IgE-Mediated Fish Allergy in Children

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Epidemiology
3. Pathogenesis and Clinical Features
3.1. Nonimmunological Adverse Reactions
- Anisakiasis: infection by the parasite Anisakis, resulting in mostly gastrointestinal clinical manifestations. It requires the ingestion of live parasites. Therefore it is only contracted after consuming raw, undercooked, or pickled fish [31].
- Scombroid poisoning: a syndrome caused by ingestion of poorly preserved fish (more often red meat fish like tuna), in which bacterial overgrowth allows histidine to be converted into histamine. Clinical manifestations mimic allergic reactions, with rapid onset (around 30 min after ingestion), for example, urticaria, oral allergic syndrome, nausea and vomiting, and, in rare cases, anaphylaxis. Patients, who often do not have a history of fish allergy, often report oral tingling sensation and metallic flavour when eating the responsible fish, and usually, the same signs and symptoms are reported by other family members who consumed the same food [32,33].
- Toxic algae poisoning: fish can consume several toxin-producing algae taken up by filter feeders such as mussels and clamps. Subsequently, the human ingestion of contaminated fish triggers this type of poisoning. The clinical manifestations are varied and depend on the toxin: e.g., Ciguatera, due to ciguatoxin found most commonly in tropical fishes (groupers, eel, Spanish mackerel), may present with cutaneous (urticaria), gastrointestinal (nausea, vomiting), neurological (blurred vision, paraesthesia, ataxia, seizures) and cardiovascular (bradycardia/tachycardia, hypotension/hypertension, conduction block) signs and symptoms [34,35].
- Bacterial/viral contamination: eating fish raised in or harvested from contaminated waters will result mostly in gastrointestinal clinical manifestations arising several hours after ingestion, often accompanied by fever [36].
- Seafood intolerance: due to vasoactive amines present in fish (histamine and tyramine), especially when canned or pickled, or fish autolysates [37]. Usually presents itself with a headache.
3.2. Immunological Adverse Reactions
- IgE-mediated adverse reactions to fish: the most common form of adverse reaction to fish, which involves the development of sensitisation, a type 2 T helper (Th2) response, and production of sIgEs against fish allergens. Its pathogenesis and clinical features are discussed in more detail below.
- Non-IgE-mediated adverse reactions to fish: they include Food Protein-Induced Enterocolitis Syndrome (FPIES) and Food Protein-Induced Allergic Proctocolitis (FPIAP), of which fish is a major causative agent [38,39,40], and eosinophilic esophagitis (EoE)/gastritis [41,42,43]. In the management of EoE, an empiric six-food elimination diet is generally recommended and includes the elimination of fish/shellfish along with milk, egg, wheat, nuts, and soy [41,42,43,44]. Of note, some clinicians advise a four-food elimination diet and allow taking nuts and fish.
- Immunological, IgE-mediated adverse reactions to parasite infested fish. An immunological, IgE-mediated adverse reaction to Anisakis could occur due to the sensitisation to the nematode’s proteins, which infests various fish species. The clinical presentation is indistinguishable from a fish allergy, but sIgEs are not directed towards fish protein but rather towards the parasite. Thus, SPT and sIgE will detect Anisakis sensitisation [45].
4. Fish Allergens and Cross-Reactivity
4.1. Parvalbumin
4.2. Enolase and Aldolase
4.3. Collagen
4.4. Other Allergens
4.5. Cross-Reactivity
4.6. Food Processing
5. Diagnosis
5.1. Clinical History
5.2. SPTs
5.3. sIgEs
5.4. OFCs
- Open OFC: the food is administered in its daily-consumed form.
- Single-blind placebo-controlled food challenge (SBPCFC): the patient is blinded to the tested food.
- DBPCFC: both patient and observing heath care staff are blinded to the tested food.
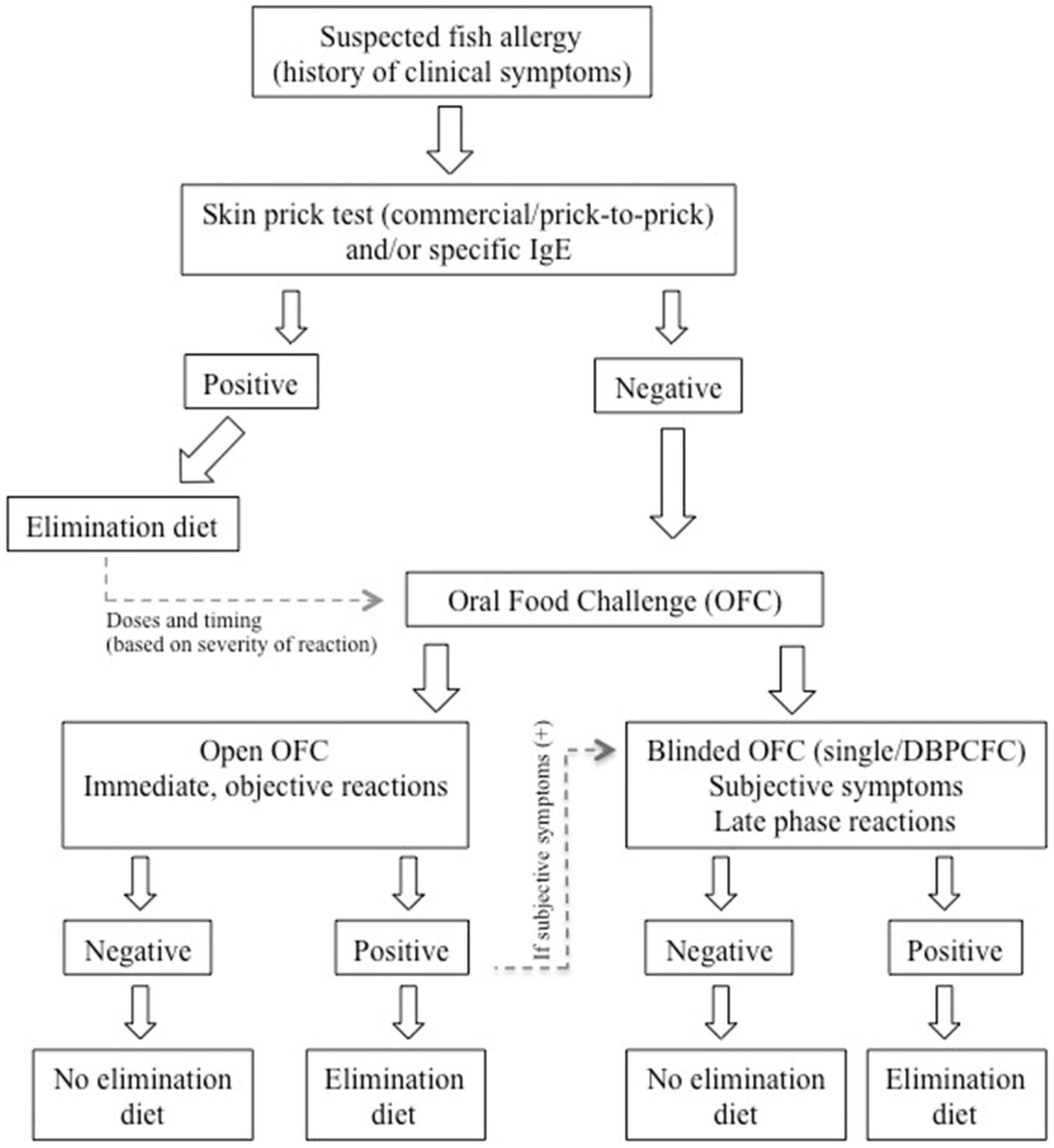
- In allergy practice, open food challenges are usually preferred, particularly in patients with clear history. However, awareness of the tested food by both the patient and the physician may bias false-positive results, particularly in patients with subjective symptoms such as abdominal pain. In these patients, DBPCFCs help confirm or rule out the disease. An algorithm for the diagnosis of fish allergy is indicated in Figure 2 [147].
5.5. Component Resolved Diagnosis (CRD)
5.6. Basophil Activation Test (BAT)
6. Management
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabenhenrich, L.; Trendelenburg, V.; Bellach, J.; Yürek, S.; Reich, A.; Fiandor, A.; Rivero, D.; Sigurdardottir, S.T.; Clausen, M.; Papadopoulos, N.G.; et al. Frequency of food allergy in school-aged children in eight European countries—The EuroPrevall-iFAAM birth cohort. Allergy 2020, 75, 2294–2308. [Google Scholar] [CrossRef]
- Adams, S.; Lopata, A.L.; Smuts, C.M.; Baatjies, R.; Jeebhay, M.F. Relationship between serum omega-3 fatty acid and asthma endpoints. Int. J. Environ. Res. Public Health 2019, 16, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moonesinghe, H.; Mackenzie, H.; Venter, C.; Kilburn, S.; Turner, P.; Weir, K.; Dean, T. Prevalence of fish and shellfish allergy: A systematic review. Ann. Allergy Asthma Immunol. 2016, 117, 264–272.e4. [Google Scholar] [CrossRef] [PubMed]
- Tsabouri, S.; Triga, M.; Makris, M.; Kalogeromitros, D.; Church, M.K.; Priftis, K.N. Fish and shellfish allergy in children: Review of a persistent food allergy. Pediatr. Allergy Immunol. 2012, 23, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Savage, J.; Sicherer, S.; Wood, R. The Natural History of Food Allergy. J. Allergy Clin. Immunol. Pract. 2016, 4, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Pyrhönen, K.; Näyhä, S.; Kaila, M.; Hiltunen, L.; Läärä, E. Occurrence of parent-reported food hypersensitivities and food allergies among children aged 1–4 years. Pediatr. Allergy Immunol. 2009, 20, 328–338. [Google Scholar] [CrossRef]
- Kajosaari, M. Food Allergy in Finnish Children Aged 1 to 6 Years. Acta Pediatr. 1982, 71, 815–819. [Google Scholar] [CrossRef]
- Dalal, I.; Binson, I.; Reifen, R.; Amitai, Z.; Shohat, T.; Rahmani, S.; Levine, A.; Ballin, A.; Somekh, E. Food allergy is a matter of geography after all: Sesame as a major cause of severe IgE-mediated food allergic reactions among infants and young children in Israel. Allergy Eur. J. Allergy Clin. Immunol. 2002, 57, 362–365. [Google Scholar] [CrossRef]
- Al-Hammadi, S.; Al-Maskari, F.; Bernsen, R. Prevalence of food allergy among children in Al-Ain City, United Arab Emirates. Int. Arch. Allergy Immunol. 2010, 151, 336–342. [Google Scholar] [CrossRef]
- Eggesbø, M.; Halvorsen, R.; Tambs, K.; Botten, G. Prevalence of parentally perceived adverse reactions to food in young children. Pediatr. Allergy Immunol. 1999, 10, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Pascual, C.Y.; Reche, M.; Fiandor, A.; Valbuena, T.; Cuevas, T.; Esteban, M.M. Fish allergy in childhood. Pediatr. Allergy Immunol. 2008, 19, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.F.; Pascual, C.; Burks, A.W.; Helm, R.M.; Esteban, M.M. Frequency of food allergy in a pediatric population from Spain. Pediatr. Allergy Immunol. 1995, 6, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Von Hertzen, L.; Mäkelä, M.J.; Petäys, T.; Jousilahti, P.; Kosunen, T.U.; Laatikainen, T.; Vartiainen, E.; Haahtela, T. Growing disparities in atopy between the Finns and the Russians: A comparison of 2 generations. J. Allergy Clin. Immunol. 2006, 117, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.; Venter, C.; Grundy, J.; Clayton, C.B.; Arshad, S.H.; Dean, T. Prevalence of sensitization to food allergens, reported adverse reaction to foods, food avoidance, and food hypersensitivity among teenagers. J. Allergy Clin. Immunol. 2005, 116, 884–892. [Google Scholar] [CrossRef]
- Pénard-Morand, C.; Raherison, C.; Kopferschmitt, C.; Caillaud, D.; Lavaud, F.; Charpin, D.; Bousquet, J.; Annesi-Maesano, I. Prevalence of food allergy and its relationship to asthma and allergic rhinitis in schoolchildren. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 1165–1171. [Google Scholar] [CrossRef]
- Osterballe, M.; Hansen, T.K.; Mortz, C.G.; Høst, A.; Bindslev-Jensen, C. The prevalence of food hypersensitivity in an unselected population of children and adults. Pediatr. Allergy Immunol. 2005, 16, 567–573. [Google Scholar] [CrossRef]
- Venter, C.; Pereira, B.; Grundy, J.; Clayton, C.B.; Arshad, S.H.; Dean, T. Prevalence of sensitization reported and objectively assessed food hypersensitivity amongst six-year-old children: A population-based study. Pediatr. Allergy Immunol. 2006, 17, 356–363. [Google Scholar] [CrossRef]
- Orhan, F.; Karakas, T.; Cakir, M.; Aksoy, A.; Baki, A.; Gedik, Y. Prevalence of immunoglobulin E-mediated food allergy in 6–9-year-old urban schoolchildren in the eastern Black Sea region of Turkey. Clin. Exp. Allergy 2009, 39, 1027–1035. [Google Scholar] [CrossRef]
- Kristinsdóttir, H.; Clausen, M.; Ragnarsdóttir, H.S.; Halldórsdóttir, I.H.; McBride, D.; Beyer, K.; Sigurdardóttir, S.T. Prevalence of food allergy in Icelandic infants during first year of life. Laeknabladid 2011, 97, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Roberts, G.; Muraro, A.; Sheikh, A. Prevalence of common food allergies in Europe: A systematic review and meta-analysis. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Thalayasingam, M.; Lee, B.W. Food allergy in Asia: How does it compare? Asia Pac. Allergy 2013, 3, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connett, G.J.; Gerez, I.; Cabrera-Morales, E.A.; Yuenyongviwat, A.; Ngamphaiboon, J.; Chatchatee, P.; Sangsupawanich, P.; Soh, S.E.; Yap, G.C.; Shek, L.P.C.; et al. A population-based study of fish allergy in the Philippines, Singapore and Thailand. Int. Arch. Allergy Immunol. 2012, 159, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Lao-Araya, M.; Trakultivakorn, M. Prevalence of food allergy among preschool children in northern Thailand. Pediatr. Int. 2012, 54, 238–243. [Google Scholar] [CrossRef]
- Le, T.T.K.; Nguyen, D.H.; Vu, A.T.L.; Ruethers, T.; Taki, A.C.; Lopata, A.L. A cross-sectional, population-based study on the prevalence of food allergies among children in two different socio-economic regions of Vietnam. Pediatr. Allergy Immunol. 2019, 30, 348–355. [Google Scholar] [CrossRef]
- Chen, J.; Hu, Y.; Allen, K.J.; Ho, M.H.K.; Li, H. The prevalence of food allergy in infants in Chongqing, China. Pediatr. Allergy Immunol. 2011, 22, 356–360. [Google Scholar] [CrossRef]
- Gupta, R.S.; Springston, E.E.; Warrier, M.R.; Smith, B.; Kumar, R.; Pongracic, J.; Holl, J.L. The prevalence, severity, and distribution of childhood food allergy in the United States. Pediatrics 2011, 128, e9–e17. [Google Scholar] [CrossRef] [Green Version]
- Mahdavinia, M.; Fox, S.R.; Smith, B.M.; James, C.; Palmisano, E.L.; Mohammed, A.; Zahid, Z.; Assa’ad, A.H.; Tobin, M.C.; Gupta, R.S. Racial Differences in Food Allergy Phenotype and Health Care Utilization among US Children. J. Allergy Clin. Immunol. Pract. 2017, 5, 352–357.e1. [Google Scholar] [CrossRef]
- Ben-Shoshan, M.; Harrington, D.W.; Soller, L.; Fragapane, J.; Joseph, L.; St Pierre, Y.; Godefroy, S.B.; Elliot, S.J.; Clarke, A.E. A population-based study on peanut, tree nut, fish, shellfish, and sesame allergy prevalence in Canada. J. Allergy Clin. Immunol. 2010, 125, 1327–1335. [Google Scholar] [CrossRef] [Green Version]
- Obeng, B.B.; Amoah, A.S.; Larbi, I.A.; Yazdanbakhsh, M.; Van Ree, R.; Boakye, D.A.; Hartgers, F.C. Food allergy in Ghanaian schoolchildren: Data on sensitization and reported food allergy. Int. Arch. Allergy Immunol. 2011, 155, 63–73. [Google Scholar] [CrossRef]
- Baird, F.J.; Gasser, R.B.; Jabbar, A.; Lopata, A.L. Foodborne anisakiasis and allergy. Mol. Cell. Probes 2014, 28, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guergué-Díaz de Cerio, O.; Barrutia-Borque, A.; Gardeazabal-García, J. Escombroidosis: Abordaje práctico. Actas Dermosifiliogr. 2016, 107, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Teuber, S.; Gershwin, M.E. Histamine (Scombroid) Fish Poisoning: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2016, 50, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Wjst, M. Ciguatera: A multifaceted and puzzling disorder. MMW—Fortschr. Med. 2016, 158, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Stephan, W.; Gribble, M.O.; Bienfang, P.; et al. An updated review of ciguatera fish poisoning: Clinical, epidemiological, environmental, and public health management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Johnston, E.B.; Nugraha, R.; Le, T.T.K.; Kalic, T.; McLean, T.R.; Kamath, S.D.; Lopata, A.L. Seafood allergy: A comprehensive review of fish and shellfish allergens. Mol. Immunol. 2018, 100, 28–57. [Google Scholar] [CrossRef] [PubMed]
- Biji, K.B.; Ravishankar, C.N.; Venkateswarlu, R.; Mohan, C.O.; Gopal, T.K.S. Biogenic amines in seafood: A review. J. Food Sci. Technol. 2016, 53, 2210–2218. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Wegrzyn, A.; Berin, M.C.; Mehr, S. Food Protein-Induced Enterocolitis Syndrome. J. Allergy Clin. Immunol. Pract. 2020, 8, 24–35. [Google Scholar] [CrossRef] [PubMed]
- González-Delgado, P.; Caparrós, E.; Moreno, M.V.; Clemente, F.; Flores, E.; Velásquez, L.; Rubio, G.; Fernández, J. Clinical and immunological characteristics of a pediatric population with food protein-induced enterocolitis syndrome (FPIES) to fish. Pediatr. Allergy Immunol. 2016, 27, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Miceli Sopo, S.; Monaco, S.; Badina, L.; Barni, S.; Longo, G.; Novembre, E.; Viola, S.; Monti, G. Food protein-induced enterocolitis syndrome caused by fish and/or shellfish in Italy. Pediatr. Allergy Immunol. 2015, 26, 731–736. [Google Scholar] [CrossRef]
- Patton, T.; Chugh, A.; Padhye, L.; DeGeeter, C.; Guandalini, S. Pediatric Celiac Disease and Eosinophilic Esophagitis: Outcome of Dietary Therapy. J. Pediatr. Gastroenterol. Nutr. 2019, 69, e43–e48. [Google Scholar] [CrossRef] [PubMed]
- Vinit, C.; Dieme, A.; Courbage, S.; Dehaine, C.; Dufeu, C.M.; Jacquemot, S.; Lajus, M.; Montigny, L.; Payen, E.; Yang, D.D.; et al. Eosinophilic esophagitis: Pathophysiology, diagnosis, and management. Arch. Pediatr. 2019, 26, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Sugimoto, M.; Tokaji, N.; Irahara, M.; Okamoto, K.; Uehara, H.; Kagami, S. Efficacy of an elimination diet in a patient with eosinophilic gastroenteritis: A pediatric case with multiple food allergies. J. Med. Investig. 2019, 66, 201–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xepapadaki, P.; Kitsioulis, N.A.; Manousakis, E.; Manolaraki, I.; Douladiris, N.; Papadopoulos, N.G. Remission Patterns of Food Protein-Induced Enterocolitis Syndrome in a Greek Pediatric Population. Int. Arch. Allergy Immunol. 2019, 180, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Tripodi, S.; Pingitore, G.; Calvani, M.; Scala, G.; Rodriguez-Perez, R.; Sfika, I.; Asero, R. Anisakis sensitivity in italian children: A prospective study. J. Investig. Allergol. Clin. Immunol. 2017, 27, 142–143. [Google Scholar] [CrossRef] [Green Version]
- McDole, J.R.; Wheeler, L.W.; McDonald, K.G.; Wang, B.; Konjufca, V.; Knoop, K.A.; Newberry, R.D.; Miller, M.J. Goblet cells deliver luminal antigen to CD103 + dendritic cells in the small intestine. Nature 2012, 483, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Niess, J.H.; Brand, S.; Gu, X.; Landsman, L.; Jung, S.; McCormick, B.A.; Vyas, J.M.; Boes, M.; Ploegh, H.L.; Fox, J.G.; et al. CX3CR1-mediated dendritic cell access to the intestinal lumen and bacterial clearance. Science 2005, 307, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Schülke, S. Induction of interleukin-10 producing dendritic cells as a tool to suppress allergen-specific T helper 2 responses. Front. Immunol. 2018, 9, 455. [Google Scholar] [CrossRef]
- Sheng, J.; Chen, W.; Zhu, H.J. The immune suppressive function of transforming growth factor-β (TGF-β) in human diseases. Growth Factors 2015, 33, 92–101. [Google Scholar] [CrossRef]
- Wambre, E.; Bajzik, V.; DeLong, J.H.; O’Brien, K.; Nguyen, Q.A.; Speake, C.; Gersuk, V.H.; DeBerg, H.A.; Whalen, E.; Ni, C.; et al. A phenotypically and functionally distinct human TH2 cell subpopulation is associated with allergic disorders. Sci. Transl. Med. 2017, 9, eaam9171. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.H.K.; Wong, W.H.S.; Chang, C. Clinical spectrum of food allergies: A comprehensive review. Clin. Rev. Allergy Immunol. 2014, 46, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Iweala, O.I.; Wesley Burks, A. Food allergy: Our evolving understanding of its pathogenesis, prevention, and treatment. Curr. Allergy Asthma Rep. 2016, 16, 37. [Google Scholar] [CrossRef] [PubMed]
- Pali-Schöll, I.; Untersmayr, E.; Klems, M.; Jensen-Jarolim, E. The effect of digestion and digestibility on allergenicity of food. Nutrients 2018, 10, 1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untersmayr, E.; Vestergaard, H.; Malling, H.J.; Jensen, L.B.; Platzer, M.H.; Boltz-Nitulescu, G.; Scheiner, O.; Skov, P.S.; Jensen-Jarolim, E.; Poulsen, L.K. Incomplete digestion of codfish represents a risk factor for anaphylaxis in patients with allergy. J. Allergy Clin. Immunol. 2007, 119, 711–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicherer, S.H.; Muñoz-Furlong, A.; Sampson, H.A. Prevalence of seafood allergy in the United States determined by a random telephone survey. J. Allergy Clin. Immunol. 2004, 114, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.A.; Regateiro, F.; Pereira, C.; Faria, E.; Pita, J.; Todo-Bom, A.; Carrapatoso, I. Anaphylaxis in a food allergy outpatient department: One-year review. Eur. Ann. Allergy Clin. Immunol. 2018, 50, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.H.; Lee, S.; Ahn, K.; Lee, S.Y.; Kim, K.W.; Kim, H.H.; Kim, J.H.; Yum, H.Y.; Kim, W.K.; Park, Y.M.; et al. Infantile anaphylaxis in Korea: A multicenter retrospective case study. J. Korean Med. Sci. 2019, 34, 1–12. [Google Scholar] [CrossRef]
- Sharp, M.F.; Lopata, A.L. Fish allergy: In review. Clin. Rev. Allergy Immunol. 2014, 46, 258–271. [Google Scholar] [CrossRef]
- Beaudouin, E.; Renaudin, J.M.; Morisset, M.; Codreanu, F.; Kanny, G.; Moneret-Vautrin, D.A. Food-dependent exercise-induced anaphylaxis—Update and current data. Eur. Ann. Allergy Clin. Immunol. 2006, 38, 45–51. [Google Scholar]
- Feldweg, A.M. Food-Dependent, Exercise-Induced Anaphylaxis: Diagnosis and Management in the Outpatient Setting. J. Allergy Clin. Immunol. Pract. 2017, 5, 283–288. [Google Scholar] [CrossRef]
- Foong, R.X.; Giovannini, M.; du Toit, G. Food-dependent exercise-induced anaphylaxis. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Kano, H.; Juji, F.; Shibuya, N.; Narita, M.; Naritaka, S.; Suko, M.; Morita, Y.; Iwata, T. Clinical courses of 18 cases with food-dependent exercise-induced anaphylaxis. Arerugi 2000, 49, 472–478. [Google Scholar]
- Wang, T.; Wu, K.; Li, L.Q.; Gong, L.L. A case of freshwater fish-dependent exercise-induced anaphylaxis. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 2019, 54, 469–470. [Google Scholar] [CrossRef]
- Lopata, A.L.; Jeebhay, M.F. Airborne seafood allergens as a cause of occupational allergy and asthma. Curr. Allergy Asthma Rep. 2013, 13, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Jeebhay, M.F.; Robins, T.G.; Lehrer, S.B.; Lopata, A.L. Occupational seafood allergy: A review. Occup. Environ. Med. 2001, 58, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Boulet, L.P.; Laberge, F. Occupational asthma to fish. Occup. Environ. Med. 2014, 71, 804. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, N.; Yagami, A.; Nakamura, M.; Nagai, A.; Matsunaga, K. Occupational fish allergy caused by percutaneous sensitization with α-actinin-3. Contact Dermat. 2017, 76, 322–323. [Google Scholar] [CrossRef] [PubMed]
- Yagami, A.; Suzuki, K.; Nakamura, M.; Sano, A.; Kobayashi, T.; Iwata, Y.; Arima, M.; Hara, K.; Matsunaga, K. Occupational food allergy due to parvalbumin and phaseolin induced by epicutaneous sensitization. Allergol. Int. 2015, 64, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Crespo, J.F.; Pascual, C.; Vallecillo, A.; Esteban, M.M. Sensitization to inhalant allergens in children diagnosed with food hypersensitivity. Allergy Proc. 1995, 16, 89–92. [Google Scholar] [CrossRef]
- James, J.M.; Crespo, J.F. Allergic reactions to foods by inhalation. Curr. Allergy Asthma Rep. 2007, 7, 167–174. [Google Scholar] [CrossRef]
- Beale, J.E.; Jeebhay, M.F.; Lopata, A.L. Characterisation of purified parvalbumin from five fish species and nucleotide sequencing of this major allergen from Pacific pilchard, Sardinops sagax. Mol. Immunol. 2009, 46, 2985–2993. [Google Scholar] [CrossRef] [PubMed]
- Van der Ventel, M.L.; Nieuwenhuizen, N.E.; Kirstein, F.; Hikuam, C.; Jeebhay, M.F.; Swoboda, I.; Brombacher, F.; Lopata, A.L. Differential responses to natural and recombinant allergens in a murine model of fish allergy. Mol. Immunol. 2011, 48, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, D.A.; Bahna, S.L. Food hypersensitivity by inhalation. Clin. Mol. Allergy 2009, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, S.; Pecoraro, R.; Filippelli, M.; Miraglia Del Giudice, M.; Marseglia, G.; Salpietro, C.; Arrigo, T.; Stringari, G.; Ricò, S.; La Rosa, M.; et al. Allergic reactions to foods by inhalation in children. Allergy Asthma Proc. 2014, 35, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Jeebhay, M.F.; Lopata, A.L. Occupational Allergies in Seafood-Processing Workers, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; Volume 66. [Google Scholar] [CrossRef]
- Tamura, M.; Matsui, K.; Kobayashi, Y.; Ogita, C.; Tsuboi, K.; Kusakabe, M.; Azuma, K.; Abe, T.; Yoshikawa, T.; Sekiguchi, M.; et al. A case of eel collagen allergy. Allergol. Int. 2018, 67, 138–140. [Google Scholar] [CrossRef]
- Linneberg, A.; Fenger, R.V.; Husemoen, L.L.N.; Thuesen, B.H.; Skaaby, T.; Gonzalez-Quintela, A.; Vidal, C.; Carlsen, B.C.; Johansen, J.D.; Menné, T.; et al. Association between loss-of-function mutations in the filaggrin gene and self-reported food allergy and alcohol sensitivity. Int. Arch. Allergy Immunol. 2013, 161, 234–242. [Google Scholar] [CrossRef]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatr. Allergy Immunol. 2016, 27, 1–250. [Google Scholar] [CrossRef]
- Aas, K.; Elsayed, S.M. Characterization of a major allergen (cod). Effect of enzymic hydrolysis on the allergenic activity. J. Allergy 1969, 44, 333–343. [Google Scholar] [CrossRef]
- Lim, D.L.C.; Neo, K.H.; Goh, D.L.M.; Shek, L.P.C.; Lee, B.W. Missing parvalbumin: Implications in diagnostic testing for tuna allergy [3]. J. Allergy Clin. Immunol. 2005, 115, 874–875. [Google Scholar] [CrossRef]
- Bugajska-Schretter, A.; Pastore, A.; Vangelista, L.; Rumpold, H.; Valenta, R.; Spitzauer, S. Molecular and immunological characterization of carp parvalbumin, a major fish allergen. Int. Arch. Allergy Immunol. 1999, 118, 306–308. [Google Scholar] [CrossRef]
- Hamada, Y.; Tanaka, H.; Ishizaki, S.; Ishida, M.; Nagashima, Y.; Shiomi, K. Purification, reactivity with IgE and cDNA cloning of parvalbumin as the major allergen of mackerels. Food Chem. Toxicol. 2003, 41, 1149–1156. [Google Scholar] [CrossRef]
- Arif, S.H. A Ca2+-binding protein with numerous roles and uses: Parvalbumin in molecular biology and physiology. BioEssays 2009, 31, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Stephen, J.N.; Sharp, M.F.; Ruethers, T.; Taki, A.; Campbell, D.E.; Lopata, A.L. Allergenicity of bony and cartilaginous fish—Molecular and immunological properties. Clin. Exp. Allergy 2017, 47, 300–312. [Google Scholar] [CrossRef]
- Kalic, T.; Morel-Codreanu, F.; Radauer, C.; Ruethers, T.; Taki, A.C.; Swoboda, I.; Hilger, C.; Hoffmann-Sommergruber, K.; Ollert, M.; Hafner, C.; et al. Patients Allergic to Fish Tolerate Ray Based on the Low Allergenicity of Its Parvalbumin. J. Allergy Clin. Immunol. Pract. 2019, 7, 500–508.e11. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Tanaka, H.; Hamada, Y.; Ishizaki, S.; Nagashima, Y.; Shiomi, K. Comparison of allergenicity and allergens between fish white and dark muscles. Allergy Eur. J. Allergy Clin. Immunol. 2006, 61, 357–363. [Google Scholar] [CrossRef]
- Valverde-Monge, M.; Pastor-Vargas, C.; Rodríguez del Rio, P.; Escudero, C.; Sánchez-García, S.; Mendez Brea, P.; Ibáñez, M.D. Anaphylaxis by exclusive allergy to swordfish and identification of a new fish allergen. Pediatr. Allergy Immunol. 2018, 29, 563–565. [Google Scholar] [CrossRef]
- Nakamura, R.; Satoh, R.; Nakajima, Y.; Kawasaki, N.; Yamaguchi, T.; Sawada, J.-i.; Nagoya, H.; Teshima, R. Comparative study of GH-transgenic and non-transgenic amago salmon (Oncorhynchus masou ishikawae) allergenicity and proteomic analysis of amago salmon allergens. Regul. Toxicol. Pharmacol. 2009, 55, 300–308. [Google Scholar] [CrossRef]
- Liu, R.; Krishnan, H.B.; Xue, W.; Liu, C. Characterization of allergens isolated from the freshwater fish blunt snout bream (Megalobrama amblycephala). J. Agric. Food Chem. 2011, 59, 458–463. [Google Scholar] [CrossRef]
- Kuehn, A.; Hilger, C.; Lehners-Weber, C.; Codreanu-Morel, F.; Morisset, M.; Metz-Favre, C.; Pauli, G.; de Blay, F.; Revets, D.; Muller, C.P.; et al. Identification of enolases and aldolases as important fish allergens in cod, salmon and tuna: Component resolved diagnosis using parvalbumin and the new allergens. Clin. Exp. Allergy 2013, 43, 811–822. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Toda, M.; Ebihara, T.; Irie, S.; Hori, H.; Imai, A.; Yanagida, M.; Miyazawa, H.; Ohsuna, H.; Ikezawa, Z.; et al. IgE antibody to fish gelatin (type I collagen) in patients with fish allergy Masahiro. J. Allergy Clin. Immunol. 2000, 106, 579–584. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Akiyama, H.; Huge, J.; Kubota, H.; Chikazawa, S.; Satoh, T.; Miyake, T.; Uhara, H.; Okuyama, R.; Nakagawara, R.; et al. Fish collagen is an important panallergen in the Japanese population. Allergy Eur. J. Allergy Clin. Immunol. 2016, 71, 720–723. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Kuriyama, T.; Nakagawara, R.; Aihara, M.; Hamada-Sato, N. Allergy to fish collagen: Thermostability of collagen and IgE reactivity of patients’ sera with extracts of 11 species of bony and cartilaginous fish. Allergol. Int. 2016, 65, 450–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.K.; Poulsen, L.K.; Stahl Skov, P.; Hefle, S.L.; Hlywka, J.J.; Taylor, S.L.; Bindslev-Jensen, U.; Bindslev-Jensen, C. A randomized, double-blinded, placebo-controlled oral challenge study to evaluate the allergenicity of commercial, food-grade fish gelatin. Food Chem. Toxicol. 2004, 42, 2037–2044. [Google Scholar] [CrossRef]
- Liu, R.; Holck, A.L.; Yang, E.; Liu, C.; Xue, W. Tropomyosin from tilapia (Oreochromis mossambicus) as an allergen. Clin. Exp. Allergy 2013, 43, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Karnaneedi, S.; Nie, S.; Kalic, T.; Dai, D.; Daduang, S.; Leeming, M.; Williamson, N.A.; Breiteneder, H.; et al. Expanding the allergen repertoire of salmon and catfish. Allergy 2020, all.14574. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gordo, M.; Sanchez-Garcia, S.; Cases, B.; Pastor, C.; Vivanco, F.; Cuesta-Herranz, J. Identification of vitellogenin as an allergen in Beluga caviar allergy. Allergy Eur. J. Allergy Clin. Immunol. 2008, 63, 479–480. [Google Scholar] [CrossRef] [PubMed]
- Das Dores, S.; Chopin, C.; Romano, A.; Galland-Irmouli, A.V.; Quaratino, D.; Pascual, C.; Fleurence, J.; Guéant, J.L. IgE-binding and cross-reactivity of a new 41 kDa allergen of codfish. Allergy Eur. J. Allergy Clin. Immunol. Suppl. 2002, 57, 84–87. [Google Scholar] [CrossRef]
- Swoboda, I.; Balic, N.; Klug, C.; Focke, M.; Weber, M.; Spitzauer, S.; Neubauer, A.; Quirce, S.; Douladiris, N.; Papadopoulos, N.G.; et al. A general strategy for the generation of hypoallergenic molecules for the immunotherapy of fish allergy. J. Allergy Clin. Immunol. 2013, 132, 979–982. [Google Scholar] [CrossRef]
- Bugajska-Schretter, A.; Rumpold, H.; Spitzauer, S.; Grote, M.; Reichelt, R.; Vangelista, L.; Valent, P.; Sperr, W.R.; Pastore, A.; Valenta, R. Purification, biochemical, and immunological characterisation of a major food allergen: Different immunoglobulin E recognition of the apo- and calcium-bound forms of carp parvalbumin. Gut 2000, 46, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Bugajska-Schretter, A.; Elfman, L.; Fuchs, T.; Kapiotis, S.; Rumpold, H.; Valenta, R.; Spitzauer, S. Parvalbumin, a cross-reactive fish allergen, contains IgE-binding epitopes sensitive to periodate treatment and Ca2+ depletion. J. Allergy Clin. Immunol. 1998, 101, 67–74. [Google Scholar] [CrossRef]
- Van Do, T.; Hordvik, I.; Endresen, C.; Elsayed, S. The major allergen (parvalbumin) of codfish is encoded by at least two isotypic genes: cDNA cloning, expression and antibody binding of the recombinant allergens. Mol. Immunol. 2003, 39, 595–602. [Google Scholar] [CrossRef]
- Sharp, M.F.; Kamath, S.D.; Koeberl, M.; Jerry, D.R.; O’Hehir, R.E.; Campbell, D.E.; Lopata, A.L. Differential IgE binding to isoallergens from Asian seabass (Lates calcarifer) in children and adults. Mol. Immunol. 2014, 62, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Kalic, T.; Kamath, S.D.; Ruethers, T.; Taki, A.C.; Nugraha, R.; Le, T.T.K.; Humeniuk, P.; Williamson, N.A.; Hira, D.; Rolland, J.M.; et al. Collagen—An Important Fish Allergen for Improved Diagnosis. J. Allergy Clin. Immunol. Pract. 2020, 8, 3084–3092.e10. [Google Scholar] [CrossRef] [PubMed]
- Griesmeier, U.; Vázquez-Cortés, S.; Bublin, M.; Radauer, C.; Ma, Y.; Briza, P.; Fernández-Rivas, M.; Breiteneder, H. Expression levels of parvalbumins determine allergenicity of fish species. Allergy Eur. J. Allergy Clin. Immunol. 2010, 65, 191–198. [Google Scholar] [CrossRef]
- Kondo, Y.; Ahn, J.; Komatsubara, R.; Terada, A.; Yasüda, T.; Tsuge, I.; Urisu, A. Comparison of allergenic properties of Salmon (Oncorhynchus nerka) between landlocked and anadromous species. Allergol. Int. 2009, 58, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Ruethers, T.; Raith, M.; Sharp, M.F.; Koeberl, M.; Stephen, J.N.; Nugraha, R.; Le, T.T.K.; Quirce, S.; Nguyen, H.X.M.; Kamath, S.D.; et al. Characterization of Ras k 1 a novel major allergen in Indian mackerel and identification of parvalbumin as the major fish allergen in 33 Asia-Pacific fish species. Clin. Exp. Allergy 2018, 48, 452–463. [Google Scholar] [CrossRef]
- Lindstrøm, C.D.V.; Van Dô, T.; Hordvik, I.; Endresen, C.; Elsayed, S. Cloning of two distinct cDNAs encoding parvalbumin, the major allergen of Atlantic salmon (Salmo salar). Scand. J. Immunol. 1996, 44, 335–344. [Google Scholar] [CrossRef]
- Gajewski, K.G.; Hsieh, Y.H.P. Monoclonal antibody specific to a major fish allergen: Parvalbumin. J. Food Prot. 2009, 72, 818–825. [Google Scholar] [CrossRef]
- Van Do, T.; Elsayed, S.; Florvaag, E.; Hordvik, I.; Endresen, C. Allergy to fish parvalbumins: Studies on the cross-reactivity of allergens from 9 commonly consumed fish. J. Allergy Clin. Immunol. 2005, 116, 1314–1320. [Google Scholar] [CrossRef]
- Shimizu, Y.; Nakamura, A.; Kishimura, H.; Hara, A.; Watanabe, K.; Saeki, H. Major allergen and its IgE cross-reactivity among salmonid fish roe allergy. J. Agric. Food Chem. 2009, 57, 2314–2319. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A.; Nugraha, R.; Karnaneedi, S.; Cao, T.; Dai, D.; Shanmuganathan, T.; Leeming, M.; Nie, S.; Williamson, N.; et al. IgE-reactive proteins defined by 88 fish-allergic children, predicting the allergenicity of 66 Asia-Pacific fish species. Allergy 2019, 74, 109. [Google Scholar]
- Kuehn, A.; Hutt-Kempf, E.; Hilger, C.; Hentges, F. Clinical monosensitivity to salmonid fish linked to specific IgE-epitopes on salmon and trout beta-parvalbumins. Allergy Eur. J. Allergy Clin. Immunol. 2011, 66, 299–301. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Cortés, S.; Nuez-Acevedo, B.; Jimeno-Nogales, L.; Ledesma, A.; Fernández-Rivas, M. Selective allergy to the salmonidae fish family: A selective parvalbumin epitope? Ann. Allergy Asthma Immunol. 2012, 108, 62–63. [Google Scholar] [CrossRef] [PubMed]
- Asero, R.; Mistrello, G.; Roncarolo, D.; Casarini, M.; Falagiani, P. True monosensitivity to a tropical sole. Allergy Eur. J. Allergy Clin. Immunol. 1999, 54, 1228–1229. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy. J. Allergy Clin. Immunol. 2010. [Google Scholar] [CrossRef]
- Lopata, A.L.; O’Hehir, R.E.; Lehrer, S.B. Shellfish allergy. Clin. Exp. Allergy 2010, 40, 850–858. [Google Scholar] [CrossRef]
- González-Fernández, J.; Alguacil-Guillén, M.; Cuéllar, C.; Daschner, A. Possible Allergenic Role of Tropomyosin in Patients with Adverse Reactions after Fish Intake. Immunol. Investig. 2018, 47, 416–429. [Google Scholar] [CrossRef]
- Peixoto, S.; Monteiro, T.; Carvalho, M.; Santos, M.; Matos, C.; Bartolomé, B.; Labrador-Horrillo, M.; Quaresma, M. Vertebrate tropomyosin as an allergen. J. Investig. Allergol. Clin. Immunol. 2018, 28, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Hilger, C.; Thill, L.; Grigioni, F.; Lehners, C.; Falagiani, P.; Ferrara, A.; Romano, C.; Stevens, W.; Hentges, F. IgE-antibodies of fish allergic patients cross-react with frog parvalbumin. Allergy Eur. J. Allergy Clin. Immunol. 2004, 59, 653–660. [Google Scholar] [CrossRef]
- Kuehn, A.; Codreanu-Morel, F.; Lehners-Weber, C.; Doyen, V.; Gomez-André, S.A.; Bienvenu, F.; Fischer, J.; Ballardini, N.; van Hage, M.; Perotin, J.M.; et al. Cross-reactivity to fish and chicken meat—A new clinical syndrome. Allergy Eur. J. Allergy Clin. Immunol. 2016, 71, 1772–1781. [Google Scholar] [CrossRef]
- Ruethers, T.; Nugraha, R.; Taki, A.C.; Mehr, S.S.; Campbell, D.E.; Lopata, A.L. Crocodile parvalbumin is a new allergen: The fish-crocodile syndrome. Allergy 2018, 73, 493. [Google Scholar]
- Xu, L.L.; Lin, H.; Li, Z.X.; Ahmed, I.; Pramod, S.N.; Lin, H.; Lv, L.T.; Tian, S.L.; Yu, Z.W. Influence of nonthermal extraction technique and allergenicity characteristics of tropomyosin from fish (Larimichthys crocea) in comparison with shrimp (Litopenaeus vannamei) and clam (Ruditapes philippinarum). Food Chem. 2020, 309, 125575. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.L.; Chen, J.; Sun, L.R.; Gao, X.; Lin, H.; Ahmed, I.; Pramod, S.N.; Li, Z.X. Analysis of the allergenicity and B cell epitopes in tropomyosin of shrimp (Litopenaeus vannamei) and correlation to cross-reactivity based on epitopes with fish (Larimichthys crocea) and clam (Ruditapes philippinarum). Food Chem. 2020, 323, 126763. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Khangurha, J.; Roberts, J.; Buddhadasa, S.; Clarke, D.; Hedges, C.E.; Campbell, D.E.; Kamath, S.D.; Lopata, A.L.; et al. Commercial fish ELISA kits have a limited capacity to detect different fish species and their products. J. Sci. Food Agric. 2020, 100, 4353–4363. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Scheuermann, T.; Hilger, C.; Hentges, F. Important variations in parvalbumin content in common fish species: A factor possibly contributing to variable allergenicity. Int. Arch. Allergy Immunol. 2019, 153, 359–366. [Google Scholar] [CrossRef]
- Lewis, T.; Zotterman, Y. Vascular reactions of the skin to injury. J. Physiol. 1927, 62, 280–288. [Google Scholar] [CrossRef]
- Friedman, M.M.; Kaliner, M. Ultrastructural changes in human skin mast cells during antigen-induced degranulation in vivo. J. Allergy Clin. Immunol. 1988, 82, 998–1005. [Google Scholar] [CrossRef]
- Heinzerling, L.; Mari, A.; Bergmann, K.C.; Bresciani, M.; Burbach, G.; Darsow, U.; Durham, S.; Fokkens, W.; Gjomarkaj, M.; Haahtela, T.; et al. The skin prick test—European standards. Clin. Transl. Allergy 2013, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, I.L.; Li, J.T.; Bernstein, D.I.; Hamilton, R.; Spector, S.L.; Tan, R.; Sicherer, S.; Golden, D.B.K.; Khan, D.A.; Nicklas, R.A.; et al. Allergy diagnostic testing: An updated practice parameter. Ann. Allergy Asthma Immunol. 2008, 100. [Google Scholar] [CrossRef]
- Pitsios, C.; Dimitriou, A.; Stefanaki, E.C.; Kontou-Fili, K. Anaphylaxis during skin testing with food allergens in children. Eur. J. Pediatr. 2010, 169, 613–615. [Google Scholar] [CrossRef]
- Pitsios, C.; Dimitriou, A.; Kontou-Fili, K. Allergic reactions during allergy skin testing with food allergens. Eur. Ann. Allergy Clin. Immunol. 2009, 41, 126–128. [Google Scholar] [PubMed]
- Haktanir Abul, M.; Orhan, F. Anaphylaxis after prick-to-prick test with fish. Pediatr. Int. 2016, 58, 503–505. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Nugraha, R.; Cao, T.T.; Koeberl, M.; Kamath, S.D.; Williamson, N.A.; O’Callaghan, S.; Nie, S.; Mehr, S.S.; et al. Variability of allergens in commercial fish extracts for skin prick testing. Allergy Eur. J. Allergy Clin. Immunol. 2019, 74, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.S.; Yuen, A.W.T.; Wai, C.Y.Y.; Leung, N.Y.H.; Chu, K.H.; Leung, P.S.C. Diagnosis of fish and shellfish allergies. J. Asthma Allergy 2018, 11, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.A.; Burks, A.W. Food Allergies: Prevalence, Molecular Characterization, and Treatment/Prevention Strategies. Annu. Rev. Nutr. 2006, 26, 539–565. [Google Scholar] [CrossRef]
- Turner, P.; Ng, I.; Kemp, A.; Campbell, D. Seafood allergy in children: A descriptive study. Ann. Allergy Asthma Immunol. 2011, 106, 494–501. [Google Scholar] [CrossRef]
- Chikazawa, S.; Hashimoto, T.; Kobayashi, Y.; Satoh, T. Fish-collagen allergy: A pitfall of the prick-to-prick test with raw fish. Br. J. Dermatol. 2015, 173, 1330–1331. [Google Scholar] [CrossRef]
- Wang, J.; Godbold, J.H.; Sampson, H.A. Correlation of serum allergy (IgE) tests performed by different assay systems. J. Allergy Clin. Immunol. 2008, 121, 1219–1224. [Google Scholar] [CrossRef]
- Sampson, H.A.; Ho, D.G. Relationship between food-specific IgE concentrations and the risk of positive food challenges in children and adolescents. J. Allergy Clin. Immunol. 1997, 100, 444–451. [Google Scholar] [CrossRef]
- Boyano Martínez, T.; García-Ara, C.; Díaz-Pena, J.M.; Muñoz, F.M.; García Sánchez, G.; Esteban, M.M. Validity of specific IgE antibodies in children with egg allergy. Clin. Exp. Allergy 2001, 31, 1464–1469. [Google Scholar] [CrossRef]
- Buyuktiryaki, B.; Cavkaytar, O.; Sahiner, U.M.; Yilmaz, E.A.; Yavuz, S.T.; Soyer, O.; Sekerel, B.E.; Tuncer, A.; Sackesen, C. Cor a 14, Hazelnut-Specific IgE, and SPT as a Reliable Tool in Hazelnut Allergy Diagnosis in Eastern Mediterranean Children. J. Allergy Clin. Immunol. Pract. 2016, 4, 265–272.e3. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A. Utility of food-specific IgE concentrations in predicting symptomatic food allergy. J. Allergy Clin. Immunol. 2001, 107, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, N.Y.; Sicherer, S.H. Interpreting IgE sensitization tests in food allergy. Expert Rev. Clin. Immunol. 2016, 12, 389–403. [Google Scholar] [CrossRef]
- Rubin, Z.E.; Gu, H.; Polk, B.I. Seafood graded oral food challenge outcomes in a pediatric tertiary care center. World Allergy Organ. J. 2020, 13, 100121. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, M.; Kuehn, A.; Mills, E.N.C.; Costello, C.A.; Ollert, M.; Småbrekke, L.; Primicerio, R.; Wickman, M.; Klingenberg, C. Cross-reactivity in fish allergy: A double-blind, placebo-controlled food-challenge trial. J. Allergy Clin. Immunol. 2017, 140, 1170–1172. [Google Scholar] [CrossRef] [Green Version]
- Niggemann, B.; Beyer, K. Diagnosis of food allergy in children: Toward a standardization of food challenge. J. Pediatr. Gastroenterol. Nutr. 2007, 45, 399–404. [Google Scholar] [CrossRef]
- Bindslev-Jensen, C.; Ballmer-Welser, B.K.; Bengtsson, U.; Blanco, C.; Ebner, C.; Hourihane, J.; Knulst, A.C.; Moneret-Vautrin, D.A.; Nekam, K.; Niggemann, B.; et al. Standardization of food challenges in patients with immediate reactions to foods—Position paper from the European Academy of Allergology and Clinical Immunology. Allergy Eur. J. Allergy Clin. Immunol. 2004, 59, 690–697. [Google Scholar] [CrossRef]
- Crevel, R.W.R.; Ballmer-Weber, B.K.; Holzhauser, T.; Hourihane, J.O.B.; Knulst, A.C.; Mackie, A.R.; Timmermans, F.; Taylor, S.L. Thresholds for food allergens and their value to different stakeholders. Allergy Eur. J. Allergy Clin. Immunol. 2008, 63, 597–609. [Google Scholar] [CrossRef]
- Taylor, S.L.; Hefle, S.L.; Bindslev-Jensen, C.; Bock, S.A.; Burks, A.W.; Christie, L.; Hill, D.J.; Host, A.; Hourihane, J.O.B.; Lack, G.; et al. Factors affecting the determination of threshold doses for allergenic foods: How much is too much? J. Allergy Clin. Immunol. 2002, 109, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Ballmer-Weber, B.K.; Fernandez-Rivas, M.; Beyer, K.; Defernez, M.; Sperrin, M.; Mackie, A.R.; Salt, L.J.; Hourihane, J.O.B.; Asero, R.; Belohlavkova, S.; et al. How much is too much? Threshold dose distributions for 5 food allergens. J. Allergy Clin. Immunol. 2015, 135, 964–971. [Google Scholar] [CrossRef]
- Nowak-Wegrzyn, A.; Assa’ad, A.H.; Bahna, S.L.; Bock, S.A.; Sicherer, S.H.; Teuber, S.S. Work Group report: Oral food challenge testing. J. Allergy Clin. Immunol. 2009, 123, S365–S383. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Gerth Van Wijk, R.; Bindslev-Jensen, C.; Sicherer, S.; Teuber, S.S.; Burks, A.W.; Dubois, A.E.J.; Beyer, K.; Eigenmann, P.A.; Spergel, J.M.; et al. Standardizing double-blind, placebo-controlled oral food challenges: American Academy of Allergy, Asthma & Immunology-European Academy of Allergy and Clinical Immunology PRACTALL consensus report. J. Allergy Clin. Immunol. 2012, 130, 1260–1274. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rivas, M.; Barreales, L.; Mackie, A.R.; Fritsche, P.; Vázquez-Cortés, S.; Jedrzejczak-Czechowicz, M.; Kowalski, M.L.; Clausen, M.; Gislason, D.; Sinaniotis, A.; et al. The EuroPrevall outpatient clinic study on food allergy: Background and methodology. Allergy Eur. J. Allergy Clin. Immunol. 2015, 70, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Vassilopoulou, E.; Douladiris, N.; Sakellariou, A.; Cortes, S.V.; Sinaniotis, A.; Rivas, M.F.; Papadopoulos, N.G. Evaluation and standardisation of different matrices used for double-blind placebo-controlled food challenges to fish. J. Hum. Nutr. Diet. 2010, 23, 544–549. [Google Scholar] [CrossRef]
- Klemans, R.J.B.; Broekman, H.C.H.P.; Knol, E.F.; Bruijnzeel-Koomen, C.A.F.M.; Otten, H.G.; Pasmans, S.G.M.A.; Knulst, A.C. Ara h 2 is the best predictor for peanut allergy in adults. J. Allergy Clin. Immunol. Pract. 2013, 1, 632–638.e1. [Google Scholar] [CrossRef]
- Huss-Marp, J.; Gutermuth, J.; Schäffner, I.; Darsow, U.; Pfab, F.; Brockow, K.; Ring, J.; Behrendt, H.; Jakob, T.; Ahlgrim, C. Comparison of molecular and extract-based allergy diagnostics with multiplex and singleplex analysis. Allergo J. Int. 2015, 24, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Abreu Carvalho, S.; Alves Marcelino, J.; Cabral Duarte De Moura Portugal, M.; Lopes Costa Hipólito, A.; de Castro Pereira Barbosa, M.; Galvão Barata Pinto Pereira Dos Santos, M. Contribution of recombinant Parvalbumin Gad c 1 in the diagnosis and prognosis of fish allergy. J. Investig. Allergol. Clin. Immunol. 2019, 30. [Google Scholar] [CrossRef]
- Douladiris, N.; Linhart, B.; Swoboda, I.; Gstöttner, A.; Vassilopoulou, E.; Stolz, F.; Valenta, R.; Papadopoulos, N.G. In vivo allergenic activity of a hypoallergenic mutant of the major fish allergen CYP c 1 evaluated by means of skin testing. J. Allergy Clin. Immunol. 2015, 136, 493–495.e8. [Google Scholar] [CrossRef]
- Kelso, J.M.; Jones, R.T.; Yunginger, J.W. Monospecific allergy to swordfish. Ann. Allergy Asthma Immunol. 1996, 77, 227–228. [Google Scholar] [CrossRef]
- Kuehn, A.; Fischer, J.; Hilger, C.; Sparla, C.; Biedermann, T.; Hentges, F. Correlation of clinical monosensitivity to cod with specific IgE to enolase and aldolase. Ann. Allergy Asthma Immunol. 2014, 113, 670–671.e2. [Google Scholar] [CrossRef]
- Buyuktiryaki, B.; Santos, A.F. Food allergy severity predictions based on cellular in vitro tests. Expert Rev. Mol. Diagn. 2020, 20, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, O.; Kwok, M.; McKendry, R.; Santos, A.F. Basophil Activation Test: Old and New Applications in Allergy. Curr. Allergy Asthma Rep. 2018, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Wang, J.; Leung, N.; Wang, L.X.; Lisann, L.; Sicherer, S.H.; Scurlock, A.M.; Pesek, R.; Perry, T.T.; Jones, S.M.; et al. Correlations between basophil activation, allergen-specific IgE with outcome and severity of oral food challenges. Ann. Allergy Asthma Immunol. 2015, 114, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Imakiire, R.; Fujisawa, T.; Nagao, M.; Tokuda, R.; Hattori, T.; Kainuma, K.; Kawano, Y. Basophil Activation Test Based on CD203c Expression in the Diagnosis of Fish Allergy. Allergy. Asthma Immunol. Res. 2020, 12, 641. [Google Scholar] [CrossRef] [PubMed]
- Burney, P.G.J.; Potts, J.; Kummeling, I.; Mills, E.N.C.; Clausen, M.; Dubakiene, R.; Barreales, L.; Fernandez-Perez, C.; Fernandez-Rivas, M.; Le, T.M.; et al. The prevalence and distribution of food sensitization in European adults. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Kobayashi, Y.; Shiomi, K. Fish allergy in patients with parvalbumin-specific immunoglobulin E depends on parvalbumin content rather than molecular differences in the protein among fish species. Biosci. Biotechnol. Biochem. 2016, 80, 2018–2021. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.M.; Gupta, R.S.; Aktas, O.N.; Diaz, V.; Kamath, S.D.; Lopata, A.L. Clinical Management of Seafood Allergy. J. Allergy Clin. Immunol. Pract. 2020, 8, 37–44. [Google Scholar] [CrossRef]
- De Martino, M.; Novembre, E.; Galli, L.; de Marco, A.; Botarelli, P.; Marano, E.; Vierucci, A. Allergy to different fish species in cod-allergic children: In vivo and in vitro studies. J. Allergy Clin. Immunol. 1990, 86, 909–914. [Google Scholar] [CrossRef]
- Bernhisel-Broadbent, J.; Scanlon, S.M.; Sampson, H.A. Fish hypersensitivity. I. In vitro and oral challenge results in fish-allergic patients. J. Allergy Clin. Immunol. 1992, 89, 730–737. [Google Scholar] [CrossRef]
- Helbling, A.; Haydel, R.; McCants, M.L.; Musmand, J.J.; El-Dahr, J.; Lehrer, S.B. Fish allergy: Is cross-reactivity among fish species relevant? Double-blind placebo-controlled food challenge studies of fish allergic adults. Ann. Allergy Asthma Immunol. 1999, 83, 517–523. [Google Scholar] [CrossRef]
- Mourad, A.A.; Bahna, S.L. Fish-allergic patients may be able to eat fish. Expert Rev. Clin. Immunol. 2015, 11, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Bernhisel-Broadbent, J.; Strause, D.; Sampson, H.A. Fish hypersensitivity. II: Clinical relevance of altered fish allergenicity caused by various preparation methods. J. Allergy Clin. Immunol. 1992, 90, 622–629. [Google Scholar] [CrossRef]
- Mejrhit, N.; Azdad, O.; Aarab, L. Effect of industrial processing on the IgE reactivity of three commonly consumed moroccan fish species in Fez region. Eur. Ann. Allergy Clin. Immunol. 2018, 50, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Barbarroja-Escudero, J.; Sánchez-González, M.J.; Bartolomé, B.; Pastor-Vargas, C.; Rodríguez-Rodríguez, M.; Cuesta-Herranz, J.; Alvarez-Mon, M. Actin as a possible cross-reactive allergen between fish and poultry. J. Investig. Allergol. Clin. Immunol. 2019, 29, 152–154. [Google Scholar] [CrossRef]
- Haroun-Díaz, E.; Blanca-López, N.; Vázquez de la Torre, M.; Ruano, F.J.; Somoza Álvarez, M.L.; Labrador Horrillo, M.; Bartolomé, B.; Blanca, M.; Canto Díez, G. Severe anaphylaxis due to crocodile-meat allergy exhibiting wide cross-reactivity with fish allergens. J. Allergy Clin. Immunol. Pract. 2018, 6, 669–670.e1. [Google Scholar] [CrossRef]
- Añíbarro, B.; Seoane, F.J.; Múgica, M.V. Involvement of hidden allergens in food allergic reactions. J. Investig. Allergol. Clin. Immunol. 2007, 17, 168–172. [Google Scholar]
- Bertelsen, R.J.; Faeste, C.K.; Granum, B.; Egaas, E.; London, S.J.; Carlsen, K.H.; Lødrup Carlsen, K.C.; Løvik, M. Food allergens in mattress dust in Norwegian homes—A potentially important source of allergen exposure. Clin. Exp. Allergy 2014, 44, 142–149. [Google Scholar] [CrossRef] [Green Version]
- DunnGalvin, A.; Koman, E.; Raver, E.; Frome, H.; Adams, M.; Keena, A.; Hourihane, J.O.B.; Gallagher, P.L.; Flokstra-de Blok, B.; Dubois, A.; et al. An Examination of the Food Allergy Quality of Life Questionnaire Performance in a Countrywide American Sample of Children: Cross-Cultural Differences in Age and Impact in the United States and Europe. J. Allergy Clin. Immunol. Pract. 2017, 5, 363–368.e2. [Google Scholar] [CrossRef]
- Lau, G.Y.; Patel, N.; Umasunthar, T.; Gore, C.; Warner, J.O.; Hanna, H.; Phillips, K.; Mohd Zaki, A.; Hodes, M.; Boyle, R.J. Anxiety and stress in mothers of food-allergic children. Pediatr. Allergy Immunol. 2014, 25, 236–242. [Google Scholar] [CrossRef]
- Annunziato, R.A.; Rubes, M.; Ambrose, M.A.; Mullarkey, C.; Shemesh, E.; Sicherer, S.H. Longitudinal evaluation of food allergy-related bullying. J. Allergy Clin. Immunol. Pract. 2014, 2, 639–641. [Google Scholar] [CrossRef]
- Herbert, L.; Shemesh, E.; Bender, B. Clinical Management of Psychosocial Concerns Related to Food Allergy. J. Allergy Clin. Immunol. Pract. 2016, 4, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Freeman, J. “Rush” inoculation, with special reference to hay-fever treatment. Lancet 1930, 215, 744–747. [Google Scholar] [CrossRef]
- Patriarca, G.; Nucera, E.; Roncallo, C.; Pollastrini, E.; Bartolozzi, F.; De Pasquale, T.; Buonomo, A.; Gasbarrini, G.; Di Campli, C.; Schiavino, D. Oral desensitizing treatment in food allergy: Clinical and immunological results. Aliment. Pharmacol. Ther. 2003, 17, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Patriarca, G.; Nucera, E.; Pollastrini, E.; Roncallo, C.; De Pasquale, T.; Lombardo, C.; Pedone, C.; Gasbarrini, G.; Buonomo, A.; Schiavino, D. Oral specific desensitization in food-allergic children. Dig. Dis. Sci. 2007, 52, 1662–1672. [Google Scholar] [CrossRef] [PubMed]
- Martorell-Calatayud, C.; Carnés, J.; Michavila Gómez, A.; Dopazo Fernández, L.; Echeverría Zudaire, L.; Santana Rodríguez, C.; Gómez Galán, C.; Toral Pérez, T.; Rodríguez del Río, P.; Martorell Aragonés, A. Oral immunotherapy to hake in 8 pediatric patients. J. Investig. Allergol. Clin. Immunol. 2019, 29, 294–332. [Google Scholar] [CrossRef] [Green Version]
- D’Amelio, C.; Gastaminza, G.; Vega, O.; Bernad, A.; Madamba, R.C.; Martínez-Aranguren, R.; Ferrer, M.; Goikoetxea, M.J. Induction of tolerance to different types of fish through desensitization with hake. Pediatr. Allergy Immunol. 2017, 28, 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untersmayr, E.; Szalai, K.; Riemer, A.B.; Hemmer, W.; Swoboda, I.; Hantusch, B.; Schöll, I.; Spitzauer, S.; Scheiner, O.; Jarisch, R.; et al. Mimotopes identify conformational epitopes on parvalbumin, the major fish allergen. Mol. Immunol. 2006, 43, 1454–1461. [Google Scholar] [CrossRef]
- Swoboda, I.; Bugajska-Schretter, A.; Linhart, B.; Verdino, P.; Keller, W.; Schulmeister, U.; Sperr, W.R.; Valent, P.; Peltre, G.; Quirce, S.; et al. A Recombinant Hypoallergenic Parvalbumin Mutant for Immunotherapy of IgE-Mediated Fish Allergy. J. Immunol. 2007, 178, 6290–6296. [Google Scholar] [CrossRef] [Green Version]
- Zuidmeer-Jongejan, L.; Huber, H.; Swoboda, I.; Rigby, N.; Versteeg, S.A.; Jensen, B.M.; Quaak, S.; Akkerdaas, J.H.; Blom, L.; Asturias, J.; et al. Development of a hypoallergenic recombinant parvalbumin for first-in-man subcutaneous immunotherapy of fish allergy. Int. Arch. Allergy Immunol. 2015, 166, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Freidl, R.; Gstoettner, A.; Baranyi, U.; Swoboda, I.; Stolz, F.; Focke-Tejkl, M.; Wekerle, T.; van Ree, R.; Valenta, R.; Linhart, B. Blocking antibodies induced by immunization with a hypoallergenic parvalbumin mutant reduce allergic symptoms in a mouse model of fish allergy. J. Allergy Clin. Immunol. 2017, 139, 1897–1905.e1. [Google Scholar] [CrossRef] [Green Version]
- Barni, S.; Liccioli, G.; Sarti, L.; Giovannini, M.; Novembre, E.; Mori, F. Immunoglobulin E (IgE)-mediated food allergy in children: Epidemiology, pathogenesis, diagnosis, prevention, and management. Medicina 2020, 56, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Fish Species | Common Name | Nomenclature | Order | Allergen | MW * (kDa) | Ref |
|---|---|---|---|---|---|---|
| Clupea harengus | Atlantic herring | Clu h 1 | Clupeiformes | β-parvalbumin | 12 | [99] |
| Cyprinus carpio | Common carp | Cyp c 1 | Cypriniformes | β-parvalbumin | 12 | [100] |
| Gadus callarias | Baltic cod | Gad c 1 | Gadiformes | β-parvalbumin | 12 | [101] |
| Gadus morhua | Atlantic cod | Gad m 1 | Gadiformes | β-parvalbumin | 12 | [102] |
| Atlantic cod | Gad m 2 | Gadiformes | β-enolase | 47.3 | [90] | |
| Atlantic cod | Gad m 3 | Gadiformes | Aldolase A | 40 | [90] | |
| Lates calcarifer | Barramundi/Asian Seabass | Lat c 1 | Perciformes | β-parvalbumin | 11.5 | [103] |
| Barramundi/Asian Seabass | Lat c 6 | Perciformes | Collagen alpha | 130–140 | [104] | |
| Lepidorhombus whiffiagonis | Megrim, whiff, turbot fish | Lep w 1 | Pleuronectiformes | β-parvalbumin | 11.5 | [105] |
| Oncorhynchus mykiss | Rainbow trout | Onc m 1 | Salmoniformes | β-parvalbumin | 12 | [106] |
| Rastrelliger kanagurta | Indian mackerel | Ras k 1 | Scombriformes | β-parvalbumin | 11.3 | [107] |
| Salmo salar | Atlantic somon | Sal s 1 | Salmoniformes | β-parvalbumin | 12 | [108] |
| Atlantic somon | Sal s 2 | Salmoniformes | β-enolase | 47.3 | [90] | |
| Atlantic somon | Sal s 3 | Salmoniformes | Aldolase A | 40 | [90] | |
| Sardinops sagax | Pacific pilchard | Sar sa 1 | Clupeiformes | β-parvalbumin | 12 | [71] |
| Sebastes marinus | Ocean perch, redfish | Seb m 1 | Scorpaeniformes | β-parvalbumin | 11 | [109] |
| Thunnus albacares | Yellowfin tuna | Thu a 1 | Perciformes | β-parvalbumin | 11 | [110] |
| Yellowfin tuna | Thu a 2 | Perciformes | β-enolase | 50 | [90] | |
| Yellowfin tuna | Thu a 3 | Perciformes | Aldolase A | 40 | [90] | |
| Xiphias gladius | Swordfish | Xip g 1 | Perciformes | β-parvalbumin | 11.5 | [105] |
| Oreochromis mossambicus | Mozambique tilapia | Ore m 4 | Perciformes | Tropomyosin | 33 | [95] |
| Oncorhynchus keta | Chum salmon | Onc k 5 | Salmoniformes | Vitellogenin | 18 | [111] |
| Type of Fish | Presentation | Parvalbumin Content (mg/g) |
|---|---|---|
| Cod | Raw | 1.5–2.5 |
| Smoked | 1.0–1.3 | |
| Cooked | 1.3–1.9 | |
| Salmon | Raw | 1.9–2.5 |
| Smoked | 0.7–1.0 | |
| Cooked | 1.5–1.9 | |
| Carp | Raw | 2.5–5.0 |
| Cooked | 2.1–4.0 | |
| Tuna (white muscle) | Raw | 0.01–0.05 |
| Cooked | 0.01–0.03 | |
| Tuna (dark muscle) | Raw | ND |
| Tuna | Canned | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buyuktiryaki, B.; Masini, M.; Mori, F.; Barni, S.; Liccioli, G.; Sarti, L.; Lodi, L.; Giovannini, M.; du Toit, G.; Lopata, A.L.; et al. IgE-Mediated Fish Allergy in Children. Medicina 2021, 57, 76. https://doi.org/10.3390/medicina57010076
Buyuktiryaki B, Masini M, Mori F, Barni S, Liccioli G, Sarti L, Lodi L, Giovannini M, du Toit G, Lopata AL, et al. IgE-Mediated Fish Allergy in Children. Medicina. 2021; 57(1):76. https://doi.org/10.3390/medicina57010076
Chicago/Turabian StyleBuyuktiryaki, Betul, Marzio Masini, Francesca Mori, Simona Barni, Giulia Liccioli, Lucrezia Sarti, Lorenzo Lodi, Mattia Giovannini, George du Toit, Andreas Ludwig Lopata, and et al. 2021. "IgE-Mediated Fish Allergy in Children" Medicina 57, no. 1: 76. https://doi.org/10.3390/medicina57010076