
Long Noncoding RNA TALAM1 Is a Transcriptional Target of the RUNX2 Transcription Factor in Lung Adenocarcinoma
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Tissue Samples
2.2. Reverse Transcription and Quantitative Real-Time PCR (q-PCR)
2.3. Immunofluorescence Assay
2.4. Lentivirus Production and Lentiviral Infection of A549 Cells
2.5. ChIP-Seq Experiment and Data Analysis
2.6. ChIP and qPCR
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
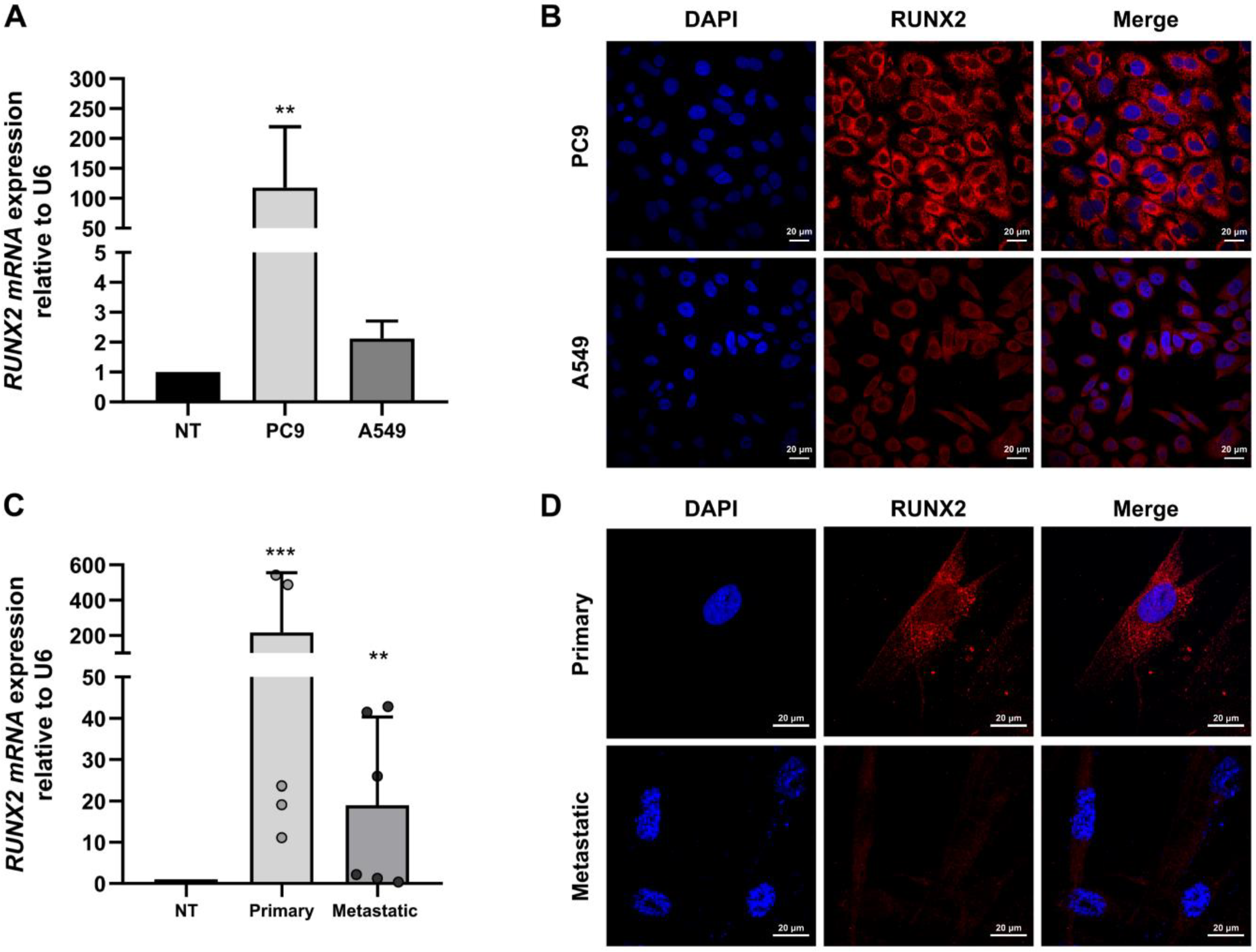
3.1. RUNX2 Is Overexpressed in Cell Lines and Primary Cultured Lung Cancer Cells
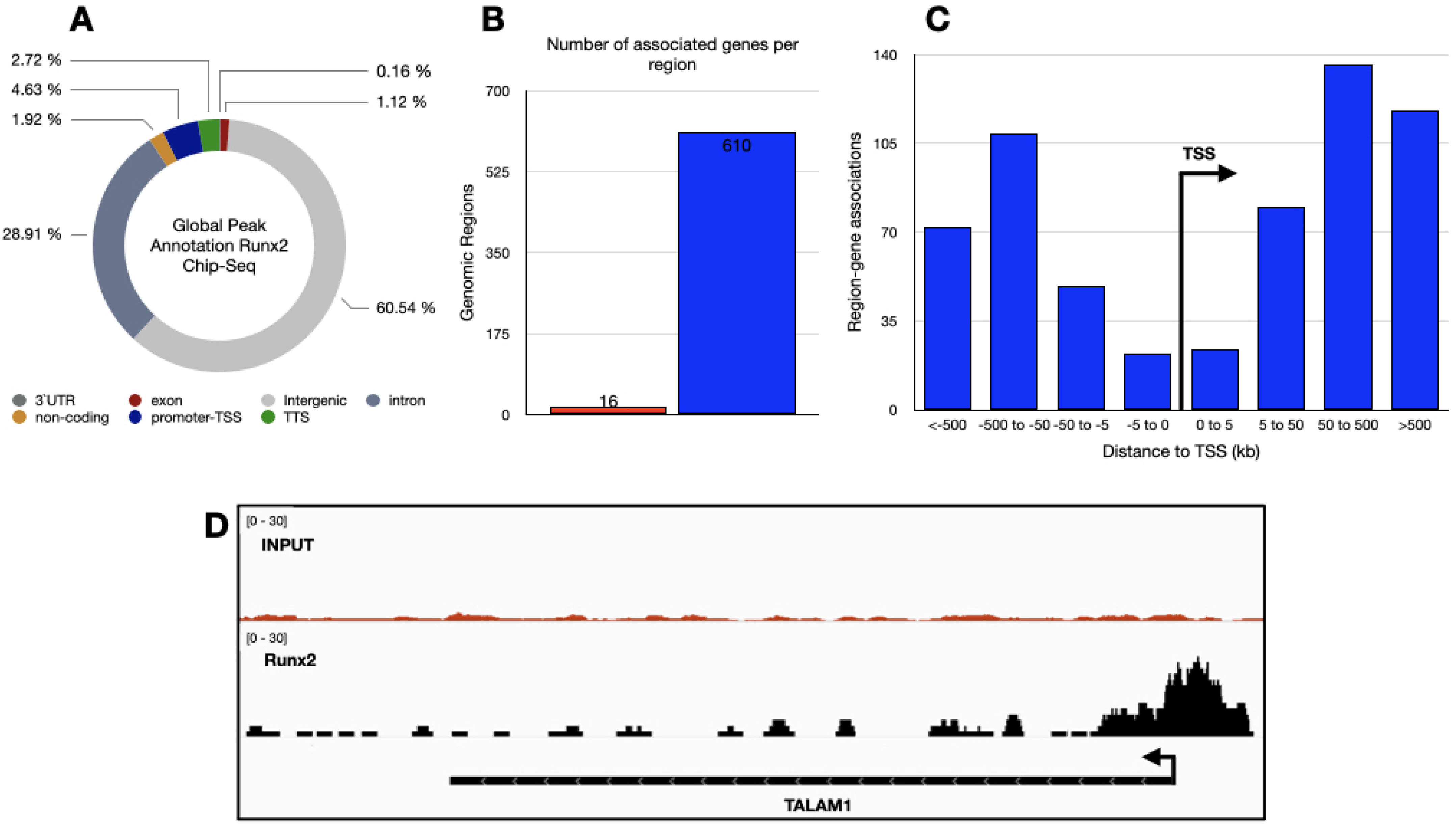
3.2. Genome-Wide Identification of RUNX2 Binding Sites in Lung Adenocarcinoma Tissue
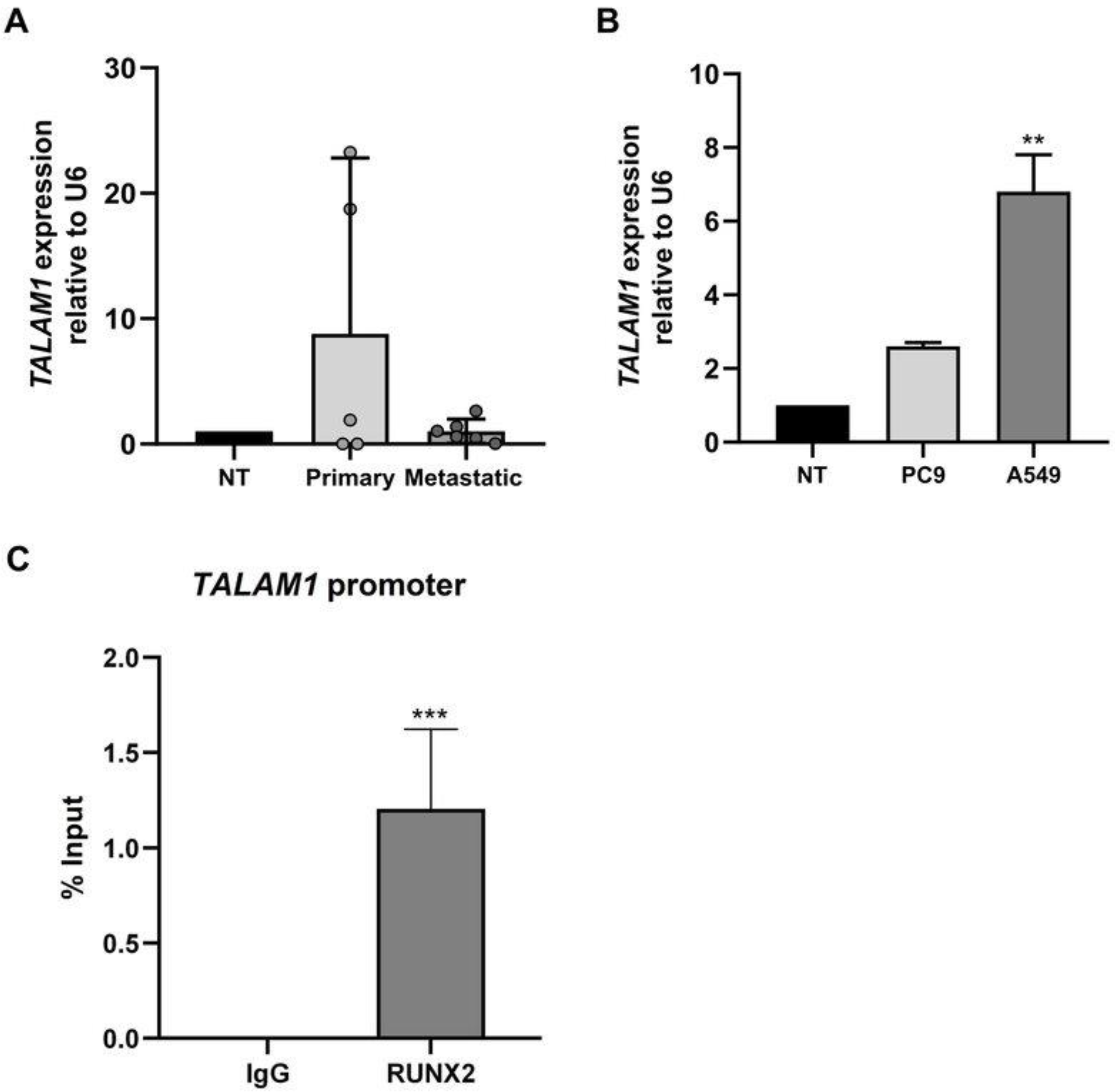
3.3. High Expression of TALAM1 in Human Primary Lung Cancer
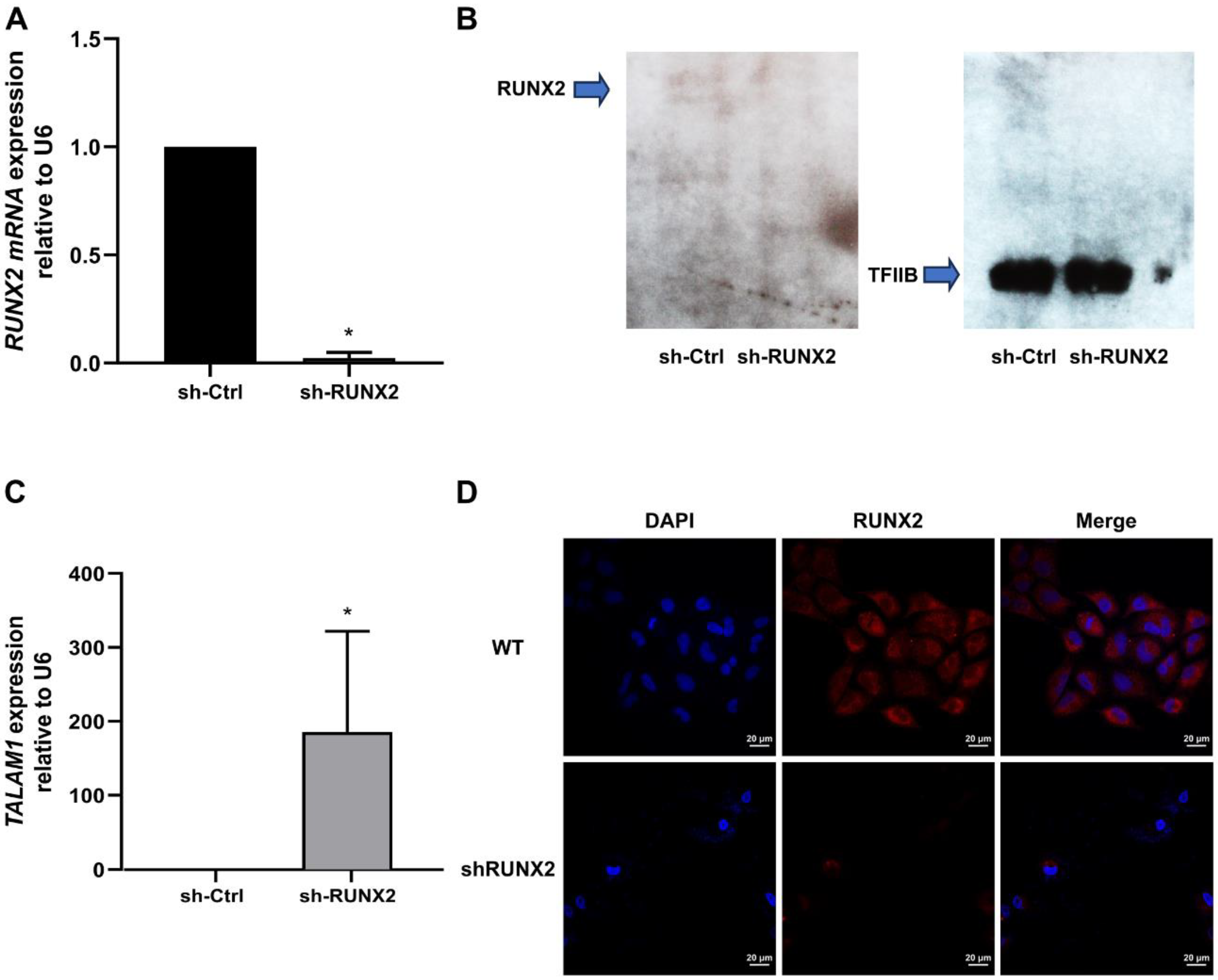
3.4. The RUNX2 TF Is Involved in Transcriptional Control of TALAM1 in Lung Adenocarcinoma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ChIP | Chromatin immunoprecipitation |
| ChIP-Seq | ChIP sequencing |
| Lnc | Long non coding |
| TF | Transcription Factor |
References
- Belluti, S.; Rigillo, G.; Imbriano, C. Transcription Factors in Cancer: When Alternative Splicing Determines Opposite Cell Fates. Cells 2020, 9, 760. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef]
- Bhagwat, A.S.; Vakoc, C.R. Targeting Transcription Factors in Cancer. Trends Cancer 2015, 1, 53–65. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Vishnoi, K.; Viswakarma, N.; Rana, A.; Rana, B. Transcription Factors in Cancer Development and Therapy. Cancers 2020, 12, 2296. [Google Scholar] [CrossRef]
- Rojas, A.; Otálora-Otálora, B.A.; Henríquez, B.; López-Kleine, L. RUNX family: Oncogenes or tumor suppressors (Review). Oncol. Rep. 2019, 42, 3–19. [Google Scholar] [CrossRef]
- Rojas, A.; Aguilar, R.; Henriquez, B.; Lian, J.B.; Stein, J.L.; Stein, G.S.; van Wijnen, A.J.; van Zundert, B.; Allende, M.L.; Montecino, M. Epigenetic Control of the Bone-master Runx2 Gene during Osteoblast-lineage Commitment by the Histone Demethylase JARID1B/KDM5B. J. Biol. Chem. 2015, 290, 28329–28342. [Google Scholar] [CrossRef]
- Ambrosini, C.; Garilli, F.; Quattrone, A. Reprogramming translation for gene therapy. Prog. Mol. Biol. Transl. Sci. 2021, 182, 439–476. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.B.; Rowell, E.; Sekimata, M. Epigenetic control of T-helper-cell differentiation. Nat. Rev. Immunol. 2009, 9, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Herreño, A.M.; Ramírez, A.C.; Chaparro, V.P.; Fernandez, M.J.; Cañas, A.; Morantes, C.F.; Moreno, O.M.; Brugés, R.E.; Mejía, J.A.; Bustos, F.J.; et al. Role of RUNX2 transcription factor in epithelial mesenchymal transition in non-small cell lung cancer: Epigenetic control of the RUNX2 P1 promoter. Tumor Biol. 2019, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Liu, K.; Chen, J.; Gong, Y.; Zhou, X.; Huang, J. RUNX2 as a Potential Prognosis Biomarker and New Target for Human Lung Cancer. Explor. Res. Hypothesis Med. 2021, 6, 99–109. [Google Scholar] [CrossRef]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding RNAs. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA–LncRNA Interactions. In Long Non-Coding RNAs; Humana Press: New York, NY, USA, 2016; Volume 1402, pp. 271–286. [Google Scholar] [CrossRef]
- Hu, Q.; Ma, H.; Chen, H.; Zhang, Z.; Xue, Q. LncRNA in tumorigenesis of non-small-cell lung cancer: From bench to bedside. Cell Death Discov. 2022, 8, 359. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.P.; Nóbrega-Pereira, S.; Silva, A.B.D.; Rebelo, K.; Alves-Vale, C.; Marinho, S.P.; Carvalho, T.; Dias, S.; De Jesus, B.B. An antisense transcript mediates MALAT1 response in human breast cancer. BMC Cancer 2019, 19, 771. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Hämmerle, M.; Eißmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Groß, M.; et al. The Noncoding RNA MALAT1 Is a Critical Regulator of the Metastasis Phenotype of Lung Cancer Cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Wang, Y.; Li, H.; Chen, L.; Liu, Q. Regulatory Networks of LncRNA MALAT-1 in Cancer. Cancer Manag. Res. 2020, 12, 10181–10198. [Google Scholar] [CrossRef]
- Lung Source: Globocan 2020 Number of New Cases in 2020, Both Sexes, All Ages. 2020. Available online: https://gco.iarc.fr/today (accessed on 13 September 2022).
- Cassim, S.; Chepulis, L.; Keenan, R.; Kidd, J.; Firth, M.; Lawrenson, R. Patient and carer perceived barriers to early presentation and diagnosis of lung cancer: A systematic review. BMC Cancer 2019, 19, 25. [Google Scholar] [CrossRef]
- Bernal, C.; Otalora, A.; Cañas, A.; Barreto, A.; Prieto, K.; Montecino, M.; Rojas, A. Regulatory Role of the RUNX2 Transcription Factor in Lung Cancer Apoptosis. Int. J. Cell Biol. 2022, 2022, 5198203. [Google Scholar] [CrossRef]
- McLean, C.Y.; Bristor, D.; Hiller, M.; Clarke, S.L.; Schaar, B.T.; Lowe, C.B.; Wenger, A.M.; Bejerano, G. GREAT improves functional interpretation of cis-regulatory regions. Nat. Biotechnol. 2010, 28, 495–501. [Google Scholar] [CrossRef]
- A549|ATCC. Available online: https://www.atcc.org/products/crm-ccl-185 (accessed on 19 January 2022).
- Croce, M.V.; Colussi, A.G.; Price, M.R.; Segal-Eiras, A. Identification and characterization of different subpopulations in a human lung adenocarcinoma cell line (A549). Pathol. Oncol. Res. 1999, 5, 197–204. [Google Scholar] [CrossRef]
- Othman, A.; Winogradzki, M.; Patel, S.; Holmes, W.; Blank, A.; Pratap, J. The Role of Runx2 in Microtubule Acetylation in Bone Metastatic Breast Cancer Cells. Cancers 2022, 14, 3436. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Yang, H.; Chai, J.; Xing, L. RUNX2 as a promising therapeutic target for malignant tumors. Cancer Manag. Res. 2021, 13, 2539–2548. [Google Scholar] [CrossRef] [PubMed]
- Zong, X.; Nakagawa, S.; Freier, S.M.; Fei, J.; Ha, T.; Prasanth, S.G.; Prasanth, K.V. Natural antisense RNA promotes 3′ end processing and maturation of MALAT1 lncRNA. Nucleic Acids Res. 2016, 44, 2898–2908. [Google Scholar] [CrossRef] [PubMed]
- Choe, M.; Brusgard, J.L.; Chumsri, S.; Bhandary, L.; Zhao, X.F.; Lu, S.; Goloubeva, O.G.; Polster, B.M.; Fiskum, G.M.; Girnun, G.D.; et al. The RUNX2 Transcription Factor Negatively Regulates SIRT6 Expression to Alter Glucose Metabolism in Breast Cancer Cells. J. Cell. Biochem. 2015, 116, 2210–2226. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Tumoral | NSCLC Cases | |
|---|---|---|
| Total individuals | 6 | 11 |
| Female | 2 (33%) | 8 (72.7%) |
| Male | 4 (67%) | 3 (27.3%) |
| Age | 66.5 (61–84) years | 67.7 (60–81) years |
| Histological tumor type | ||
| Adenocarcinoma | -- | 7 (63.6%) |
| Squamous cell carcinoma | -- | 4 (36.4%) |
| Lung cancer origin | ||
| Primary | -- | 5 (45.5%) |
| Metastatic | -- | 6 (54.5%) |
| TNM stage | ||
| II | -- | 1 (9%) |
| IIIB | -- | 1 (9%) |
| IV | -- | 9 (82%) |
| Immunohistochemical markers | ||
| P63 Positive | -- | 4 (36.4%) |
| P63 Negative | -- | 3 (27.3%) |
| No information | -- | 4 (36.4%) |
| TTF1 Positive | -- | 1 (9%) |
| TTF1 Negative | -- | 8 (72.7%) |
| No information | -- | 2 (18%) |
| Comorbidity | ||
| COPD | -- | 1 (9%) |
| Hypertension | -- | 5 (45.5%) |
| Diabetes Mellitus Type II | -- | 4 (36.4%) |
| None | 6 (100%) | 5 (45.5%) |
| Exposure history | ||
| Smokers | -- | 4 (36.4%) |
| Wood smoke | -- | 3 (27.3%) |
| None | 6 (100%) | 4 (35.7%) |
|
RUNX2 ΔCT Median (Range) | p Value |
TALAM1 ΔCT Median (Range) | p Value | |
|---|---|---|---|---|
| Female | 5.92 (1.58–10.85) | 0.1544 | 8.873 (3.724–19.28) | 0.5636 |
| Male | 10.4 (2.6–12.95) | 10.02 (8.747–11.93) | ||
| Age | Spearman r: 0.07323 | 0.8323 | Spearman r: 0.05034 | 0.8861 |
| Histological tumor type | ||||
| Adenocarcinoma | 5.705 (2.6–12.95) | 0.6606 | 10.02 (3.724–18.73) | 0.2121 |
| Squamous cell carcinoma | 6.093 (1.579–6.759) | 7.905 (7.519–19.28) | ||
| TNM stage | ||||
| Kruskal–Wallis multiple comparison analysis (Stages II, III and IV) | -- | 0.3273 | -- | 0.3273 |
| Immunohistochemical markers | ||||
| P63 Positive | 6.093 (1.579–6.759) | 0.8857 | 7.854 (7.519–7.955) | 0.6 |
| P63 Negative | 5.705 (2.6–10.32) | 8.747 (3.724–10.02) | ||
| Exposure history | ||||
| Smokers | 8.368 (4.893–12.95) | 0.5364 | 10.98 (7.519–18.73) | 0.2952 |
| Non-smokers | 5.705 (1.579–10.85) | 8.351 (3.724–13.26) | ||
| Wood smoke | 5.630 (2.6–12.95) | 0.5364 | 9.269 (3.724–11.93) | 0.8857 |
| Non-exposed | 6.418 (1.579–10.85) | 8.988 (7.519–18.73) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bermúdez, G.; Bernal, C.; Otalora, A.; Sanchez, P.; Nardocci, G.; Cañas, A.; Lopez-Kleine, L.; Montecino, M.; Rojas, A. Long Noncoding RNA TALAM1 Is a Transcriptional Target of the RUNX2 Transcription Factor in Lung Adenocarcinoma. Curr. Issues Mol. Biol. 2023, 45, 7075-7086. https://doi.org/10.3390/cimb45090447
Bermúdez G, Bernal C, Otalora A, Sanchez P, Nardocci G, Cañas A, Lopez-Kleine L, Montecino M, Rojas A. Long Noncoding RNA TALAM1 Is a Transcriptional Target of the RUNX2 Transcription Factor in Lung Adenocarcinoma. Current Issues in Molecular Biology. 2023; 45(9):7075-7086. https://doi.org/10.3390/cimb45090447
Chicago/Turabian StyleBermúdez, Gisella, Camila Bernal, Andrea Otalora, Paula Sanchez, Gino Nardocci, Alejandra Cañas, Liliana Lopez-Kleine, Martín Montecino, and Adriana Rojas. 2023. "Long Noncoding RNA TALAM1 Is a Transcriptional Target of the RUNX2 Transcription Factor in Lung Adenocarcinoma" Current Issues in Molecular Biology 45, no. 9: 7075-7086. https://doi.org/10.3390/cimb45090447