cancer invasion | [25] |
| Eckol | Modulation of caspase-3 and Caspase-9 expression; Bcl-2 and Bax; epidermal growth factor receptor | Apoptosis and antiproliferative activity | [26,27] |
| Dieckol | Regulation of Bid, Bim, BAK, caspases-3 -7 -8 -9, and cleaved poly (ADP-ribose) polymerase expression through the increase of permeability of mitochondrial membranes and the release of cytochrome c from mitochondria into the cytosol | Reduced cell viability in a dose-dependent way and apoptosis in tumoral Hep3B cells | [28] |
| Dieckol | Regulation of the expression of apoptotic proteins caspase-8, caspase-9, and caspase-3 and decreasing the expression of AKT and p38 | Apoptotic effect on A2780 and SKOV3 ovarian cancer cells | [29] |
| dieckol | Suppression of NDEA-initiated hepatocarcinogenesis by modulating xenobiotic-metabolizing enzymes | Apoptosis and inhibited proliferation, invasion and angiogenesis signaling | [30] |
| phlorofucofuroeckol A | High expression of transcription factor 3 (ATF3), associated with apoptosis via the ATF3-mediated pathway | Apoptosis in colorectal cancer | [31] |
| Dieckol | Suppression of AKT and p38 activity; overexpression of AKT and p38 | Apoptosis in SKOV3 cells | [29] |
| Eckol | Action on PI3K/AKT and RAF-1/ERK signaling pathways, which regulate the maintenance of CSCs | Inhibition of proliferation, reduction in self-renewal and anchorage-independent growth ability of glioma CSCs | [18] |
| Eckol | Blockage of both Pi3K/AKT and Ras/Raf-1/Erk signaling pathways | Suppression of expression of glioma cell markers avoiding cell death | [17] |
| Phlorotannin | Mechanism of Action | Activity | Reference |
|---|---|---|---|
| Eckol | Revival of hematopoietic ability from the splenic progenitor cells | Photoprotective action in irradiation-damaged lymphocytes | [60] |
| Eckol | Regulation of pro-apoptotic p53 and Bax proteins; increased expression of antiapoptotic Bcl-2; increased height of jejunal villi and improvement of the jejunal crypt survival | Photoprotective action in intestinal stem cells for radiation-induced apoptosis | [61,62] |
| Eckol | Reduction of p53 and Bax proteins expression due to induction of Bcl-2 | Photoprotective action to peripheral cells; increased viability of lymphocytes; inhibition of apoptotic events; DNA repair | [63] |
| Eckol | Increase in Bcl-2 expression and reduction in Bax expression; suppression of SEK1-JNK-AP-1 and caspase-dependent pathway via mitochondria | Decrease in lipid peroxidation and apoptosis-induction; decrease in DNA fragmentation | [61] |
| Dieckol | Scavenging activity on free radicals | Reduction in γ radiation-induced toxicity and enhanced antioxidant activity; decrease in DNA damage and inflammation in hepatocyte cells | [64] |
| Dieckol | Scavenging activity on free radicals | Apoptosis of lung fibroblast cells by inhibiting ROS generation | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, P.; Lomartire, S.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Call the Eckols: Present and Future Potential Cancer Therapies. Mar. Drugs 2022, 20, 387. https://doi.org/10.3390/md20060387
Monteiro P, Lomartire S, Cotas J, Marques JC, Pereira L, Gonçalves AMM. Call the Eckols: Present and Future Potential Cancer Therapies. Marine Drugs. 2022; 20(6):387. https://doi.org/10.3390/md20060387
Chicago/Turabian StyleMonteiro, Pedro, Silvia Lomartire, João Cotas, João C. Marques, Leonel Pereira, and Ana M. M. Gonçalves. 2022. "Call the Eckols: Present and Future Potential Cancer Therapies" Marine Drugs 20, no. 6: 387. https://doi.org/10.3390/md20060387