
Identifying circRNA–miRNA–mRNA Regulatory Networks in Chemotherapy-Induced Peripheral Neuropathy
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Paclitaxel-Induced Peripheral Neuropathy Model
2.3. Collection of Tissues, Extraction of RNA, Creation of Library, and Sequencing
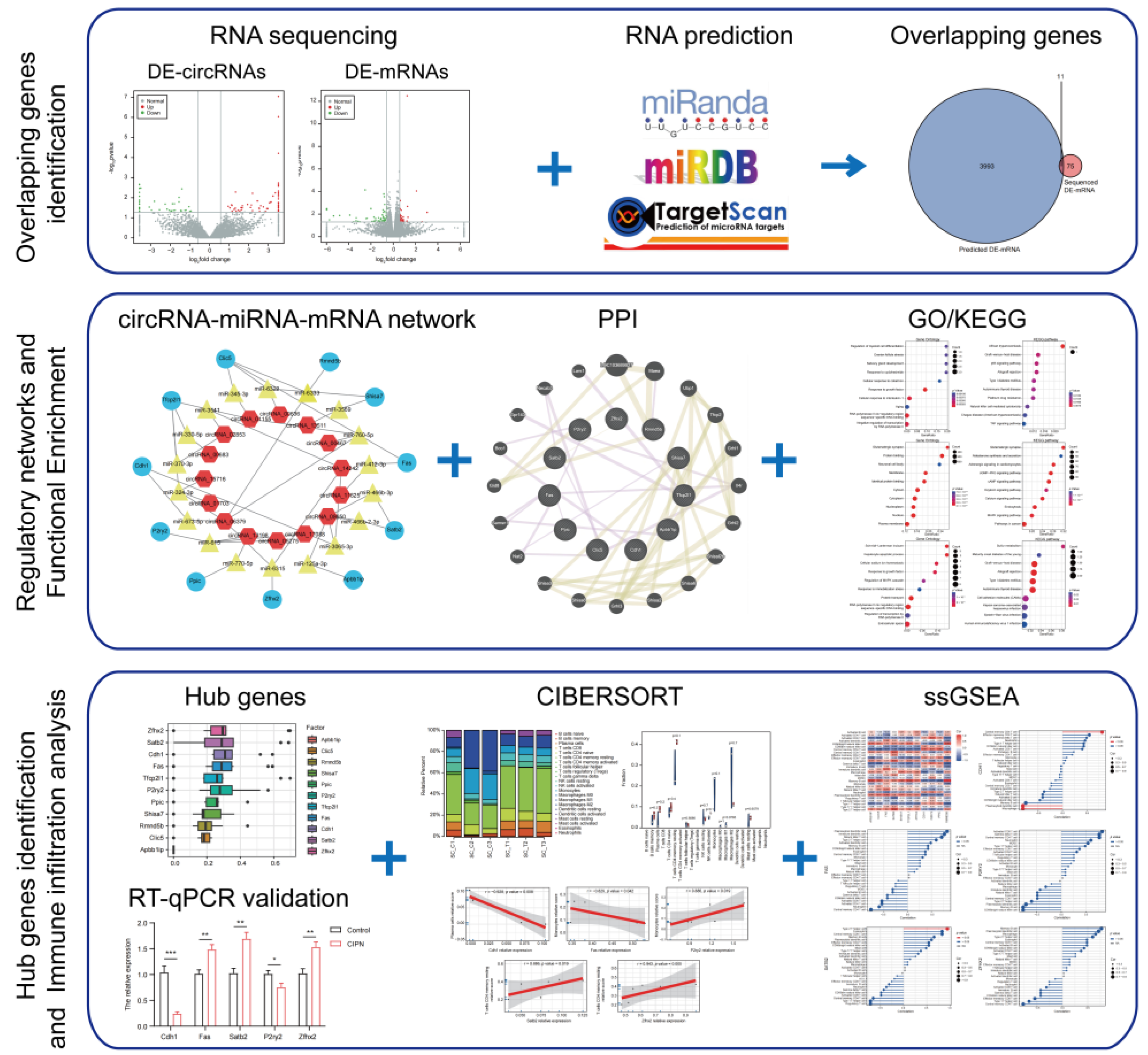
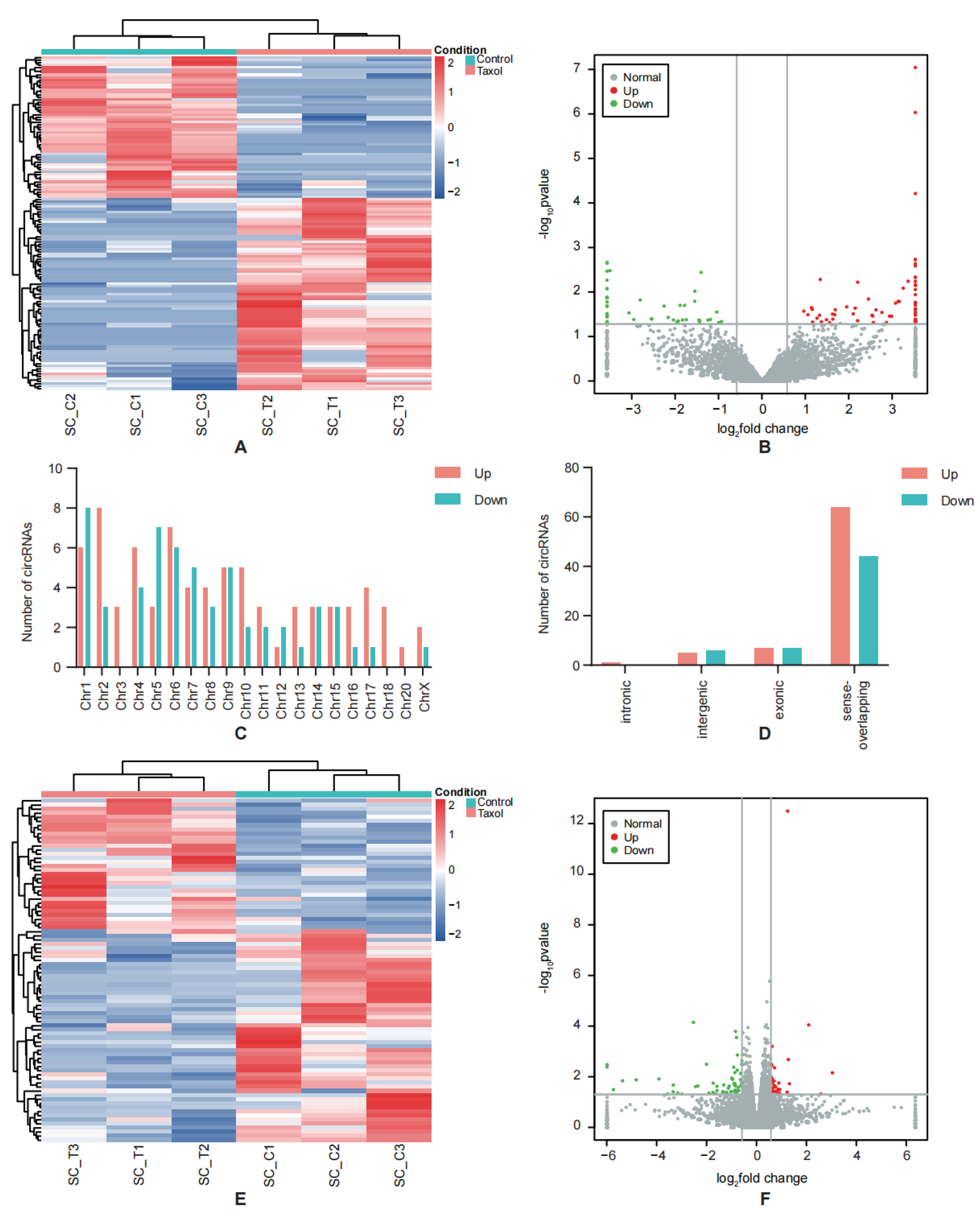
2.4. Differentially Expressed RNAs Identification
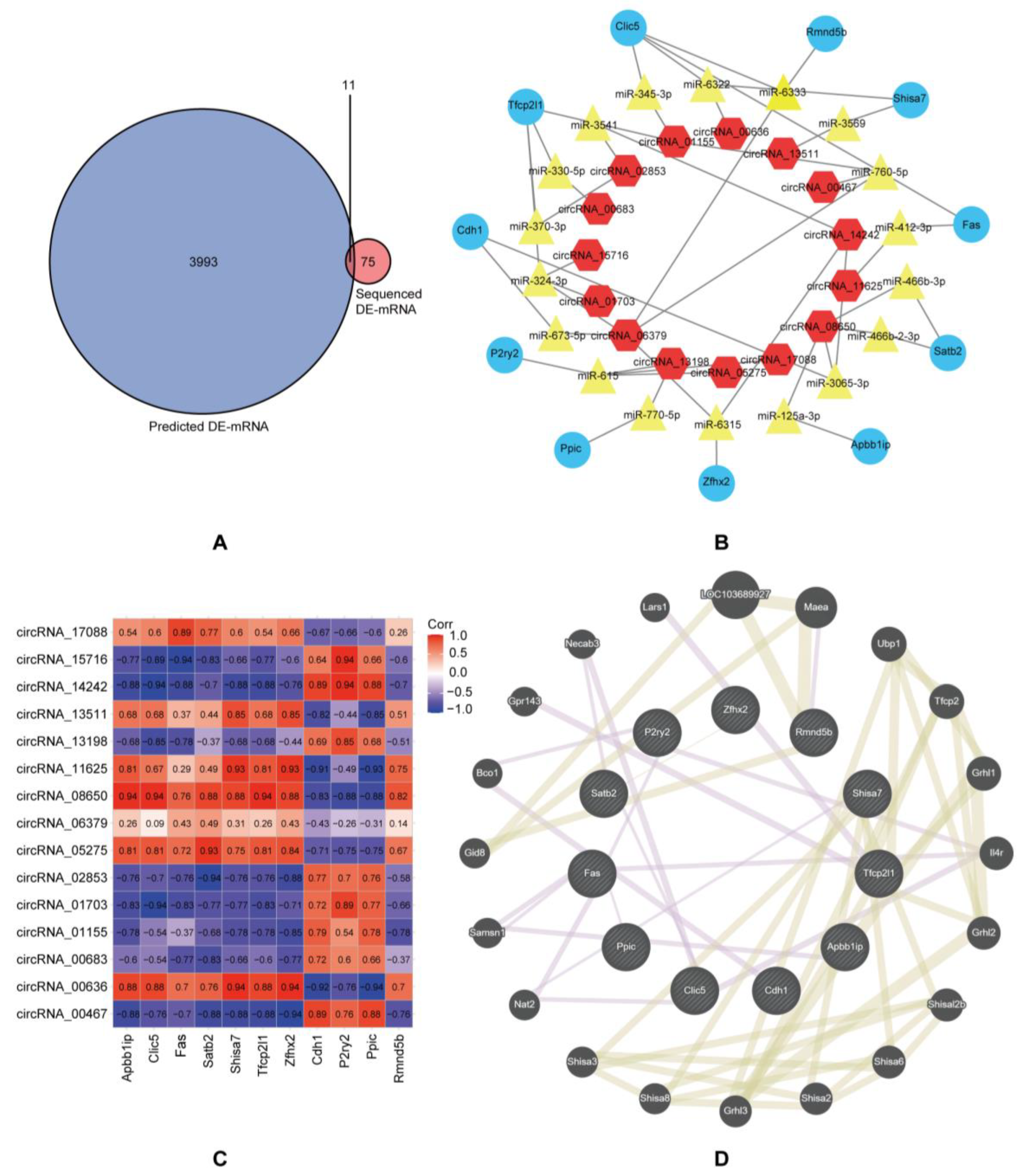
2.5. Establishment of the circRNA-miRNA-mRNA Networks and Protein–Protein Interaction (PPI) Network
2.6. Functional Enrichment Pathway Analysis
2.7. Tissue-Most Expressed Gene Analysis and Hub Genes Identification
2.8. Immune Infiltration Analysis
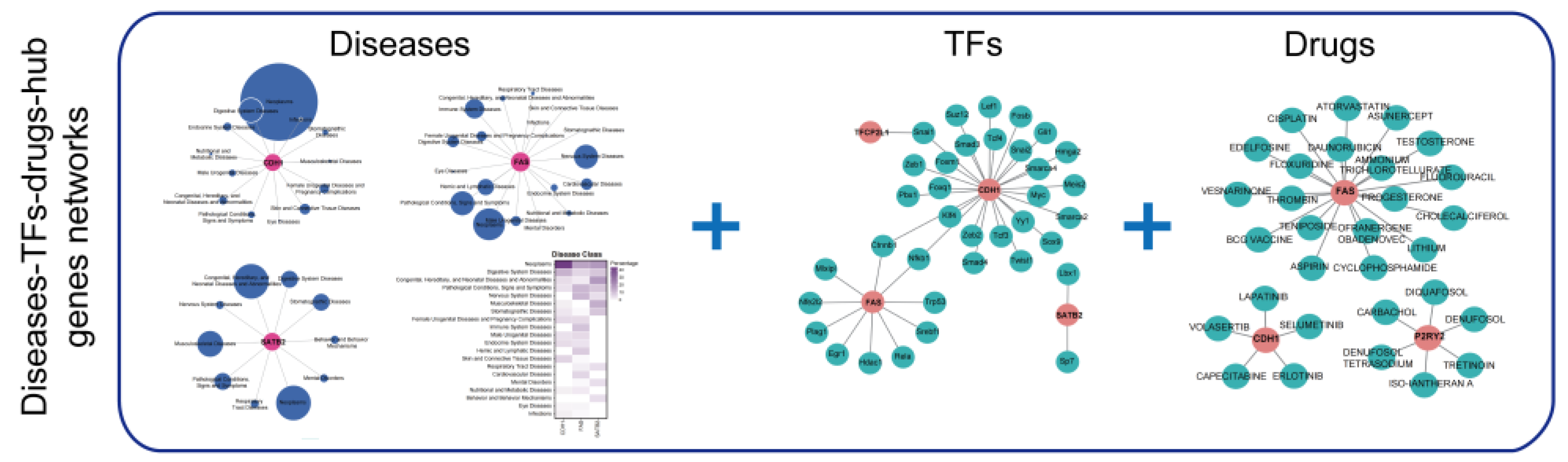
2.9. Diseases, TFs, and Drugs Prediction of Hub Genes
2.10. Validation of Hub Genes Expression
2.11. Statistical Analysis
3. Results
3.1. Profile of Differentially Expressed Genes in CIPN
3.2. Construction of circRNA-miRNA-mRNA Networks and Protein–Protein Interaction Networks
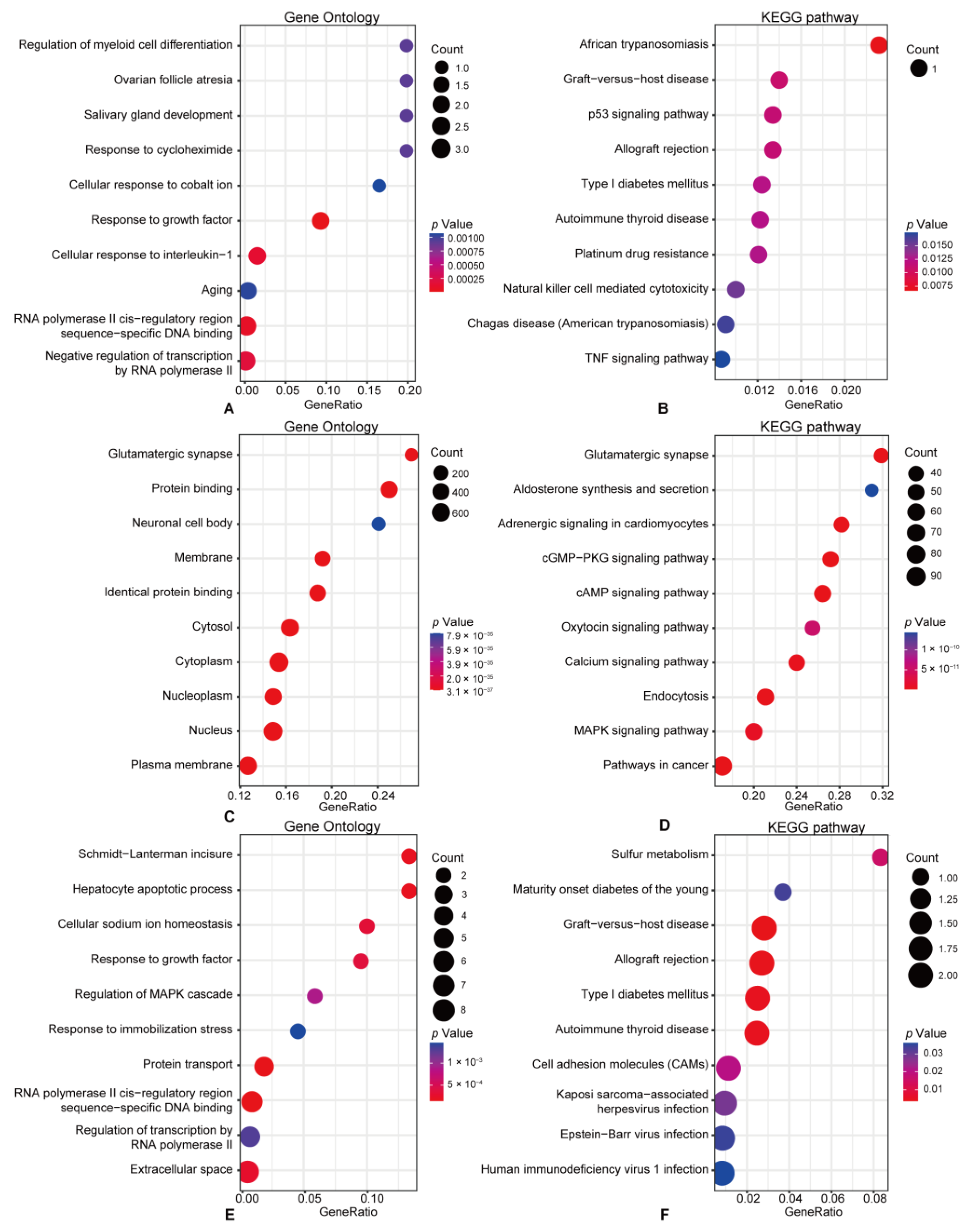
3.3. Functional Enrichment Pathway Analysis
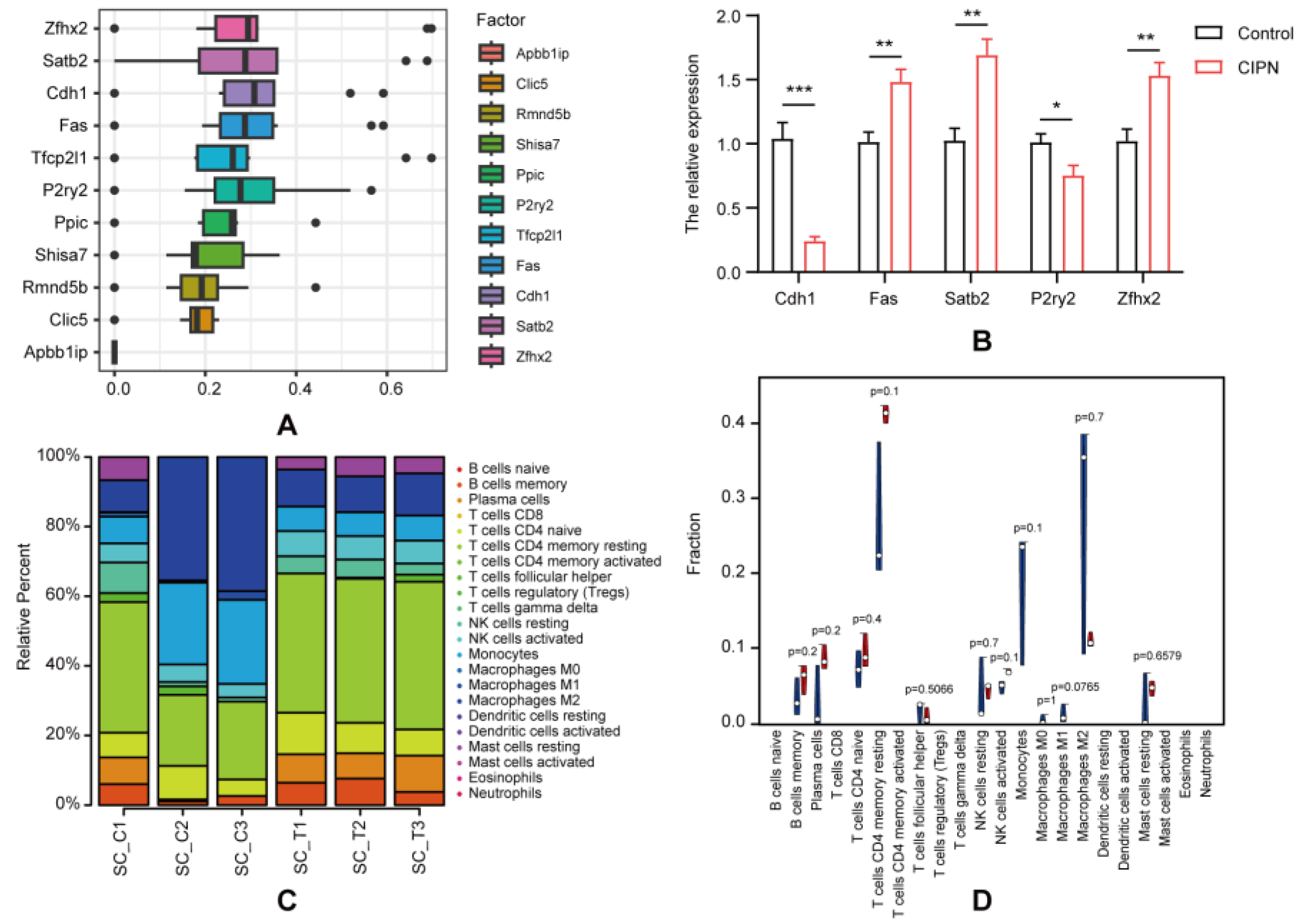
3.4. Hub Genes Identification and Validation
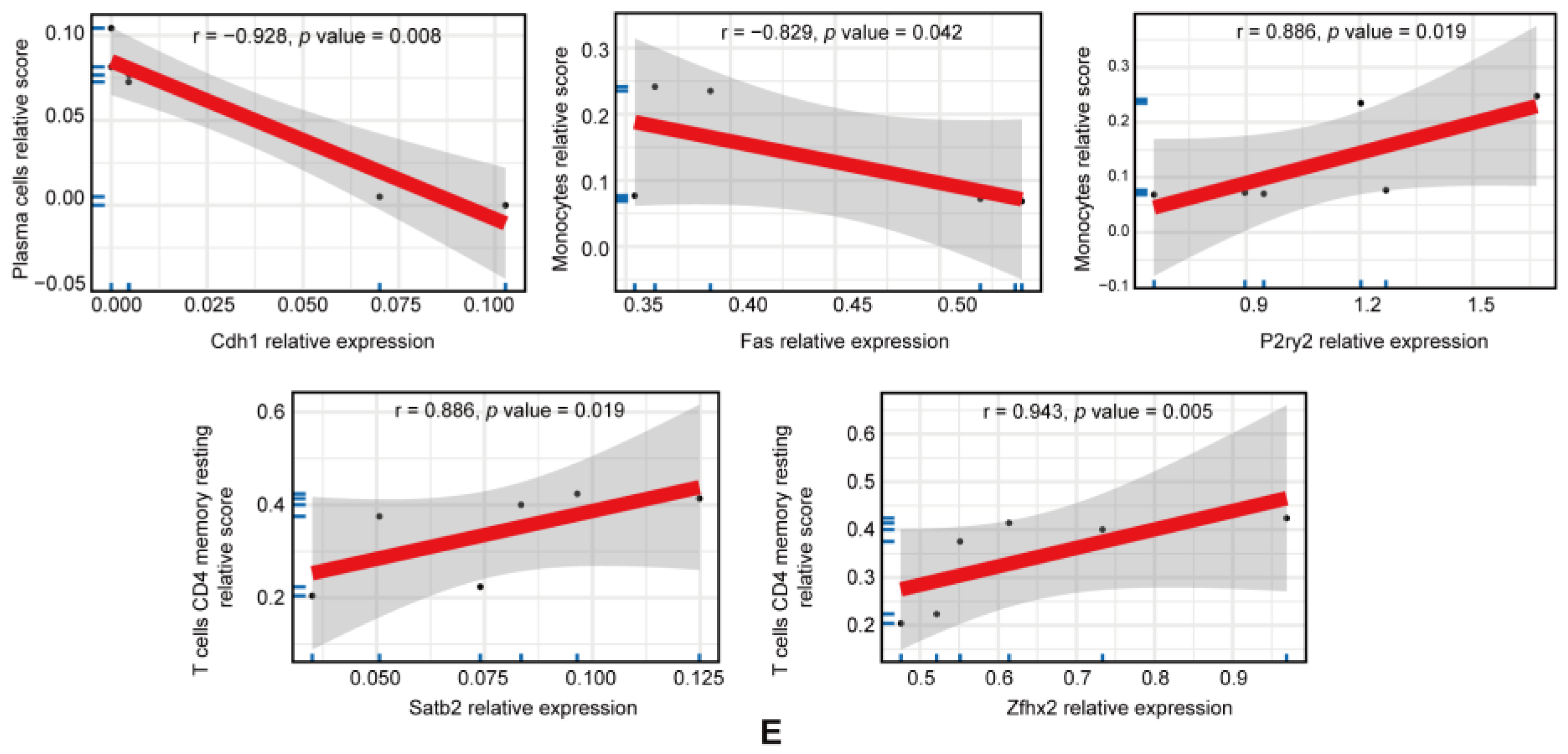
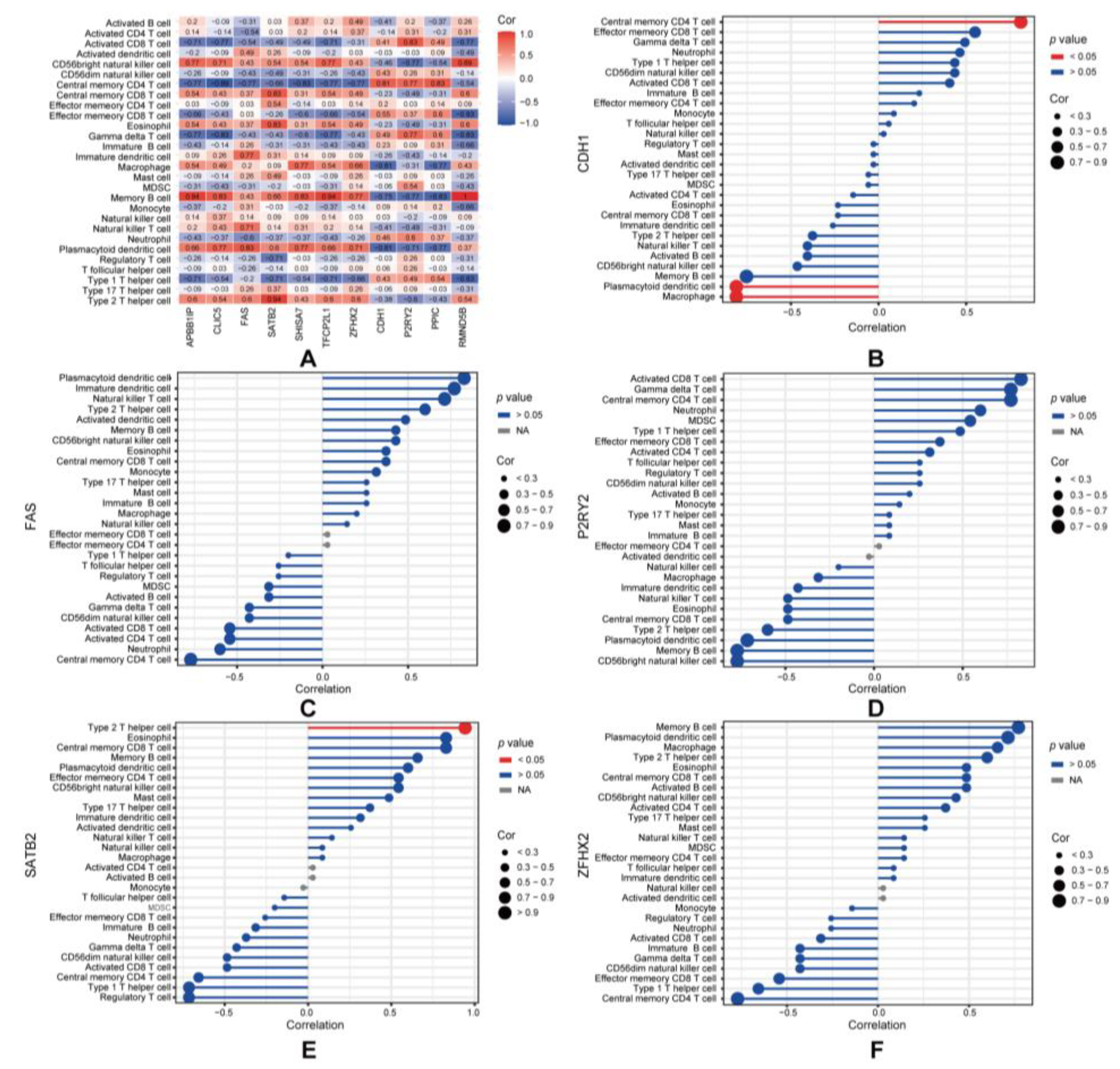
3.5. Immune Infiltration Analysis
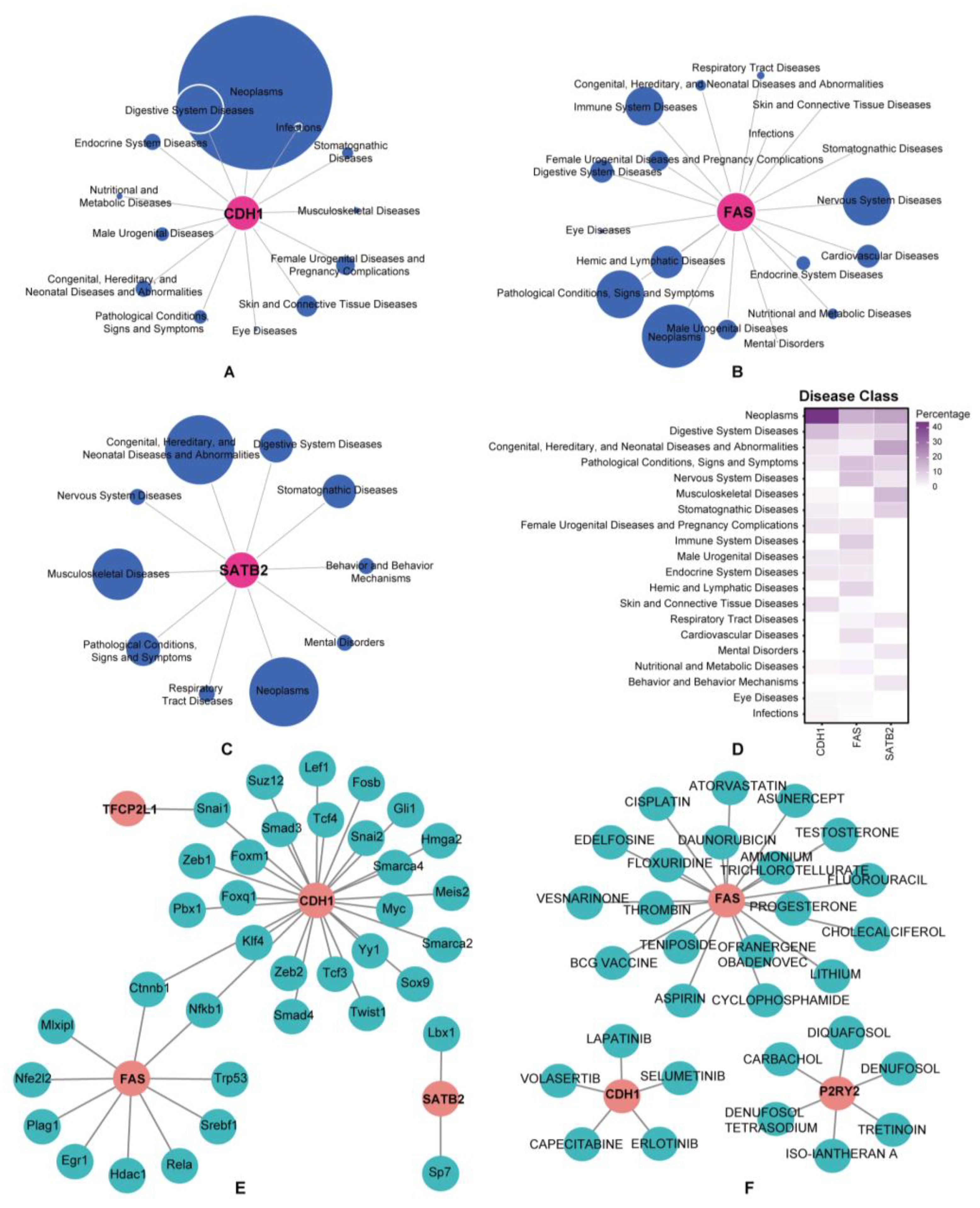
3.6. Prediction of Associated Diseases, Target Transcription Factors, and Drugs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carozzi, V.A.; Canta, A.; Chiorazzi, A. Chemotherapy-induced peripheral neuropathy: What do we know about mechanisms? Neurosci. Lett. 2015, 596, 90–107. [Google Scholar] [CrossRef]
- Park, S.B.; Goldstein, D.; Krishnan, A.V.; Lin, C.S.; Friedlander, M.L.; Cassidy, J.; Koltzenburg, M.; Kiernan, M.C. Chemotherapy-induced peripheral neurotoxicity: A critical analysis. CA Cancer J. Clin. 2013, 63, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Pike, C.T.; Birnbaum, H.G.; Muehlenbein, C.E.; Pohl, G.M.; Natale, R.B. Healthcare costs and workloss burden of patients with chemotherapy-associated peripheral neuropathy in breast, ovarian, head and neck, and nonsmall cell lung cancer. Chemother. Res. Pract. 2012, 2012, 913848. [Google Scholar] [CrossRef] [PubMed]
- Staff, N.P.; Grisold, A.; Grisold, W.; Windebank, A.J. Chemotherapy-induced peripheral neuropathy: A current review. Ann. Neurol. 2017, 81, 772–781. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, Y.; Zhou, S.; Dain, L.; Mei, L.; Zhu, G. Circular RNA: An emerging frontier in RNA therapeutic targets, RNA therapeutics, and mRNA vaccines. J. Control Release 2022, 348, 84–94. [Google Scholar] [CrossRef]
- Liu, C.X.; Chen, L.L. Circular RNAs: Characterization, cellular roles, and applications. Cell 2022, 185, 2016–2034. [Google Scholar] [CrossRef]
- Xu, D.; Ma, X.; Sun, C.; Han, J.; Zhou, C.; Chan, M.T.V.; Wu, W.K.K. Emerging roles of circular RNAs in neuropathic pain. Cell Prolif. 2021, 54, e13139. [Google Scholar] [CrossRef]
- Xin, Y.; Song, X.; Ge, Q. Circular RNA SMEK1 promotes neuropathic pain in rats through targeting microRNA-216a-5p to mediate Thioredoxin Interacting Protein (TXNIP) expression. Bioengineered 2021, 12, 5540–5551. [Google Scholar] [CrossRef]
- Xie, L.; Zhang, M.; Liu, Q.; Wei, R.; Sun, M.; Zhang, Q.; Hao, L.; Xue, Z.; Wang, Q.; Yang, L.; et al. Downregulation of ciRNA-Kat6b in dorsal spinal horn is required for neuropathic pain by regulating Kcnk1 in miRNA-26a-dependent manner. CNS Neurosci. Ther. 2023, in press. [Google Scholar] [CrossRef]
- Mao, Q.; Tian, L.; Wei, J.; Zhou, X.; Cheng, H.; Zhu, X.; Li, X.; Gao, Z.; Zhang, X.; Liang, L. Transcriptome analysis of microRNAs, circRNAs, and mRNAs in the dorsal root ganglia of paclitaxel-induced mice with neuropathic pain. Front. Mol. Neurosci. 2022, 15, 990260. [Google Scholar] [CrossRef] [PubMed]
- Polomano, R.C.; Mannes, A.J.; Clark, U.S.; Bennett, G.J. A painful peripheral neuropathy in the rat produced by the chemotherapeutic drug, paclitaxel. Pain 2001, 94, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Yu, G. Gene Ontology Semantic Similarity Analysis Using GOSemSim. Methods Mol. Biol. 2020, 2117, 207–215. [Google Scholar] [CrossRef]
- Pinero, J.; Ramirez-Anguita, J.M.; Sauch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef]
- Yang, X.; Huang, X.; Lu, W.; Yan, F.; Ye, Y.; Wang, L.; Tang, X.; Zeng, W.; Huang, J.; Xie, J. Transcriptome Profiling of miRNA-mRNA Interactions and Associated Mechanisms in Chemotherapy-Induced Neuropathic Pain. Mol. Neurobiol. 2023, in press. [Google Scholar] [CrossRef]
- Zhang, C.; Gao, R.; Zhou, R.; Chen, H.; Liu, C.; Zhu, T.; Chen, C. The emerging power and promise of non-coding RNAs in chronic pain. Front. Mol. Neurosci. 2022, 15, 1037929. [Google Scholar] [CrossRef]
- Wu, C.; Orozco, C.; Boyer, J.; Leglise, M.; Goodale, J.; Batalov, S.; Hodge, C.L.; Haase, J.; Janes, J.; Huss, J.W., 3rd; et al. BioGPS: An extensible and customizable portal for querying and organizing gene annotation resources. Genome Biol. 2009, 10, R130. [Google Scholar] [CrossRef]
- Li, Y.; Yin, C.; Liu, B.; Nie, H.; Wang, J.; Zeng, D.; Chen, R.; He, X.; Fang, J.; Du, J.; et al. Transcriptome profiling of long noncoding RNAs and mRNAs in spinal cord of a rat model of paclitaxel-induced peripheral neuropathy identifies potential mechanisms mediating neuroinflammation and pain. J. Neuroinflamm. 2021, 18, 48. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.; Wu, Y.; Shen, Y.J.; Xia, J.; Xu, Y.H.; Yue, L.; Sun, L.; Li, X.J.; Ma, Y.; et al. MiR-30d Participates in Vincristine-Induced Neuropathic Pain by Down-Regulating GAD67. Neurochem. Res. 2022, 47, 481–492. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, T.; Li, X.; Wen, C.C.; Yan, X.T.; Peng, C.; Xiao, Y. Circ_0005075 targeting miR-151a-3p promotes neuropathic pain in CCI rats via inducing NOTCH2 expression. Gene 2021, 767, 145079. [Google Scholar] [CrossRef] [PubMed]
- Klein, I.; Lehmann, H.C. Pathomechanisms of Paclitaxel-Induced Peripheral Neuropathy. Toxics 2021, 9, 229. [Google Scholar] [CrossRef]
- Krukowski, K.; Eijkelkamp, N.; Laumet, G.; Hack, C.E.; Li, Y.; Dougherty, P.M.; Heijnen, C.J.; Kavelaars, A. CD8+ T Cells and Endogenous IL-10 Are Required for Resolution of Chemotherapy-Induced Neuropathic Pain. J. Neurosci. 2016, 36, 11074–11083. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; de Carvalho-Barbosa, M.; Kavelaars, A.; Heijnen, C.J.; Albrecht, P.J.; Dougherty, P.M. Dorsal Root Ganglion Infiltration by Macrophages Contributes to Paclitaxel Chemotherapy-Induced Peripheral Neuropathy. J. Pain 2016, 17, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, P.; Ni, Y.; Zhao, J.; Liu, Z. Decreased microRNA-125a-3p contributes to upregulation of p38 MAPK in rat trigeminal ganglions with orofacial inflammatory pain. PLoS ONE 2014, 9, e111594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.B.; Lin, S.Y.; Liu, M.; Liu, C.C.; Ding, H.H.; Sun, Y.; Ma, C.; Guo, R.X.; Lv, Y.Y.; Wu, S.L.; et al. CircAnks1a in the spinal cord regulates hypersensitivity in a rodent model of neuropathic pain. Nat. Commun. 2019, 10, 4119. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ma, L.; Zhang, Z. circ_0013613 Promotes the recovery of spinal cord injury by regulating mir-370-3p/caspase1 to regulate neuronal pyroptosis. Acta Biochim. Pol. 2023, 70, 91–97. [Google Scholar] [CrossRef]
- Matsuyama, S.; Yamamoto, R.; Murakami, K.; Takahashi, N.; Nishi, R.; Ishii, A.; Nio-Kobayashi, J.; Abe, N.; Tanaka, K.; Jiang, J.J.; et al. GM-CSF Promotes the Survival of Peripheral-Derived Myeloid Cells in the Central Nervous System for Pain-Induced Relapse of Neuroinflammation. J. Immunol. 2023, 211, 34–42. [Google Scholar] [CrossRef]
- Hou, W.Y.; Shyu, B.C.; Chen, T.M.; Shieh, J.Y.; Sun, W.Z. Protein synthesis inhibitor cycloheximide dose-dependently decreases formalin-induced c-Fos protein and behavioral hyperalgesia in rats. Neurosci. Lett. 1997, 227, 99–102. [Google Scholar] [CrossRef]
- Di Cesare Mannelli, L.; Micheli, L.; Cervetto, C.; Toti, A.; Lucarini, E.; Parisio, C.; Marcoli, M.; Ghelardini, C. Neuronal alarmin IL-1alpha evokes astrocyte-mediated protective signals: Effectiveness in chemotherapy-induced neuropathic pain. Neurobiol. Dis. 2022, 168, 105716. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Liu, J.; Li, Y.; Liu, J.; Wang, H.; Chai, M.; Dong, Y.; Zhang, Z.; Su, G.; Wang, M. Targeting p53 for neuroinflammation: New therapeutic strategies in ischemic stroke. J. Neurosci. Res. 2023, 101, 1393–1408. [Google Scholar] [CrossRef]
- Kim, H.W.; Wang, S.; Davies, A.J.; Oh, S.B. The therapeutic potential of natural killer cells in neuropathic pain. Trends Neurosci. 2023, 46, 617–627. [Google Scholar] [CrossRef]
- Hayashi, K.; Yi, H.; Zhu, X.; Liu, S.; Gu, J.; Takahashi, K.; Kashiwagi, Y.; Pardo, M.; Kanda, H.; Li, H.; et al. Role of Tumor Necrosis Factor Receptor 1-Reactive Oxygen Species-Caspase 11 Pathway in Neuropathic Pain Mediated by HIV gp120 with Morphine in Rats. Anesth. Analg. 2023, 136, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G.; Colonna, M.; Di Santo, J.P.; McKenzie, A.N. Innate lymphoid cells. Innate lymphoid cells: A new paradigm in immunology. Science 2015, 348, aaa6566. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.J.; Rinaldi, S.; Costigan, M.; Oh, S.B. Cytotoxic Immunity in Peripheral Nerve Injury and Pain. Front. Neurosci. 2020, 14, 142. [Google Scholar] [CrossRef]
- Fiore, N.T.; Debs, S.R.; Hayes, J.P.; Duffy, S.S.; Moalem-Taylor, G. Pain-resolving immune mechanisms in neuropathic pain. Nat. Rev. Neurol. 2023, 19, 199–220. [Google Scholar] [CrossRef]
- Austin, P.J.; Moalem-Taylor, G. The neuro-immune balance in neuropathic pain: Involvement of inflammatory immune cells, immune-like glial cells and cytokines. J. Neuroimmunol. 2010, 229, 26–50. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, Z.; Chen, X. Mechanisms underlying paclitaxel-induced neuropathic pain: Channels, inflammation and immune regulations. Eur. J. Pharmacol. 2022, 933, 175288. [Google Scholar] [CrossRef]
- van Roy, F.; Berx, G. The cell-cell adhesion molecule E-cadherin. Cell Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef]
- Zhao, Q.; Yao, J.; Meng, B.; Shen, S.; Cao, S. Role of the blood-spinal cord barrier: An adheren junction regulation mechanism that promotes chronic postsurgical pain. Biochem. Biophys. Res. Commun. 2023, 660, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, X.; Li, X.; Hu, R.; Yao, W.; Mei, W.; Wan, L.; Gui, L.; Zhang, C. Upregulation of Cdh1 in the trigeminal spinal subnucleus caudalis attenuates trigeminal neuropathic pain via inhibiting GABAergic neuronal apoptosis. Neurochem. Int. 2020, 133, 104613. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Xue, L.; Dong, B.; He, H.; Zhang, W.; Hao, L.; Ma, W.; Zang, G.; Han, C.; Dong, Y. CDH1 overexpression predicts bladder cancer from early stage and inversely correlates with immune infiltration. BMC Urol. 2022, 22, 156. [Google Scholar] [CrossRef] [PubMed]
- Guegan, J.P.; Legembre, P. Nonapoptotic functions of Fas/CD95 in the immune response. FEBS J. 2018, 285, 809–827. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A. Fas (CD95)/FasL (CD178) system during ageing. Cell Biol. Int. 2023, 47, 1295–1313. [Google Scholar] [CrossRef] [PubMed]
- Ethell, D.W.; Buhler, L.A. Fas ligand-mediated apoptosis in degenerative disorders of the brain. J. Clin. Immunol. 2003, 23, 439–446. [Google Scholar] [CrossRef]
- Robins-Steele, S.; Nguyen, D.H.; Fehlings, M.G. The delayed post-injury administration of soluble fas receptor attenuates post-traumatic neural degeneration and enhances functional recovery after traumatic cervical spinal cord injury. J. Neurotrauma 2012, 29, 1586–1599. [Google Scholar] [CrossRef]
- Weisman, G.A.; Wang, M.; Kong, Q.; Chorna, N.E.; Neary, J.T.; Sun, G.Y.; Gonzalez, F.A.; Seye, C.I.; Erb, L. Molecular determinants of P2Y2 nucleotide receptor function: Implications for proliferative and inflammatory pathways in astrocytes. Mol. Neurobiol. 2005, 31, 169–183. [Google Scholar] [CrossRef]
- Malin, S.A.; Davis, B.M.; Koerber, R.H.; Reynolds, I.J.; Albers, K.M.; Molliver, D.C. Thermal nociception and TRPV1 function are attenuated in mice lacking the nucleotide receptor P2Y2. Pain 2008, 138, 484–496. [Google Scholar] [CrossRef]
- Zafar, S.; Luo, Y.; Zhang, L.; Li, C.H.; Khan, A.; Khan, M.I.; Shah, K.; Seo, E.K.; Wang, F.; Khan, S. Daidzein attenuated paclitaxel-induced neuropathic pain via the down-regulation of TRPV1/P2Y and up-regulation of Nrf2/HO-1 signaling. Inflammopharmacology 2023, 31, 1977–1992. [Google Scholar] [CrossRef]
- Zhao, X.; Qu, Z.; Tickner, J.; Xu, J.; Dai, K.; Zhang, X. The role of SATB2 in skeletogenesis and human disease. Cytokine Growth Factor Rev. 2014, 25, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Wang, Y.; Wang, Q.; Qian, Y.; Jiang, Y.; Dong, X.; Chen, H.; Chen, X.; Liu, X.; Yu, S.; et al. In the developing cerebral cortex: Axonogenesis, synapse formation, and synaptic plasticity are regulated by SATB2 target genes. Pediatr. Res. 2023, 93, 1519–1527. [Google Scholar] [CrossRef]
- Bissell, S.; Oliver, C.; Moss, J.; Heald, M.; Waite, J.; Crawford, H.; Kothari, V.; Rumbellow, L.; Walters, G.; Richards, C. The behavioural phenotype of SATB2-associated syndrome: A within-group and cross-syndrome analysis. J. Neurodev. Disord. 2022, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.M.; Matsuyama, A.; Okorokov, A.L.; Santana-Varela, S.; Bras, J.T.; Aloisi, A.M.; Emery, E.C.; Bogdanov, Y.D.; Follenfant, M.; Gossage, S.J.; et al. A novel human pain insensitivity disorder caused by a point mutation in ZFHX2. Brain 2018, 141, 365–376. [Google Scholar] [CrossRef]
- Komine, Y.; Takao, K.; Miyakawa, T.; Yamamori, T. Behavioral abnormalities observed in Zfhx2-deficient mice. PLoS ONE 2012, 7, e53114. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.; Bennett, D.L. Neuroinflammation and the generation of neuropathic pain. Br. J. Anaesth. 2013, 111, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Malcangio, M. Role of the immune system in neuropathic pain. Scand. J. Pain 2019, 20, 33–37. [Google Scholar] [CrossRef]
- Gao, Y.; Mei, C.; Chen, P.; Chen, X. The contribution of neuro-immune crosstalk to pain in the peripheral nervous system and the spinal cord. Int. Immunopharmacol. 2022, 107, 108700. [Google Scholar] [CrossRef]
- Wang, Z.; Song, K.; Zhao, W.; Zhao, Z. Dendritic cells in tumor microenvironment promoted the neuropathic pain via paracrine inflammatory and growth factors. Bioengineered 2020, 11, 661–678. [Google Scholar] [CrossRef]
- Shastri, A.; Al Aiyan, A.; Kishore, U.; Farrugia, M.E. Immune-Mediated Neuropathies: Pathophysiology and Management. Int. J. Mol. Sci. 2023, 24, 7288. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [PubMed]
- Grace, P.M.; Tawfik, V.L.; Svensson, C.I.; Burton, M.D.; Loggia, M.L.; Hutchinson, M.R. The Neuroimmunology of Chronic Pain: From Rodents to Humans. J. Neurosci. 2021, 41, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Physiological Basis for Using Vitamin D to Improve Health. Biomedicines 2023, 11, 1542. [Google Scholar] [CrossRef] [PubMed]
- Golan, E.; Haggiag, I.; Os, P.; Bernheim, J. Calcium, parathyroid hormone, and vitamin D: Major determinants of chronic pain in hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2009, 4, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Poisbeau, P.; Aouad, M.; Gazzo, G.; Lacaud, A.; Kemmel, V.; Landel, V.; Lelievre, V.; Feron, F. Cholecalciferol (Vitamin D(3)) Reduces Rat Neuropathic Pain by Modulating Opioid Signaling. Mol. Neurobiol. 2019, 56, 7208–7221. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. The ErbB/HER family of protein-tyrosine kinases and cancer. Pharmacol. Res. 2014, 79, 34–74. [Google Scholar] [CrossRef] [PubMed]
- Kersten, C.; Cameron, M.G.; Mjaland, S. Epithelial growth factor receptor (EGFR)-inhibition for relief of neuropathic pain—A case series. Scand. J. Pain 2013, 4, 3–7. [Google Scholar] [CrossRef]
- Noble, S.; Wagstaff, A.J. Tretinoin. A review of its pharmacological properties and clinical efficacy in the topical treatment of photodamaged skin. Drugs Aging 1995, 6, 479–496. [Google Scholar] [CrossRef]
- Palencia, G.; Hernandez-Pedro, N.; Saavedra-Perez, D.; Pena-Curiel, O.; Ortiz-Plata, A.; Ordonez, G.; Flores-Estrada, D.; Sotelo, J.; Arrieta, O. Retinoic acid reduces solvent-induced neuropathy and promotes neural regeneration in mice. J. Neurosci. Res. 2014, 92, 1062–1070. [Google Scholar] [CrossRef]
- Farazuddin, M.; Ludka, N.; Friesen, L.; Landers, J.J.; O’Konek, J.J.; Kim, C.H.; Baker, J.R. Retinoic Acid Signaling Is Required for Dendritic Cell Maturation and the Induction of T Cell Immunity. Immunohorizons 2023, 7, 480–492. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| circRNA | Log2FC | p-Value | Regulation | Chrom | Strand | circRNA_Type | Gene Symbol |
|---|---|---|---|---|---|---|---|
| circRNA_08074 | 14.139014 | 0.002317 | Up | Chr2 | - | sense-overlapping | Map3k5 |
| circRNA_10981 | 13.687971 | 0.000000 | Up | Chr4 | - | sense-overlapping | Zfp40 |
| circRNA_17435 | 13.418825 | 0.000001 | Up | ChrX | + | sense-overlapping | |
| circRNA_13195 | 13.364652 | 0.006881 | Up | Chr6 | - | sense-overlapping | Brsk2 |
| circRNA_10018 | 12.959353 | 0.000062 | Up | Chr3 | + | sense-overlapping | Jak2 |
| circRNA_12266 | 12.786437 | 0.044086 | Up | Chr5 | - | exonic | Pcgf5 |
| circRNA_08650 | 12.434478 | 0.005765 | Up | Chr2 | + | intergenic | Pwwp2a |
| circRNA_08056 | 12.433880 | 0.001857 | Up | Chr2 | + | sense-overlapping | Rabep1 |
| circRNA_03312 | 12.400673 | 0.002614 | Up | Chr11 | - | sense-overlapping | Acaca |
| circRNA_13652 | 12.391184 | 0.023951 | Up | Chr6 | + | sense-overlapping | Tex2 |
| circRNA_17582 | 12.266018 | 0.004655 | Up | ChrX | - | sense-overlapping | Arsg |
| circRNA_09053 | 12.260412 | 0.004717 | Up | Chr2 | - | exonic | Ttc3 |
| circRNA_16414 | 12.243545 | 0.008860 | Up | Chr9 | - | sense-overlapping | Bbx |
| circRNA_01593 | 12.200506 | 0.023802 | Up | Chr1 | + | sense-overlapping | Opa1 |
| circRNA_05160 | 12.188125 | 0.007071 | Up | Chr15 | - | sense-overlapping | Gtf2i |
| circRNA_08920 | 12.183688 | 0.007139 | Up | Chr2 | - | sense-overlapping | Zcchc2 |
| circRNA_04866 | 12.068021 | 0.011526 | Up | Chr14 | + | sense-overlapping | Dpp10 |
| circRNA_13511 | 12.020117 | 0.045608 | Up | Chr6 | + | sense-overlapping | Cep350 |
| circRNA_07000 | 11.982601 | 0.025790 | Up | Chr18 | + | sense-overlapping | Lin54 |
| circRNA_02033 | 11.981717 | 0.017243 | Up | Chr10 | + | sense-overlapping | Nop14 |
| circRNA | log2FC | p-Value | Regulation | Chrom | Strand | circRNA_Type | Gene Symbol |
|---|---|---|---|---|---|---|---|
| circRNA_14081 | −12.748110 | 0.046107 | Down | Chr6 | + | intergenic | |
| circRNA_04448 | −12.422531 | 0.002173 | Down | Chr14 | + | sense-overlapping | Vom2r66 |
| circRNA_11535 | −12.340272 | 0.003422 | Down | Chr4 | + | sense-overlapping | Cntn6 |
| circRNA_16338 | −12.260065 | 0.030690 | Down | Chr9 | - | intergenic | |
| circRNA_16911 | −12.245977 | 0.005434 | Down | Chr9 | - | exonic | Ncl |
| circRNA_03440 | −12.150955 | 0.008460 | Down | Chr12 | + | sense-overlapping | Arhgef18 |
| circRNA_12821 | −12.150955 | 0.008460 | Down | Chr5 | + | sense-overlapping | Pum1 |
| circRNA_14242 | −12.148621 | 0.016776 | Down | Chr7 | + | intergenic | |
| circRNA_14478 | −12.042250 | 0.013296 | Down | Chr7 | - | sense-overlapping | Trhde |
| circRNA_12787 | −12.041095 | 0.013327 | Down | Chr5 | + | sense-overlapping | Csmd2 |
| circRNA_11207 | −12.040755 | 0.013336 | Down | Chr4 | - | sense-overlapping | Mkrn1 |
| circRNA_16241 | −12.040415 | 0.013345 | Down | Chr8 | + | sense-overlapping | Itga9 |
| circRNA_02853 | −12.039258 | 0.013376 | Down | Chr10 | + | sense-overlapping | Tnrc6c |
| circRNA_00395 | −12.038917 | 0.013386 | Down | Chr1 | - | exonic | Qpctl |
| circRNA_00467 | −12.035918 | 0.013467 | Down | Chr1 | - | sense-overlapping | Gpatch1 |
| circRNA_07892 | −11.928657 | 0.020688 | Down | Chr2 | + | sense-overlapping | Ssbp2 |
| circRNA_05508 | −11.921805 | 0.020954 | Down | Chr15 | + | sense-overlapping | Vwa8 |
| circRNA_04328 | −11.920180 | 0.021017 | Down | Chr13 | + | sense-overlapping | Cdc42bpa |
| circRNA_01155 | −11.912544 | 0.036172 | Down | Chr1 | - | intergenic | |
| circRNA_14402 | −11.912544 | 0.036172 | Down | Chr7 | + | sense-overlapping | Cep83 |
| Gene | Terms | log2FC | p-Value | Change |
|---|---|---|---|---|
| Apbb1ip | Amyloid beta precursor protein binding family B member 1 interacting protein | 0.6285 | 1.87 × 10−2 | Up |
| Clic5 | Chloride intracellular channel 5 | 0.7067 | 2.25 × 10−2 | Up |
| Fas | Fas cell surface death receptor | 0.5995 | 3.08 × 10−2 | Up |
| Satb2 | SATB homeobox 2 | 0.8733 | 3.12 × 10−2 | Up |
| Shisa7 | Shisa family member 7 | 0.6166 | 1.17 × 10−2 | Up |
| Tfcp2l1 | Transcription factor CP2-like 1 | 0.5947 | 2.03 × 10−2 | Up |
| Zfhx2 | Zinc finger homeobox 2 | 0.6381 | 1.38 × 10−2 | Up |
| Cdh1 | Cadherin 1 | −5.3748 | 1.45 × 10−2 | Down |
| P2ry2 | Purinergic receptor P2Y2 | −0.7790 | 5.51 × 10−2 | Down |
| Ppic | Peptidylprolyl isomerase C | −0.8716 | 1.98 × 10−2 | Down |
| Rmnd5b | Required for meiotic nuclear division 5 homolog B | −3.1652 | 4.40 × 10−2 | Down |
| System | Tissue/Cell | Genes |
|---|---|---|
| Hematologic/ Immune | Whole blood, Macrophage bone marrow, Mast cells | Apbb1ip, Fas, P2ry2, Rmnd5b |
| Neurologic | Nucleus accumbens, Dorsal root ganglia | Shisa7, Zfhx2 |
| Digestive | Large intestine | Satb2 |
| Respiratory | Lung | Clic5 |
| Endocrine | Thyroid, Salivary gland | Cdh1, Ppic, Tfcp2l1, |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, F.; Wang, X.; Ye, Q.; Yan, F.; Lu, W.; Xie, J.; Bi, B.; Wang, X. Identifying circRNA–miRNA–mRNA Regulatory Networks in Chemotherapy-Induced Peripheral Neuropathy. Curr. Issues Mol. Biol. 2023, 45, 6804-6822. https://doi.org/10.3390/cimb45080430
Cao F, Wang X, Ye Q, Yan F, Lu W, Xie J, Bi B, Wang X. Identifying circRNA–miRNA–mRNA Regulatory Networks in Chemotherapy-Induced Peripheral Neuropathy. Current Issues in Molecular Biology. 2023; 45(8):6804-6822. https://doi.org/10.3390/cimb45080430
Chicago/Turabian StyleCao, Fei, Xintong Wang, Qingqing Ye, Fang Yan, Weicheng Lu, Jingdun Xie, Bingtian Bi, and Xudong Wang. 2023. "Identifying circRNA–miRNA–mRNA Regulatory Networks in Chemotherapy-Induced Peripheral Neuropathy" Current Issues in Molecular Biology 45, no. 8: 6804-6822. https://doi.org/10.3390/cimb45080430