Leptin in the Commissural Nucleus of the Tractus Solitarius (cNTS) and Anoxic Stimulus in the Carotid Body Chemoreceptors Increases cNTS Leptin Signaling Receptor and Brain Glucose Retention in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Animals and General Surgery
2.3. Experimental Protocol
2.4. Materials and Drugs
2.5. Isolated Carotid Sinus Preparation, Carotid Chemoreceptors Stimulation and BGR Measurement
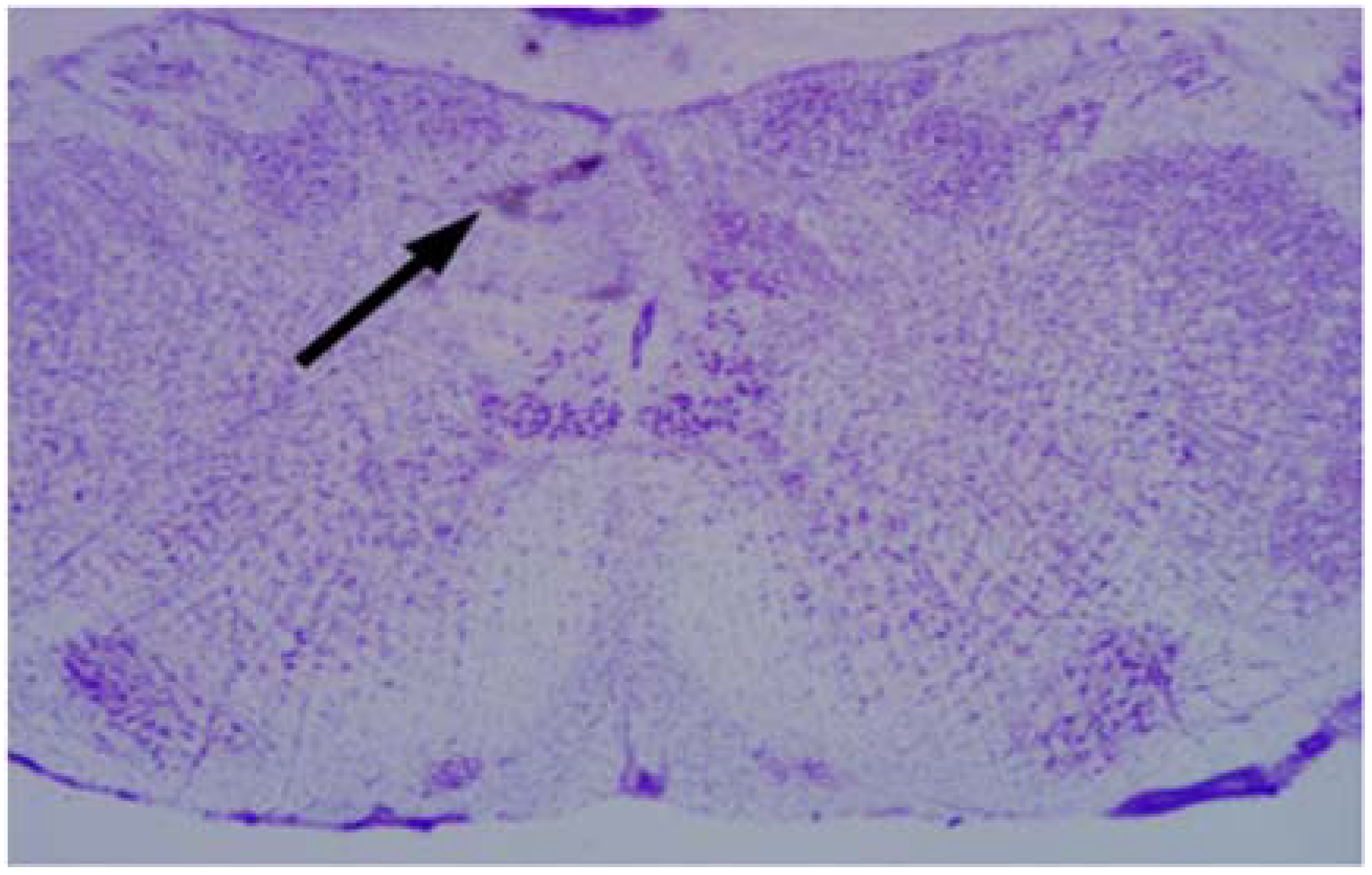
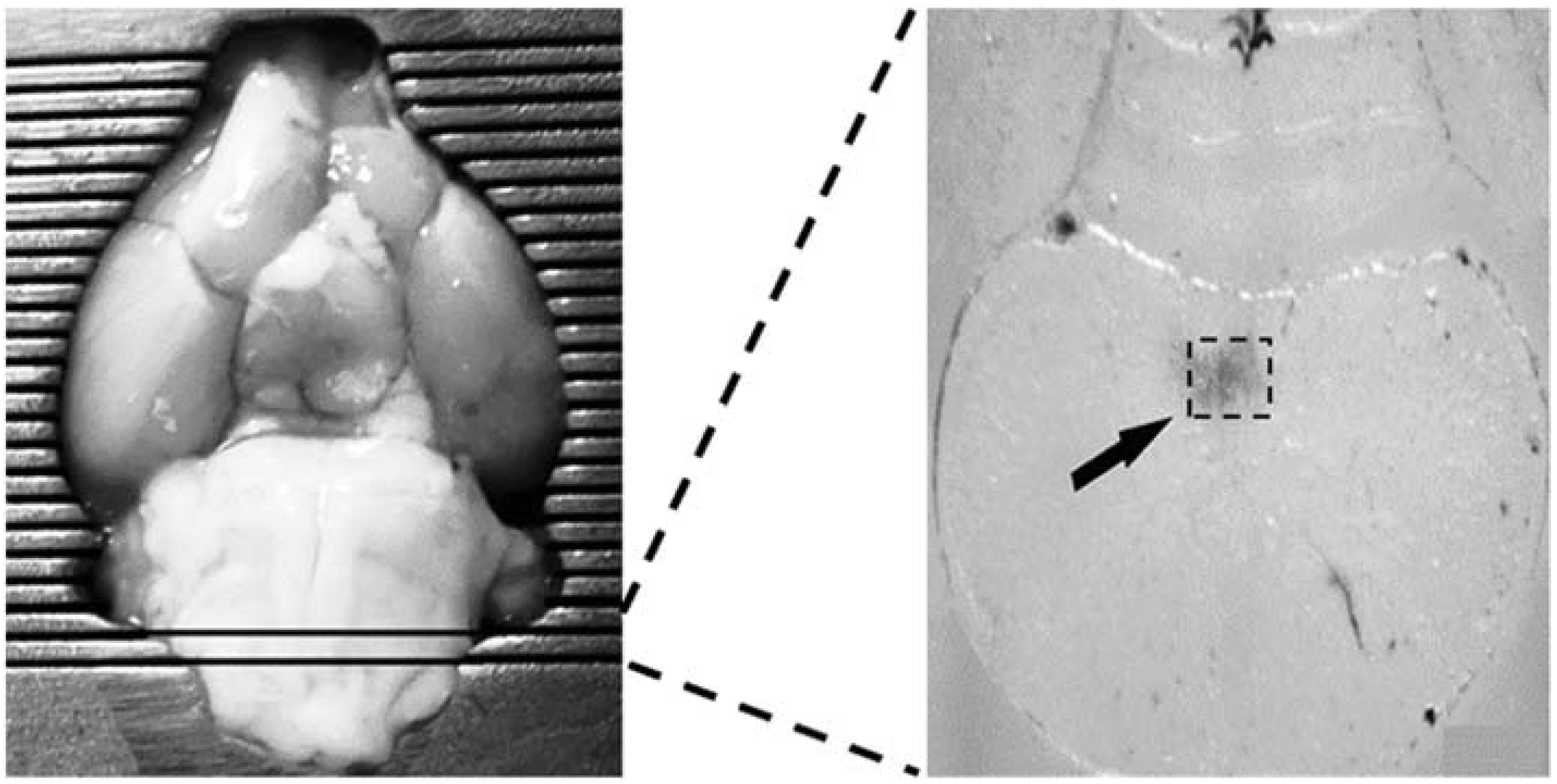
2.6. Microinjections of Drugs in the cNTS
2.7. Biochemical Assays
2.8. RT-PCR Analysis of Leptin and c-Fos mRNA Genes in the cNTS
2.9. Genes Expression Quantification
2.10. Data Analysis
3. Results
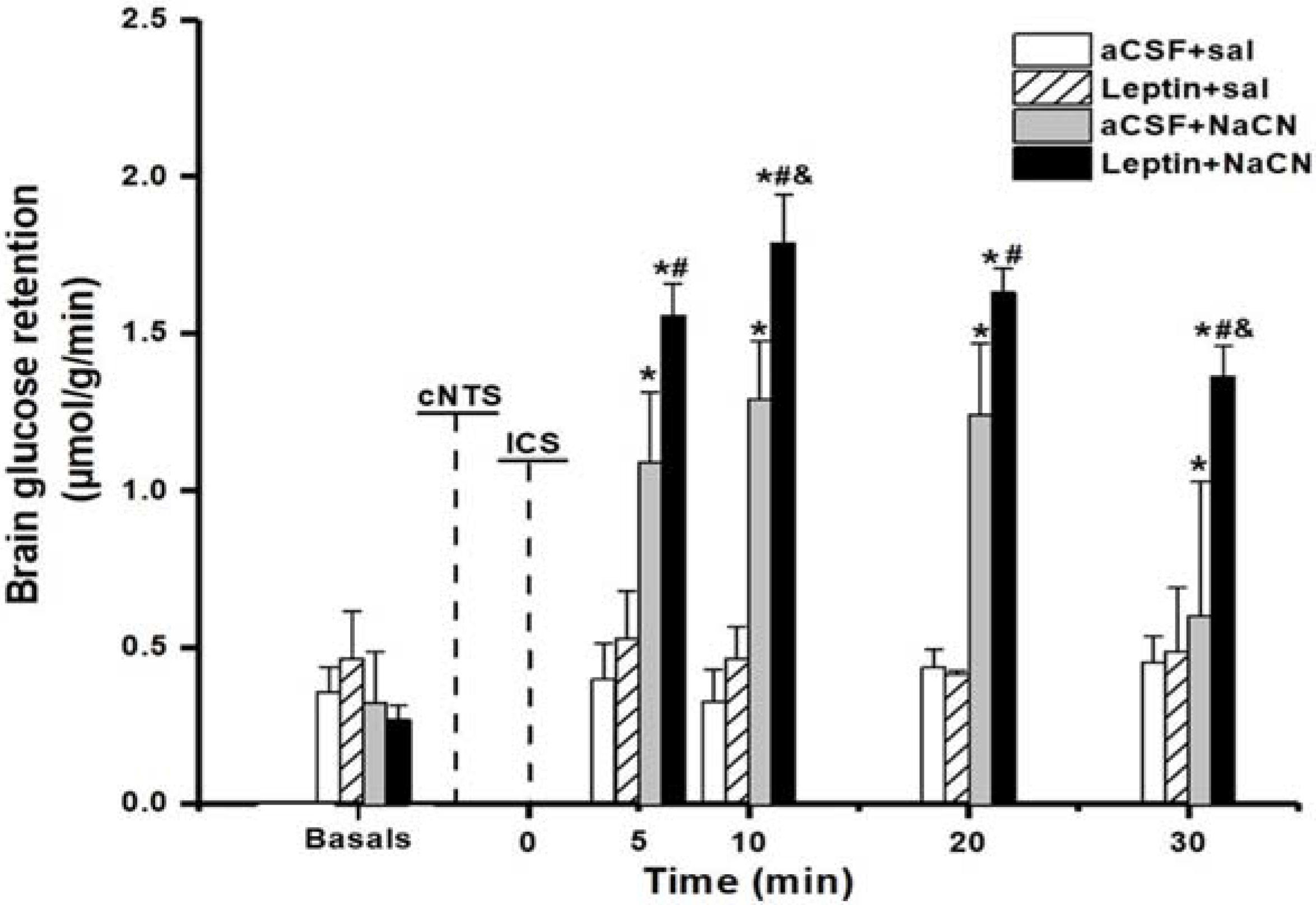
3.1. Brain Glucose Retention
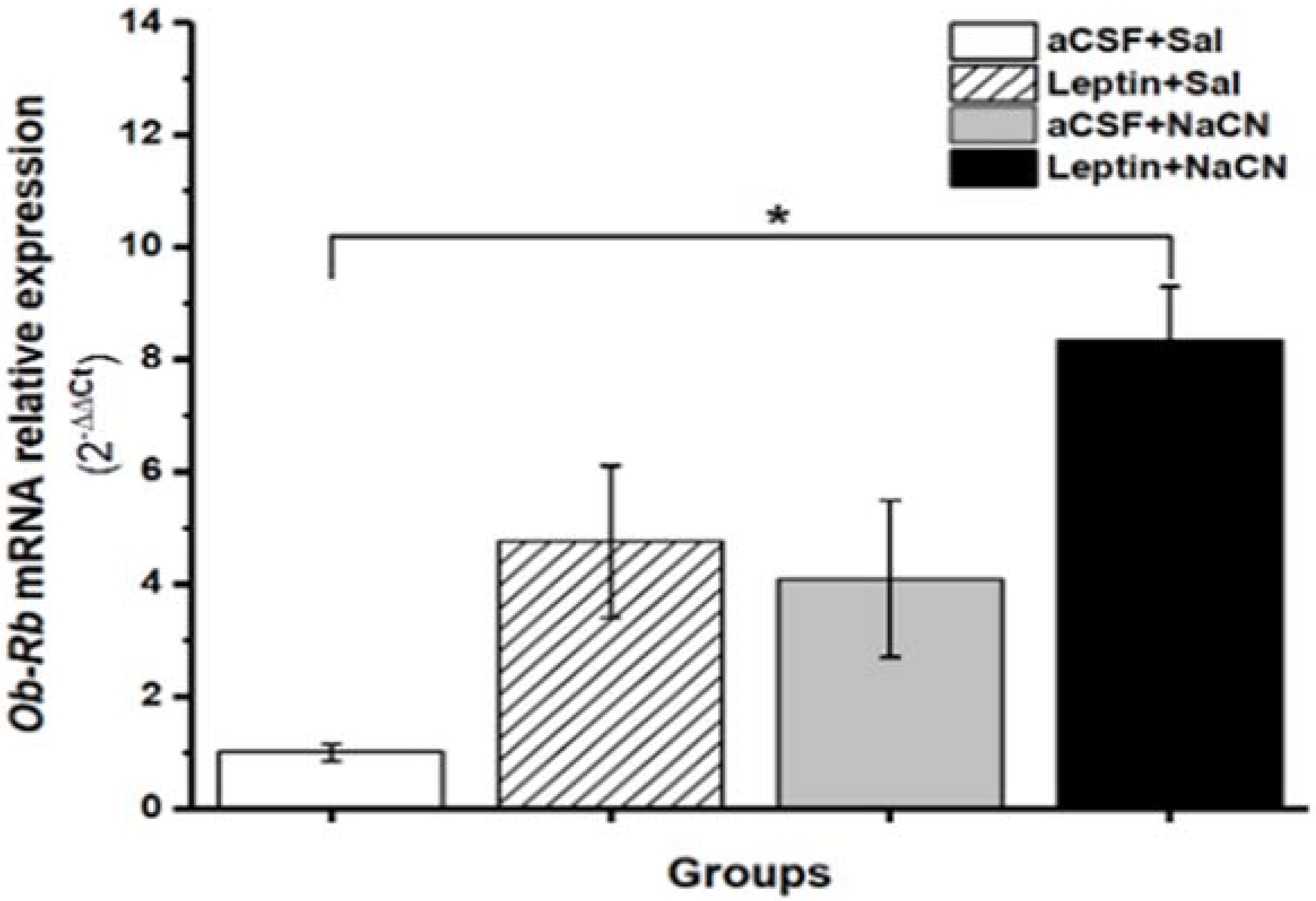
3.2. Leptin Ob-Rb mRNA Relative Expression in cNTS
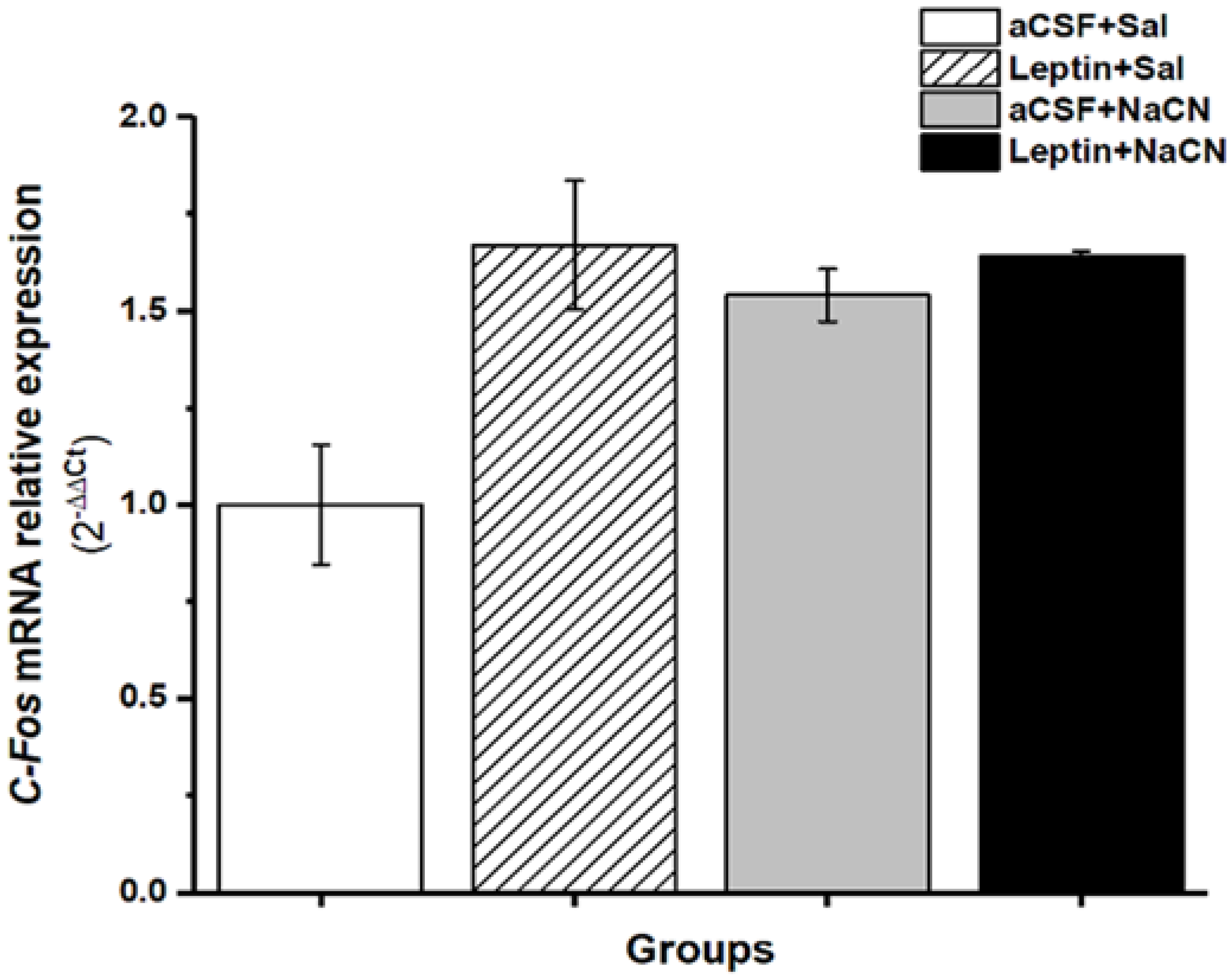
3.3. Relative Expression of c-Fos mRNA Gene Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sahu, A. Minireview: A hypothalamic role in energy balance with special emphasis on leptin. Endocrinology 2004, 145, 2613–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denroche, H.C.; Huynh, F.K.; Kieffer, T.J. The role of leptin in glucose homeostasis. J. Diabetes Investig. 2012, 3, 115–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, J.; Herzog, H. CART in the regulation of appetite and energy homeostasis. Front. Neurosci. 2014, 8, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, D.A.; Davis, S.N. Leptin: Metabolic control and regulation. J. Diabetes Complicat. 2003, 17, 108–113. [Google Scholar] [CrossRef]
- Oberlin, D.; Buettner, C. How does leptin restore euglycemia in insulin-deficient diabetes? J. Clin. Investig. 2017, 127, 450–453. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, A.M.; Neumann, U.H.; Glavas, M.M.; Kieffer, T.J. The glucoregulatory actions of leptin. Mol. Metab. 2017, 6, 1052–1065. [Google Scholar] [CrossRef]
- Hayes, M.R.; Skibicka, K.P.; Leichner, T.M.; Guarnieri, D.J.; DiLeone, R.J.; Bence, K.K.; Grill, H.J. Endogenous Leptin Signaling in the Caudal Nucleus Tractus Solitarius and Area Postrema Is Required for Energy Balance Regulation. Cell Metab. 2016, 23, 744. [Google Scholar] [CrossRef]
- Hosoi, T.; Kawagishi, T.; Okuma, Y.; Tanaka, J.; Nomura, Y. Brain stem is a direct target for leptin’s action in the central nervous system. Endocrinology 2002, 143, 3498–3504. [Google Scholar] [CrossRef] [Green Version]
- Ciriello, J.; Caverson, M.M. Carotid chemoreceptor afferent projections to leptin receptors containing neurons in the nucleus of the solitary tract. Peptides 2014, 58, 30–35. [Google Scholar] [CrossRef]
- Grill, H.J.; Schwartz, M.W.; Kaplan, J.M.; Foxhall, J.S.; Breininger, J.; Baskin, D.G. Evidence that the caudal brainstem is a target for the inhibitory effect of leptin on food intake. Endocrinology 2002, 143, 239–246. [Google Scholar] [CrossRef]
- Ciriello, J.; Moreau, J.M. Systemic administration of leptin potentiates the response of neurons in the nucleus of the solitary tract to chemoreceptor activation in the rat. Neuroscience 2013, 229, 88–99. [Google Scholar] [CrossRef]
- Chen, I.L.; Weber, J.T.; Yates, R.D. Synaptic connections of central carotid sinus afferents in the nucleus of the tractus solitarius of the rat. II. Connections with substance P-immunoreactive neurons. J. Neurocytol. 1994, 23, 313–322. [Google Scholar] [CrossRef]
- Alvarez-Buylla, R.; de Alvarez-Buylla, E. Carotid sinus receptors participate in glucose homeostasis. Resp. Physiol. 1988, 72, 347–360. [Google Scholar] [CrossRef]
- Cuéllar, R.; Montero, S.; Luquín, S.; García-Estrada, J.; Dobrovinskaya, O.; Melnikov, V.; Lemus, M.; de Álvarez-Buylla, E.R. Glutamatergic Receptor Activation in the Commissural Nucleus Tractus Solitarii (cNTS) Mediates Brain Glucose Retention (BGR) Response to Anoxic Carotid Chemoreceptor (CChr) Stimulation in Rats. Adv. Exp. Med. Biol. 2015, 860, 301–308. [Google Scholar]
- Gao, L.; Ortega-Sáenz, P.; García-Fernández, M.; González-Rodríguez, P.; Caballero-Eraso, C.; López-Barneo, J. Glucose sensing by carotid body glomus cells: Potential implications in disease. Front. Physiol. 2014, 5, 398. [Google Scholar] [CrossRef] [Green Version]
- Lemus, M.; Mojarro, C.; Montero, S.; Melnikov, V.; Ramírez-Flores, M.; de Álvarez-Buylla, E.R. Leptin in the Commissural Nucleus Tractus Solitarii Increases the Glucose Responses to Carotid Chemoreceptors Activation by Cyanide. Adv. Exp. Med. Biol. 2018, 1071, 143–149. [Google Scholar]
- Verberne, A.J.; Sartor, D.M. Rostral ventrolateral medulla neurons modulate glucose homeostasis in the rat. Am. J. Physiol. 2010, 299, E802–E807. [Google Scholar]
- Adachi, A.; Shimizu, N.; Oomura, Y.; Kobáshi, M. Convergence of hepatoportal glucose-sensitive afferent signals to glucose-sensitive units within the nucleus of the solitary tract. Neurosci. Lett. 1984, 46, 215–218. [Google Scholar] [CrossRef]
- Mizuno, Y.; Oomura, Y. Glucose responding neurons in the nucleus tractus solitarius of the rat: In vitro study. Brain Res. 1984, 307, 109–116. [Google Scholar] [CrossRef]
- Ciriello, J.; Moreau, J.M. Leptin signaling in the nucleus of the solitary tract alters the cardiovascular responses to activation of the chemoreceptor reflex. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R727–R736. [Google Scholar] [CrossRef] [Green Version]
- Association for Assessment and Accreditation of Laboratory Animal Care from National Institutes of Health Guidelines, USA. 2011. Available online: https://grants.nih.gov/grants/olaw/guide-for-the-care-and-use-of-laboratory-animals.pdf (accessed on 2 November 2020).
- Cowan, A.; Lewis, J.W.; MacFarlane, I.R. Agonist and antagonist properties of buprenorphine, a new antinociceptive agent. Br. J. Pharmacol. 1977, 60, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNay, E.C.; Sherwin, R.S. From artificial cerebro-spinal fluid (aCSF) to artificial extracellular fluid (aECF): Microdialysis perfusate composition effects in in vivo brain ECF glucose measurements. J. Neurosci. Methods 2004, 132, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: New York, NY, USA, 1986. [Google Scholar]
- Yarkov, A.V.; Montero, S.; Lemus, M.; Roces de Álvarez-Buylla, E.; Álvarez-Buylla, R. Arginine-vasopressin in nucleus of the tractus solitarius induces hyperglycemia and brain glucose retention. Brain Res. 2001, 902, 212–222. [Google Scholar] [CrossRef]
- N’Diaye, A.; Sevoz-Couche, C.; Nosjean, A.; Hamon, M.; Laguzzi, R. Stimulation of 5-HT2 receptors in the nucleus tractus solitarius enhances NMDA receptor-mediated reflex-evoked bradycardia responses in the rat. Auton. Neurosci. 2001, 92, 45–55. [Google Scholar] [CrossRef]
- Waynfort, H.B.; Flecknell, P.A. Experimental and Surgical Technique in the Rat, 2nd ed.; Academic Press: Cambridge, MA, USA, 1995; pp. 276–278. [Google Scholar]
- Lee, H.B.; Blaufox, M.D. Blood volume in the rat. J. Nucl. Med. 1985, 26, 72–76. [Google Scholar]
- Alvarez-Buylla, R.; Huberman, A.; Montero, S.; Lemus, M.; Valles, V.; de Alvarez-Buylla, E.R. Induction of brain glucose uptake by a factor secreted into cerebrospinal fluid. Brain Res. 2003, 994, 124–133. [Google Scholar] [CrossRef]
- Lemus, M.; Montero, S.A.; Leal, C.A.; Portilla-de Buen, E.; Luquín, S.; García-Estrada, J.; Melnikov, V.; de Álvarez-Buylla, E. Nitric oxide infused in the solitary tract nucleus blocks brain glucose retention induced by carotid chemoreceptor stimulation. Nitric Oxide 2011, 25, 387–395. [Google Scholar] [CrossRef]
- Lainez, N.M.; Coss, D. Obesity, neuroinflammation, and reproductive function. Endocrinology 2019, 160, 2719–2736. [Google Scholar] [CrossRef]
- Buyse, M.; Ovesjö, M.-L.; Goïot, H.; Guilmeau, S.; Péranzi, G.; Moizo, L.; Walker, F.; Lewin, M.J.M.; Meister, B.; Bado, A. Expression and regulation of leptin receptor proteins in afferent and efferent neurons of the vagus nerve. Eur. Neurosci. 2001, 14, 64–72. [Google Scholar] [CrossRef]
- Feng, X.; Shikama, Y.; Shichishima, T.; Noji, H.; Ikeda, K.; Ogawa, K.; Kimura, H.; Takeishi, Y.; Kimura, J. Impairment of FOS mRNA stabilization following translation arrest in granulocytes from myelodysplastic syndrome patients. PLoS ONE 2013, 8, e61107. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Huo, L.; Maeng, L.; Bjørbaek, C.; Grill, H.J. Leptin and the control of food intake: Neurons in the nucleus of the solitary tract are activated by both gastric distension and leptin. Endocrinology 2007, 148, 2189–2197. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.; Gamber, K.M.; Grill, H.J.; Bjørbaek, C. Divergent leptin signaling in proglucagon neurons of the nucleus of the solitary tract in mice and rats. Endocrinology 2008, 149, 492–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.L.; Baskin, D.G.; Schwartz, M.W. Hindbrain leptin receptor stimulation enhances the anorexic response to cholecystokinin. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R1238–R1246. [Google Scholar] [CrossRef] [Green Version]
- Inyushkin, A.N.; Inyushkina, E.M.; Merkulova, N.A. Respiratory responses to microinjections of leptin into the solitary tract nucleus. Neurosci. Behav. Physiol. 2009, 39, 231–240. [Google Scholar] [CrossRef]
- Harris, R.B.S. Low-dose infusions of leptin into the nucleus of the solitary tract increase sensitivity to third ventricle leptin. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E719–E728. [Google Scholar] [CrossRef]
- Hübschle, T.; Thom, E.; Watson, A.; Roth, J.; Klaus, S.; Meyerhof, W. Leptin-Induced Nuclear Translocation of STAT3 Immunoreactivity in Hypothalamic Nuclei Involved in Body Weight Regulation. J. Neurosci. 2001, 21, 2413–2424. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Karkanias, G.B.; Morales, J.C.; Hawkins, M.; Barzilai, N.; Wang, J.; Rossetti, L. Intracerebroventricular leptin regulates hepatic but not peripheral glucose fluxes. J. Biol. Chem. 1998, 273, 31160–31167. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Ahn, I.S.; Kim, D.S. Central infusion of leptin improves insulin resistance and suppresses beta-cell function, but not beta-cell mass, primarily through the sympathetic nervous system in a type 2 diabetic rat model. Life Sci. 2010, 86, 854–862. [Google Scholar] [CrossRef]
- Maroni, P.; Citterio, L.; Piccoletti, R.; Bendinelli, P. Sam68 and ERKs regulate leptin-induced expression of OB-Rb mRNA in C2C12 myotubes. Mol. Cell Endocrinol. 2009, 309, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Dallaporta, M.; Pecchi, E.; Pio, J.; Jean, A.; Horner, K.C.; Troadec, J.D. Expression of leptin receptor by glial cells of the nucleus tractus solitarius: Possible involvement in energy homeostasis. J. Neuroendocrinol. 2009, 21, 57–67. [Google Scholar] [CrossRef]
- Neyens, D.; Zhao, H.; Huston, N.J.; Wayman, G.A.; Ritter, R.C.; Appleyard, S.M. Leptin Sensitizes NTS Neurons to Vagal Input by Increasing Postsynaptic NMDA Receptor Currents. J. Neurosci. 2020, 40, 7054–7064. [Google Scholar] [CrossRef] [PubMed]
- Akieda-Asai, S.; Poleni, P.E.; Date, Y. Coinjection of CCK and leptin reduces food intake via increased CART/TRH and reduced AMPK phosphorylation in the hypothalamus. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1284–E1291. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemus, M.; Mojarro, C.; Montero, S.; Ramírez-Flores, M.; Torres-Magallanes, J.; Maturano-Melgoza, A.; Roces de Álvarez-Buylla, E. Leptin in the Commissural Nucleus of the Tractus Solitarius (cNTS) and Anoxic Stimulus in the Carotid Body Chemoreceptors Increases cNTS Leptin Signaling Receptor and Brain Glucose Retention in Rats. Medicina 2022, 58, 550. https://doi.org/10.3390/medicina58040550
Lemus M, Mojarro C, Montero S, Ramírez-Flores M, Torres-Magallanes J, Maturano-Melgoza A, Roces de Álvarez-Buylla E. Leptin in the Commissural Nucleus of the Tractus Solitarius (cNTS) and Anoxic Stimulus in the Carotid Body Chemoreceptors Increases cNTS Leptin Signaling Receptor and Brain Glucose Retention in Rats. Medicina. 2022; 58(4):550. https://doi.org/10.3390/medicina58040550
Chicago/Turabian StyleLemus, Mónica, Cynthia Mojarro, Sergio Montero, Mario Ramírez-Flores, José Torres-Magallanes, Adrián Maturano-Melgoza, and Elena Roces de Álvarez-Buylla. 2022. "Leptin in the Commissural Nucleus of the Tractus Solitarius (cNTS) and Anoxic Stimulus in the Carotid Body Chemoreceptors Increases cNTS Leptin Signaling Receptor and Brain Glucose Retention in Rats" Medicina 58, no. 4: 550. https://doi.org/10.3390/medicina58040550