
Bone Morphogenetic Protein-9 Promotes Osteogenic Differentiation and Mineralization in Human Stem-Cell-Derived Spheroids


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Current Study’s Design Involved GMSCs
2.2. Producing Stem Cell Spheroids
2.3. Evaluation of Cell Vitality
2.4. Alkaline Phosphatase Activity Levels and Calcium Deposits
2.5. Real-Time qPCR Measurement of mRNA for RUNX2 and COL1A1
2.6. Statistic Assessment
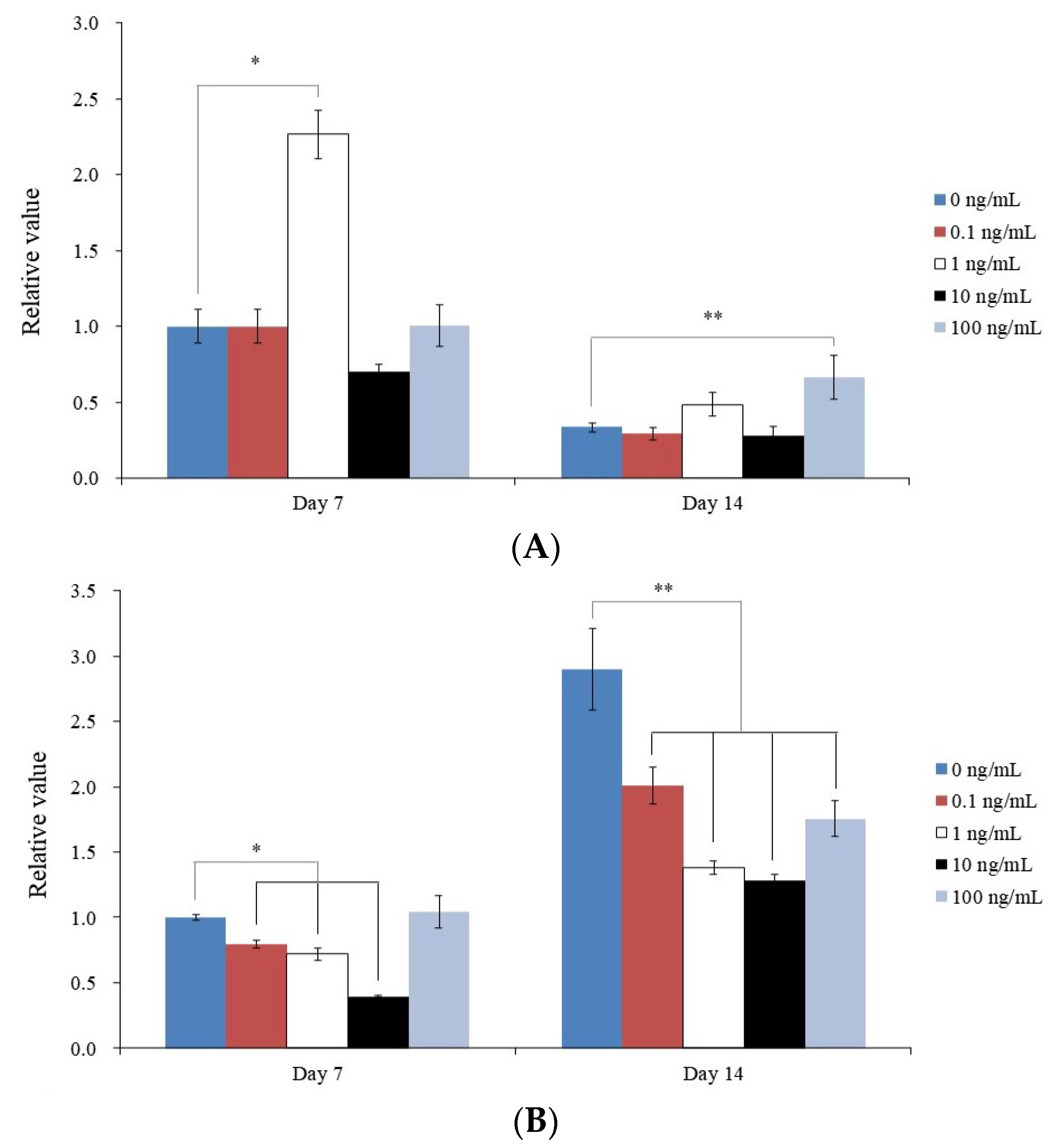
3. Results
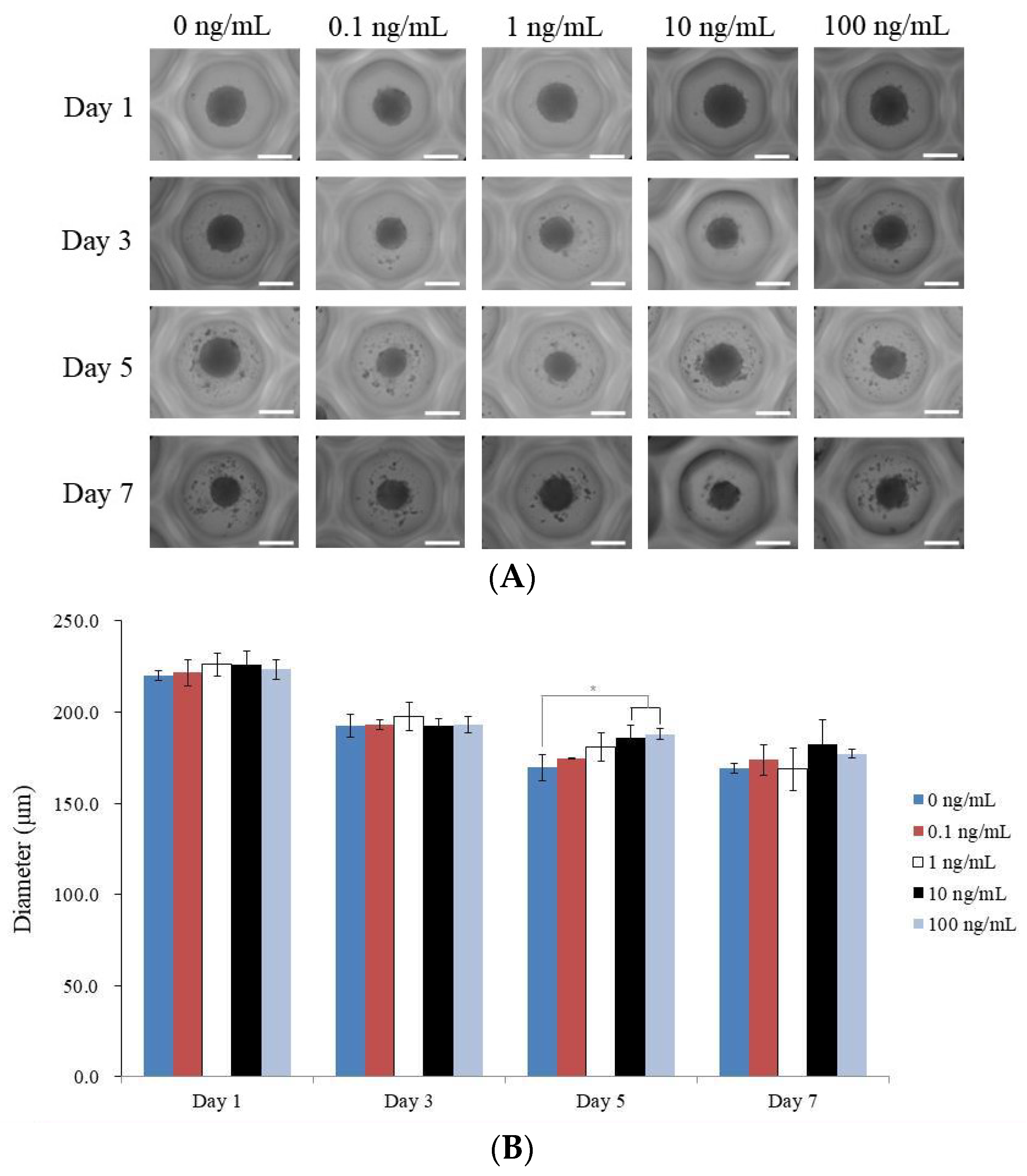
3.1. Human GMSC Spheroids
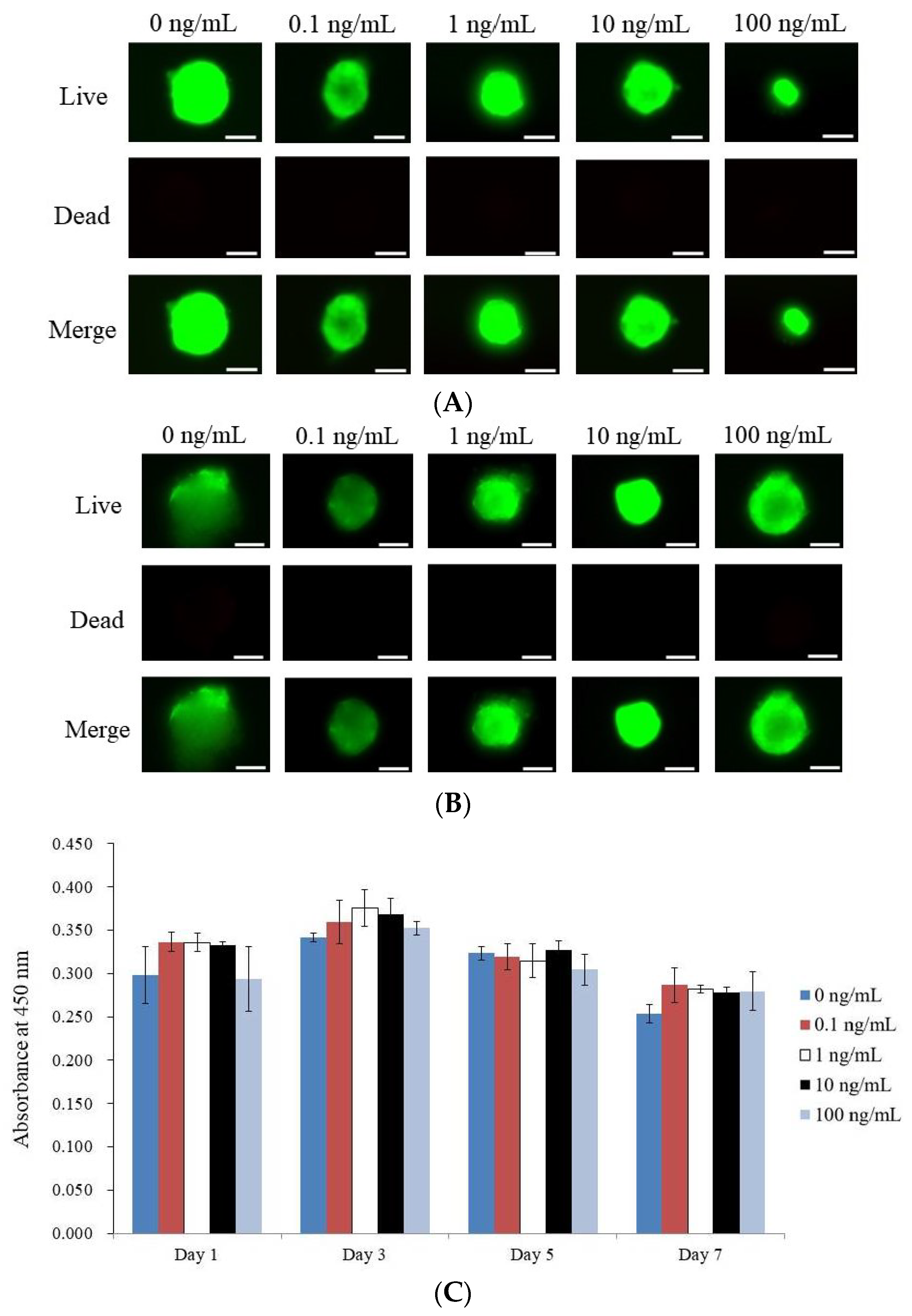
3.2. Determining the Vitality of Cell Spheroids Qualitatively and Quantitative Values for Cell Spheroid Viability
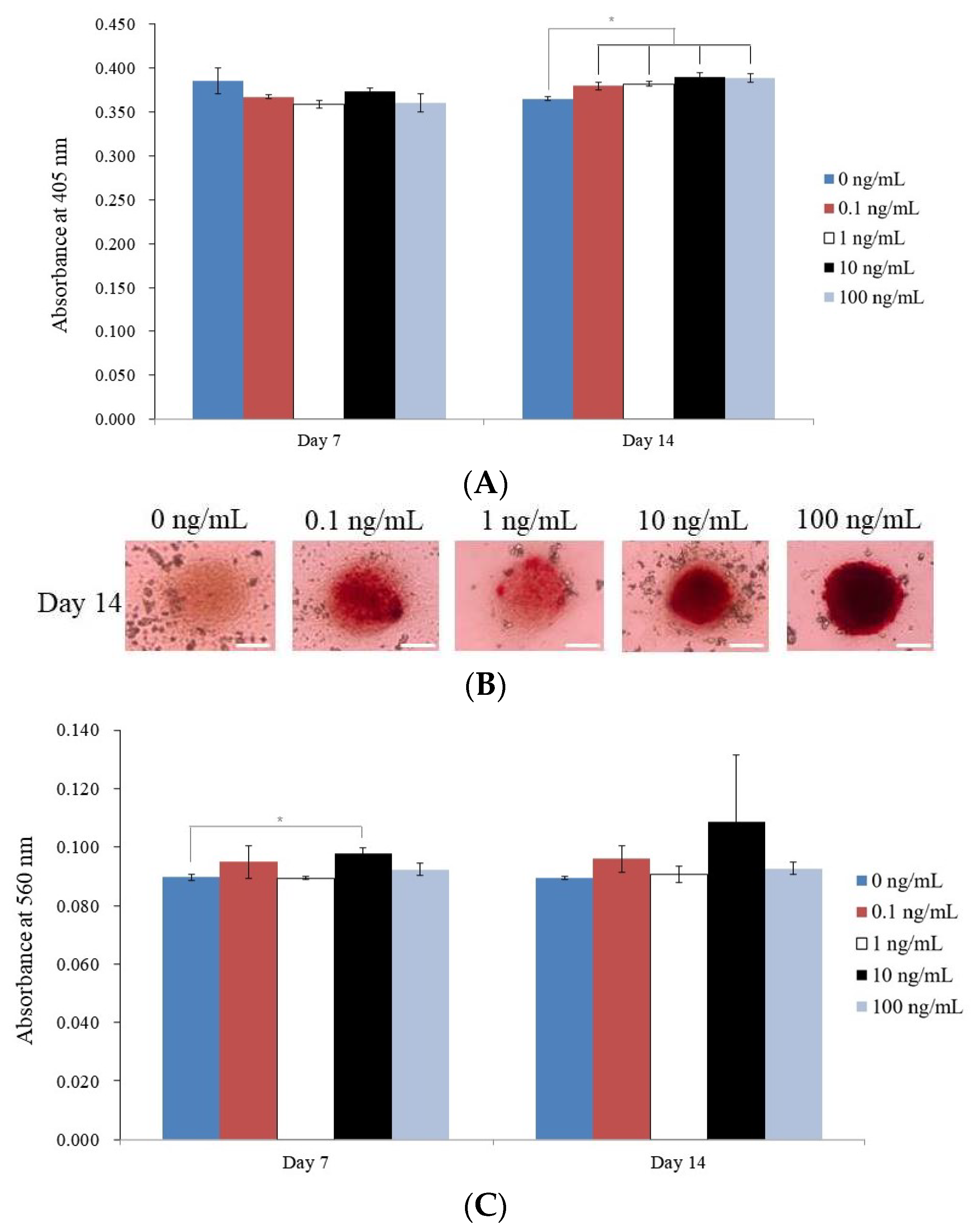
3.3. Evaluating Alkaline Phosphatase Activity and Alizarin Red S Staining
3.4. Analysis of RUNX2 and COL1A1 by qPCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, J.; Li, L. BMP signaling and stem cell regulation. Dev. Biol. 2005, 284, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lamplot, J.D.; Qin, J.; Nan, G.; Wang, J.; Liu, X.; Yin, L.; Tomal, J.; Li, R.; Shui, W.; Zhang, H.; et al. BMP9 signaling in stem cell differentiation and osteogenesis. Am. J. Stem Cells 2013, 2, 1–21. [Google Scholar]
- Wang, R.N.; Green, J.; Wang, Z.; Deng, Y.; Qiao, M.; Peabody, M.; Zhang, Q.; Ye, J.; Yan, Z.; Denduluri, S.; et al. Bone Morphogenetic Protein (BMP) signaling in development and human diseases. Genes Dis. 2014, 1, 87–105. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Mallet, C.; Keramidas, M.; Lamandé, N.; Gasc, J.-M.; Dupuis-Girod, S.; Plauchu, H.; Feige, J.-J.; Bailly, S. Bone morphogenetic protein-9 is a circulating vascular quiescence factor. Circ. Res. 2008, 102, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Levet, S.; Ciais, D.; Merdzhanova, G.; Mallet, C.; Zimmers, T.A.; Lee, S.-J.; Navarro, F.P.; Texier, I.; Feige, J.-J.; Bailly, S. Bone morphogenetic protein 9 (BMP9) controls lymphatic vessel maturation and valve formation. Blood J. Am. Soc. Hematol. 2013, 122, 598–607. [Google Scholar] [CrossRef] [Green Version]
- Luther, G.; Wagner, E.R.; Zhu, G.; Kang, Q.; Luo, Q.; Lamplot, J.; Bi, Y.; Luo, X.; Luo, J.; Teven, C. BMP-9 induced osteogenic differentiation of mesenchymal stem cells: Molecular mechanism and therapeutic potential. Curr. Gene Ther. 2011, 11, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Luu, H.H.; Song, W.X.; Luo, X.; Manning, D.; Luo, J.; Deng, Z.L.; Sharff, K.A.; Montag, A.G.; Haydon, R.C.; He, T.C. Distinct roles of bone morphogenetic proteins in osteogenic differentiation of mesenchymal stem cells. J. Orthop. Res. 2007, 25, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Song, W.X.; Luo, J.; Luo, X.; Chen, J.; Sharff, K.A.; Bi, Y.; He, B.C.; Huang, J.Y.; Zhu, G.H. BMP-9-induced osteogenic differentiation of mesenchymal progenitors requires functional canonical Wnt/β-catenin signalling. J. Cell. Mol. Med. 2009, 13, 2448–2464. [Google Scholar] [CrossRef] [Green Version]
- Hu, N.; Jiang, D.; Huang, E.; Liu, X.; Li, R.; Liang, X.; Kim, S.H.; Chen, X.; Gao, J.-L.; Zhang, H. BMP9-regulated angiogenic signaling plays an important role in the osteogenic differentiation of mesenchymal progenitor cells. J. Cell Sci. 2013, 126, 532–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, E.; Zhu, G.; Jiang, W.; Yang, K.; Gao, Y.; Luo, Q.; Gao, J.L.; Kim, S.H.; Liu, X.; Li, M. Growth hormone synergizes with BMP9 in osteogenic differentiation by activating the JAK/STAT/IGF1 pathway in murine multilineage cells. J. Bone Miner. Res. 2012, 27, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Kang, Q.; Luo, Q.; Jiang, W.; Si, W.; Liu, B.A.; Luu, H.H.; Park, J.K.; Li, X.; Luo, J. Inhibitor of DNA binding/differentiation helix-loop-helix proteins mediate bone morphogenetic protein-induced osteoblast differentiation of mesenchymal stem cells. J. Biol. Chem. 2004, 279, 32941–32949. [Google Scholar] [CrossRef] [Green Version]
- López-Coviella, I.; Berse, B.; Krauss, R.; Thies, R.S.; Blusztajn, J.K. Induction and maintenance of the neuronal cholinergic phenotype in the central nervous system by BMP-9. Science 2000, 289, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Zhou, W.; Dong, J.; Li, S.; Lv, P.; Liu, C. Scaffolds of bioactive glass (Bioglass®) combined with recombinant human bone morphogenetic protein -9 (rhBMP-9) for tooth extraction site preservation. Heliyon 2022, 8, e08796. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Yang, X.; Duan, L.; Huang, E.; Pengfei, Z.; Luo, W.; Zhang, Y.; Zeng, X.; Qiu, Y.; Cai, T. The healing of alveolar bone defects with novel bio-implants composed of Ad-BMP9-transfected rDFCs and CHA scaffolds. Sci. Rep. 2017, 7, 6373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Luo, W.; Zheng, L.; Hu, J.; Nie, L.; Zeng, H.; Tan, X.; Jiang, Y.; Li, Y.; Zhao, T.; et al. Efficient bone regeneration of BMP9-stimulated human periodontal ligament stem cells (hPDLSCs) in decellularized bone matrix (DBM) constructs to model maxillofacial intrabony defect repair. Stem Cell Res. Ther. 2022, 13, 535. [Google Scholar] [CrossRef]
- Jeong, G.S.; Song, J.H.; Kang, A.R.; Jun, Y.; Kim, J.H.; Chang, J.Y.; Lee, S.H. Surface tension-mediated, concave-microwell arrays for large-scale, simultaneous production of homogeneously sized embryoid bodies. Adv. Healthc. Mater. 2013, 2, 119–125. [Google Scholar] [CrossRef]
- Lee, G.H.; Park, Y.E.; Cho, M.; Park, H.; Park, J.Y. Magnetic force-assisted self-locking metallic bead array for fabrication of diverse concave microwell geometries. Lab Chip 2016, 16, 3565–3575. [Google Scholar] [CrossRef]
- Cesarz, Z.; Tamama, K. Spheroid Culture of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 9176357. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, Y.; Syková, E.; Kubinová, Š. The therapeutic potential of three-dimensional multipotent mesenchymal stromal cell spheroids. Stem Cell Res. Ther. 2017, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Ryu, N.E.; Lee, S.H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8, 1620. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.I.; Ko, Y.; Park, J.B. Evaluation of the shape, viability, stemness and osteogenic differentiation of cell spheroids formed from human gingiva-derived stem cells and osteoprecursor cells. Exp. Ther. Med. 2017, 13, 3467–3473. [Google Scholar] [CrossRef] [PubMed]
- Raik, S.; Thakur, R.; Rattan, V.; Kumar, N.; Pal, A.; Bhattacharyya, S. Temporal Modulation of DNA Methylation and Gene Expression in Monolayer and 3D Spheroids of Dental Pulp Stem Cells during Osteogenic Differentiation: A Comparative Study. Tissue Eng. Regen. Med. 2022, 19, 1267–1282. [Google Scholar] [CrossRef] [PubMed]
- Chao, X.; Zhao, F.; Hu, J.; Yu, Y.; Xie, R.; Zhong, J.; Huang, M.; Zeng, T.; Yang, H.; Luo, D.; et al. Comparative Study of Two Common In Vitro Models for the Pancreatic Islet with MIN6. Tissue Eng. Regen. Med. 2023, 20, 127–141. [Google Scholar] [CrossRef]
- Hu, T.; Zhang, H.; Yu, W.; Yu, X.; Li, Z.; He, L. The Combination of Concentrated Growth Factor and Adipose-Derived Stem Cell Sheet Repairs Skull Defects in Rats. Tissue Eng. Regen. Med. 2021, 18, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, A.E.; Xu, Q.; Zhang, Q.; Le, A.D. Gingiva-Derived Mesenchymal Stem Cells: Potential Application in Tissue Engineering and Regenerative Medicine—A Comprehensive Review. Front. Immunol. 2021, 12, 667221. [Google Scholar] [CrossRef]
- Lozano-Guillén, A.; López-García, S.; Rodríguez-Lozano, F.J.; Sanz, J.L.; Lozano, A.; Llena, C.; Forner, L. Comparative cytocompatibility of the new calcium silicate-based cement NeoPutty versus NeoMTA Plus and MTA on human dental pulp cells: An in vitro study. Clin. Oral Investig. 2022, 26, 7219–7228. [Google Scholar] [CrossRef]
- Guerrero-Gironés, J.; López-García, S.; Pecci-Lloret, M.R.; Pecci-Lloret, M.P.; Rodríguez Lozano, F.J.; García-Bernal, D. In vitro biocompatibility testing of 3D printing and conventional resins for occlusal devices. J. Dent. 2022, 123, 104163. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Cheng, T.S.; Liao, H.J.; Chuang, M.H.; Chen, H.T.; Chen, C.H.; Zhang, K.L.; Chang, C.H.; Lin, P.C.; Huang, C.F. Mesenchymal Stem Cell Secreted-Extracellular Vesicles are Involved in Chondrocyte Production and Reduce Adipogenesis during Stem Cell Differentiation. Tissue Eng. Regen. Med. 2022, 19, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Górski, B. Gingiva as a new and the most accessible source of mesenchymal stem cells from the oral cavity to be used in regenerative therapies. Postep. Hig. Med. Dosw. 2016, 70, 858–871. [Google Scholar] [CrossRef]
- Fawzy El-Sayed, K.M.; Dörfer, C.E. Gingival Mesenchymal Stem/Progenitor Cells: A Unique Tissue Engineering Gem. Stem Cells Int. 2016, 2016, 7154327. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Wang, A.; Hu, G.; Bian, M.; Chen, L.; Zhao, Q.; Sun, W.; Wu, Y. Potential of an Aligned Porous Hydrogel Scaffold Combined with Periodontal Ligament Stem Cells or Gingival Mesenchymal Stem Cells to Promote Tissue Regeneration in Rat Periodontal Defects. ACS Biomater. Sci. Eng. 2023, 9, 1961–1975. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cao, Z. Gingiva-derived Mesenchymal Stem Cells and Their Potential Applications in Oral and Maxillofacial Diseases. Curr. Stem Cell Res. Ther. 2020, 15, 43–53. [Google Scholar] [CrossRef]
- Jin, S.H.; Lee, J.E.; Yun, J.H.; Kim, I.; Ko, Y.; Park, J.B. Isolation and characterization of human mesenchymal stem cells from gingival connective tissue. J. Periodontal Res. 2015, 50, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.H.; Park, J.B.; Kim, I.; Lee, W.; Kim, H. Assessment of stem cell viability in the initial healing period in rabbits with a cranial bone defect according to the type and form of scaffold. J. Periodontal Implant. Sci. 2019, 49, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.B.; Tae, J.Y.; Ko, Y.; Park, J.B. Lovastatin increases the proliferation and osteoblastic differentiation of human gingiva-derived stem cells in three-dimensional cultures. Exp. Ther. Med. 2019, 18, 3425–3430. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Son, J.; Min, S.K.; Na, C.B.; Yi, G.; Koo, H.; Park, J.B. A Study of the Effects of Doxorubicin-Containing Liposomes on Osteogenesis of 3D Stem Cell Spheroids Derived from Gingiva. Materials 2019, 12, 2693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, J.; Chen, L.; Fan, Y.; Long, J.; Liu, S. Osteogenic and Angiogenic Potency of VEGF165-Transfected Canine Bone Marrow Mesenchymal Cells Combined with Coral Hydroxyapatite in Vitro. Tissue Eng. Regen. Med. 2021, 18, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Elango, J. Proliferative and Osteogenic Supportive Effect of VEGF-Loaded Collagen-Chitosan Hydrogel System in Bone Marrow Derived Mesenchymal Stem Cells. Pharmaceutics 2023, 15, 1297. [Google Scholar] [CrossRef]
- Rodríguez-Lozano, F.J.; López-García, S.; García-Bernal, D.; Sanz, J.L.; Lozano, A.; Pecci-Lloret, M.P.; Melo, M.; López-Ginés, C.; Forner, L. Cytocompatibility and bioactive properties of the new dual-curing resin-modified calcium silicate-based material for vital pulp therapy. Clin. Oral Investig. 2021, 25, 5009–5024. [Google Scholar] [CrossRef]
- Lee, H.; Lee, H.; Na, C.B.; Park, J.B. The effects of simvastatin on cellular viability, stemness and osteogenic differentiation using 3-dimensional cultures of stem cells and osteoblast-like cells. Adv. Clin. Exp. Med. 2019, 28, 699–706. [Google Scholar] [CrossRef]
- Min, S.K.; Kim, M.; Park, J.B. Bone morphogenetic protein 2-enhanced osteogenic differentiation of stem cell spheres by regulation of Runx2 expression. Exp. Ther. Med. 2020, 20, 79. [Google Scholar] [CrossRef]
- Son, J.; Tae, J.Y.; Min, S.K.; Ko, Y.; Park, J.B. Fibroblast growth factor-4 maintains cellular viability while enhancing osteogenic differentiation of stem cell spheroids in part by regulating RUNX2 and BGLAP expression. Exp. Ther. Med. 2020, 20, 2013–2020. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Weisgerber, D.W.; Bischoff, D.; Lewis, M.S.; Reid, R.R.; He, T.C.; Yamaguchi, D.T.; Miller, T.A.; Harley, B.A.; Lee, J.C. Nanoparticulate Mineralized Collagen Scaffolds and BMP-9 Induce a Long-Term Bone Cartilage Construct in Human Mesenchymal Stem Cells. Adv. Healthc. Mater. 2016, 5, 1821–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, T.; Liang, P.; Song, J.; Wang, J.; Zhou, P.; Tang, Y.; Li, J.; Huang, E. Matrigel Scaffolding Enhances BMP9-induced Bone Formation in Dental Follicle Stem/Precursor Cells. Int. J. Med. Sci. 2019, 16, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addante, A.; González-Corralejo, C.; Roncero, C.; Lazcanoiturburu, N.; García-Sáez, J.; Herrera, B.; Sánchez, A. BMP9 Promotes an Epithelial Phenotype and a Hepatocyte-like Gene Expression Profile in Adult Hepatic Progenitor Cells. Cells 2022, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Fujioka-Kobayashi, M.; Schaller, B.; Saulacic, N.; Pippenger, B.E.; Zhang, Y.; Miron, R.J. Absorbable collagen sponges loaded with recombinant bone morphogenetic protein 9 induces greater osteoblast differentiation when compared to bone morphogenetic protein 2. Clin. Exp. Dent. Res. 2017, 3, 32–40. [Google Scholar] [CrossRef]
- Zheng, W.; Chen, Q.; Zhang, Y.; Xia, R.; Gu, X.; Hao, Y.; Yu, Z.; Sun, X.; Hu, D. BMP9 promotes osteogenic differentiation of SMSCs by activating the JNK/Smad2/3 signaling pathway. J. Cell. Biochem. 2020, 121, 2851–2863. [Google Scholar] [CrossRef]
- Fujioka-Kobayashi, M.; Marjanowski, S.D.; Kono, M.; Hino, S.; Saulacic, N.; Schaller, B. Osteoinductive potential of recombinant BMP-9 in bone defects of mice treated with antiresorptive agents. Int. J. Oral Maxillofac. Surg. 2022, 51, 566–575. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Y.; Yang, J.; Huang, J.; Luo, C.; Zhang, J.; Yan, W.; Ao, Y. Bone morphogenetic protein 9 enhances osteogenic and angiogenic responses of human amniotic mesenchymal stem cells cocultured with umbilical vein endothelial cells through the PI3K/AKT/m-TOR signaling pathway. Aging 2021, 13, 24829–24849. [Google Scholar] [CrossRef]
- van Caam, A.; Blaney Davidson, E.; Garcia de Vinuesa, A.; van Geffen, E.; van den Berg, W.; Goumans, M.J.; ten Dijke, P.; van der Kraan, P. The high affinity ALK1-ligand BMP9 induces a hypertrophy-like state in chondrocytes that is antagonized by TGFβ1. Osteoarthr. Cartil. 2015, 23, 985–995. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Ma, Z.X.; Deng, R.; Jiang, H.T.; Chu, L.; Deng, Z.L. The SIRT1 activator SRT2104 promotes BMP9-induced osteogenic and angiogenic differentiation in mesenchymal stem cells. Mech. Ageing Dev. 2022, 207, 111724. [Google Scholar] [CrossRef]
- Dumont, R.J.; Dayoub, H.; Li, J.Z.; Dumont, A.S.; Kallmes, D.F.; Hankins, G.R.; Helm, G.A. Ex vivo bone morphogenetic protein-9 gene therapy using human mesenchymal stem cells induces spinal fusion in rodents. Neurosurgery 2002, 51, 1239–1244. [Google Scholar] [CrossRef]
- Zheng, Y.; Dong, X.; Wang, X.; Wang, J.; Chen, S.; He, Y.; An, J.; He, L.; Zhang, Y. Exosomes Derived from Adipose Tissue-Derived Mesenchymal Stromal Cells Prevent Medication-Related Osteonecrosis of the Jaw through IL-1RA. Int. J. Mol. Sci. 2023, 24, 8694. [Google Scholar] [CrossRef]
- Rodríguez-Lozano, F.J.; Oñate-Sánchez, R.; Gonzálvez-García, M.; Vallés-Bergadá, M.; Martínez, C.M.; Revilla-Nuin, B.; Guerrero-Gironés, J.; Moraleda, J.M.; García-Bernal, D. Allogeneic Bone Marrow Mesenchymal Stem Cell Transplantation in Tooth Extractions Sites Ameliorates the Incidence of Osteonecrotic Jaw-Like Lesions in Zoledronic Acid-Treated Rats. J. Clin. Med. 2020, 9, 1649. [Google Scholar] [CrossRef] [PubMed]
- Pryzhkova, M.V.; Boers, R.; Jordan, P.W. Modeling Human Gonad Development in Organoids. Tissue Eng. Regen. Med. 2022, 19, 1185–1206. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Yoon, H.S.; Lee, Y.; Kim, Y.H.; Cho, K.A.; Woo, S.Y.; Kim, H.S.; Ryu, K.H.; Park, J.W. Matrix Metalloproteinase 1 as a Marker of Tonsil-Derived Mesenchymal Stem Cells to Assess Bone Marrow Cell Migration. Tissue Eng. Regen. Med. 2022, 20, 271–284. [Google Scholar] [CrossRef]
- Han, Z.; He, X.; Feng, Y.; Jiang, W.; Zhou, N.; Huang, X. Hsp20 Promotes Endothelial Progenitor Cell Angiogenesis via Activation of PI3K/Akt Signaling Pathway under Hypoxia. Tissue Eng. Regen. Med. 2022, 19, 1251–1266. [Google Scholar] [CrossRef] [PubMed]
- Marrazzo, P.; Pizzuti, V.; Zia, S.; Sargenti, A.; Gazzola, D.; Roda, B.; Bonsi, L.; Alviano, F. Microfluidic Tools for Enhanced Characterization of Therapeutic Stem Cells and Prediction of Their Potential Antimicrobial Secretome. Antibiotics 2021, 10, 750. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-B.; Lee, H.-J.; Park, J.-B. Bone Morphogenetic Protein-9 Promotes Osteogenic Differentiation and Mineralization in Human Stem-Cell-Derived Spheroids. Medicina 2023, 59, 1315. https://doi.org/10.3390/medicina59071315
Lee S-B, Lee H-J, Park J-B. Bone Morphogenetic Protein-9 Promotes Osteogenic Differentiation and Mineralization in Human Stem-Cell-Derived Spheroids. Medicina. 2023; 59(7):1315. https://doi.org/10.3390/medicina59071315
Chicago/Turabian StyleLee, Sung-Bin, Hyun-Jin Lee, and Jun-Beom Park. 2023. "Bone Morphogenetic Protein-9 Promotes Osteogenic Differentiation and Mineralization in Human Stem-Cell-Derived Spheroids" Medicina 59, no. 7: 1315. https://doi.org/10.3390/medicina59071315