

In Silico Bioinformatics Analysis on the Role of Long Non-Coding RNAs as Drivers and Gatekeepers of Androgen-Independent Prostate Cancer Using LNCaP and PC-3 Cells
 and
and 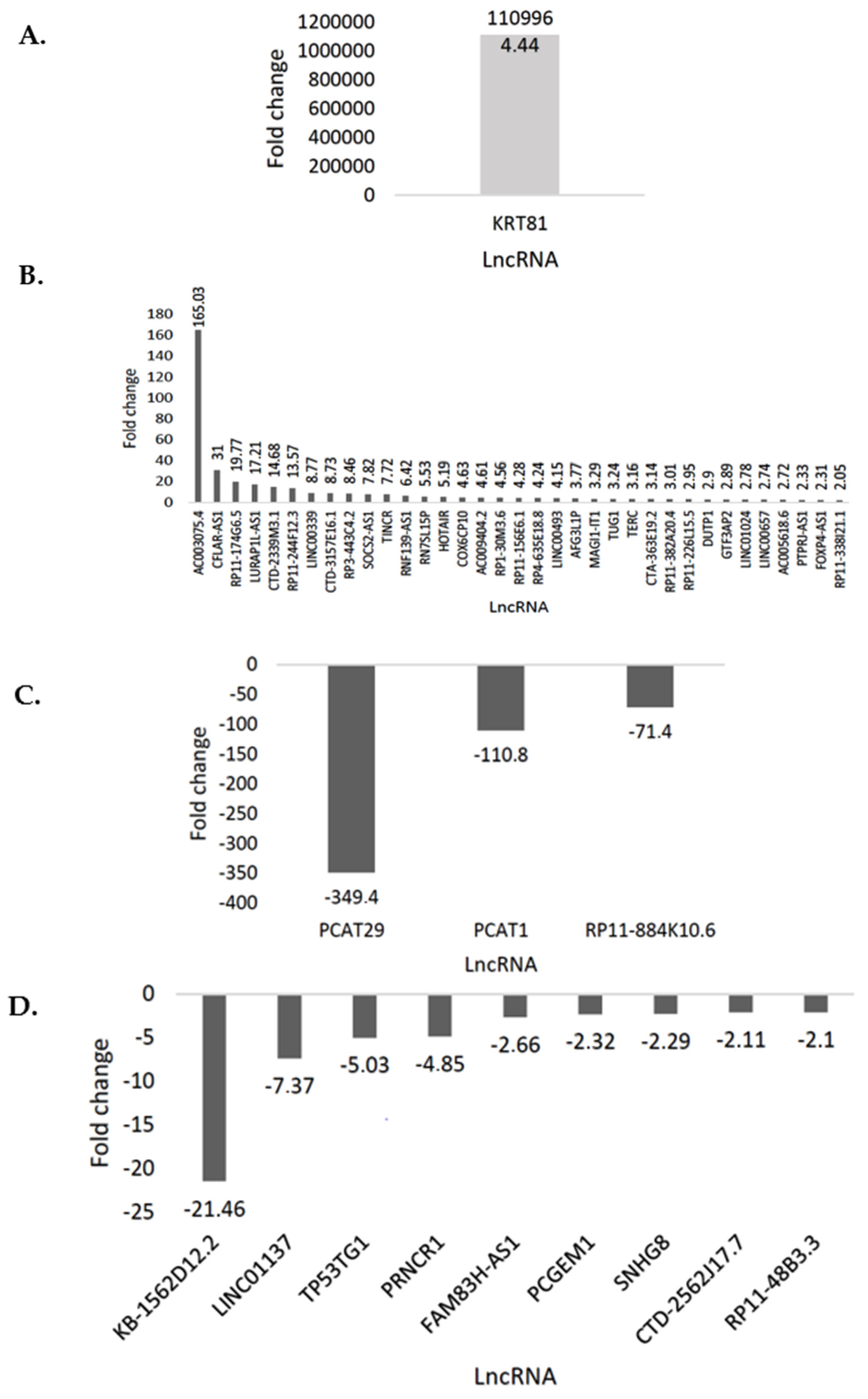{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Extraction and PrimePCR Array Analysis
2.3. LncSEA Database v1.0
2.4. DIANA-miRPath v3.0
2.4.1. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathways
2.4.2. Gene Ontology
2.5. Cytoscape v3.9.1
2.6. Real-Time-Quantitative PCR (RT-qPCR)
2.7. Statistical Analysis
3. Results
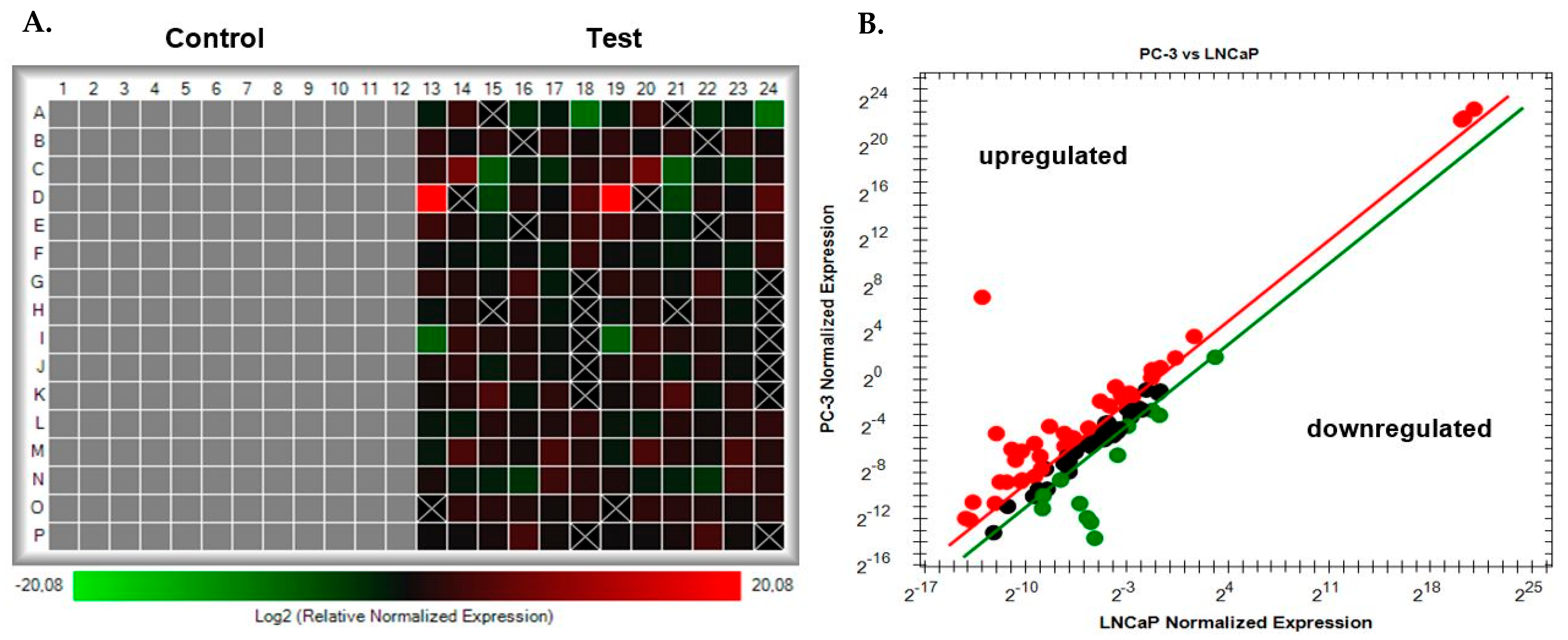
3.1. PrimePCR Array Analysis Using CFX Maestro
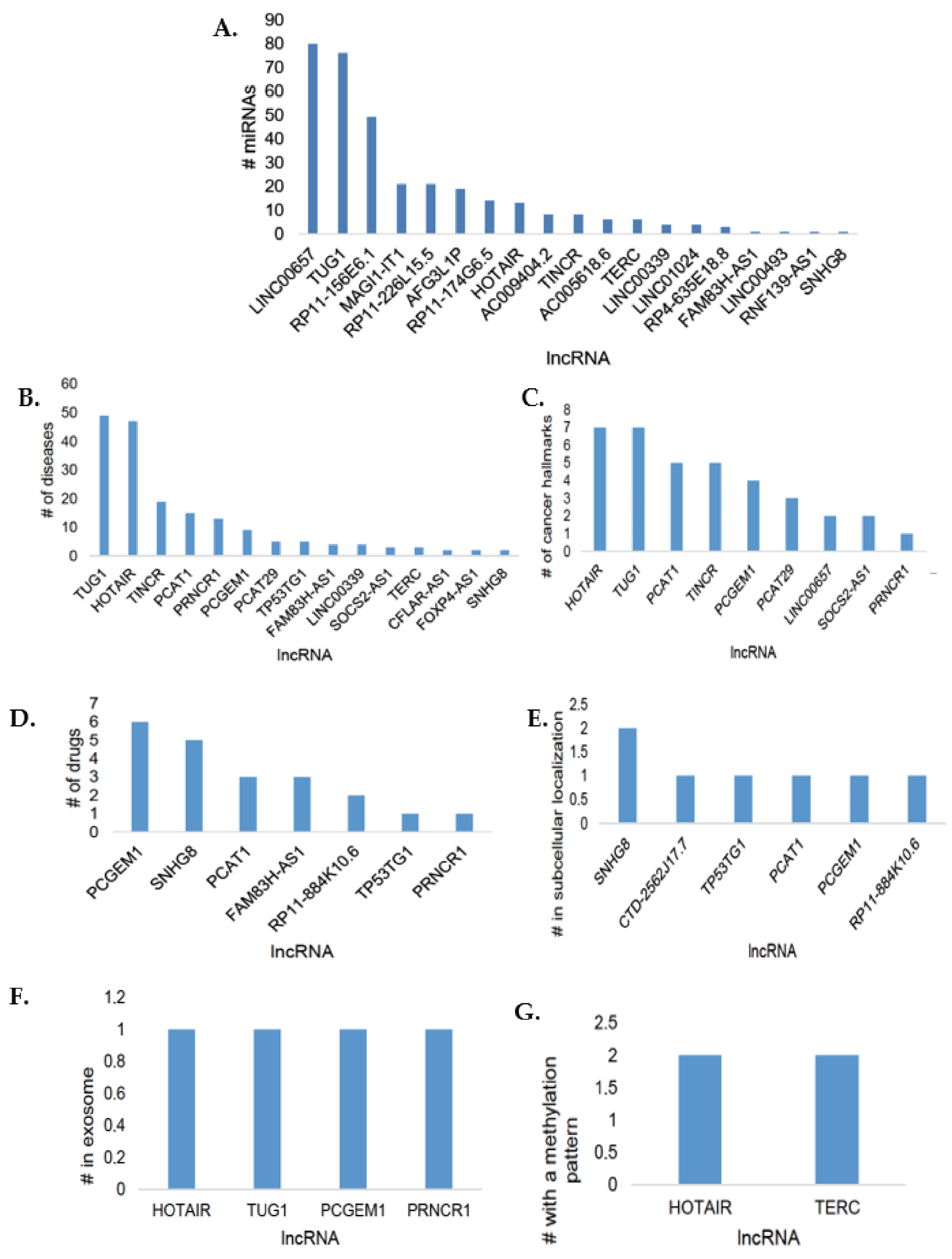
3.2. LncSEA Analysis of Differentially Expressed lncRNAs
3.3. DIANA-miRPath Analysis of the Differentially Expressed lncRNAs and Their Associated miRNA Interactions
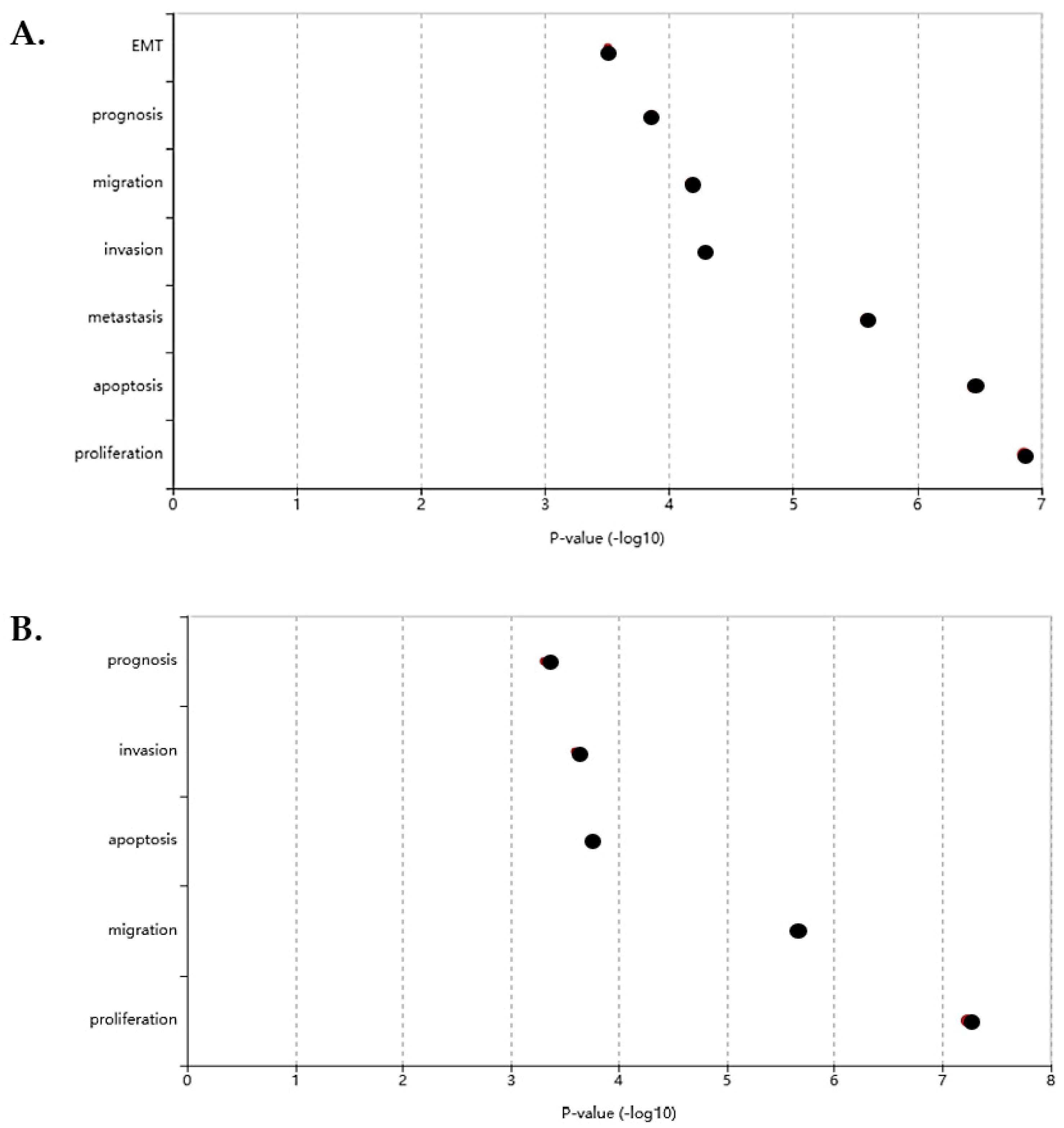
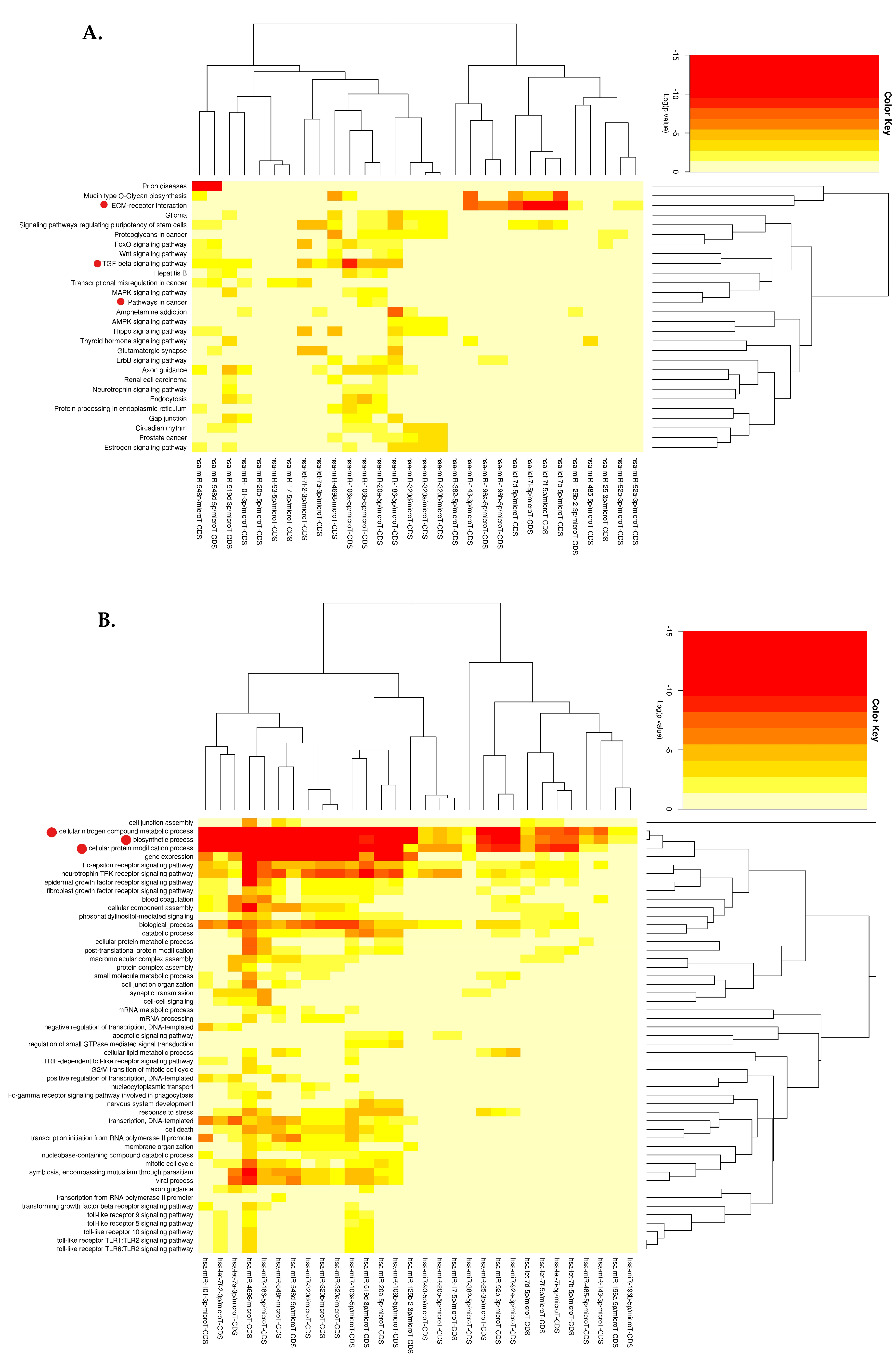
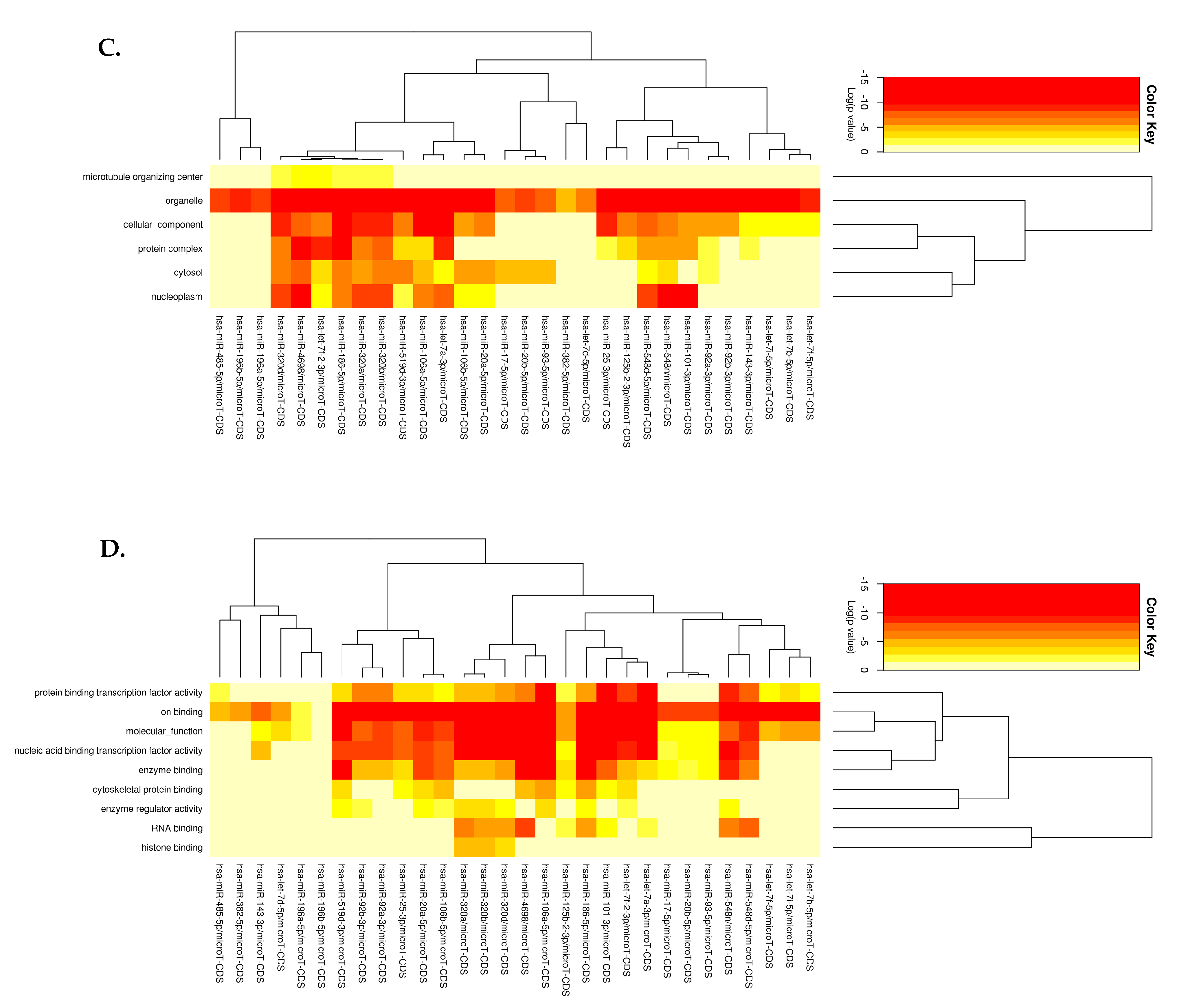
3.3.1. KEGG Pathway Analysis
3.3.2. GO Enrichment Analysis
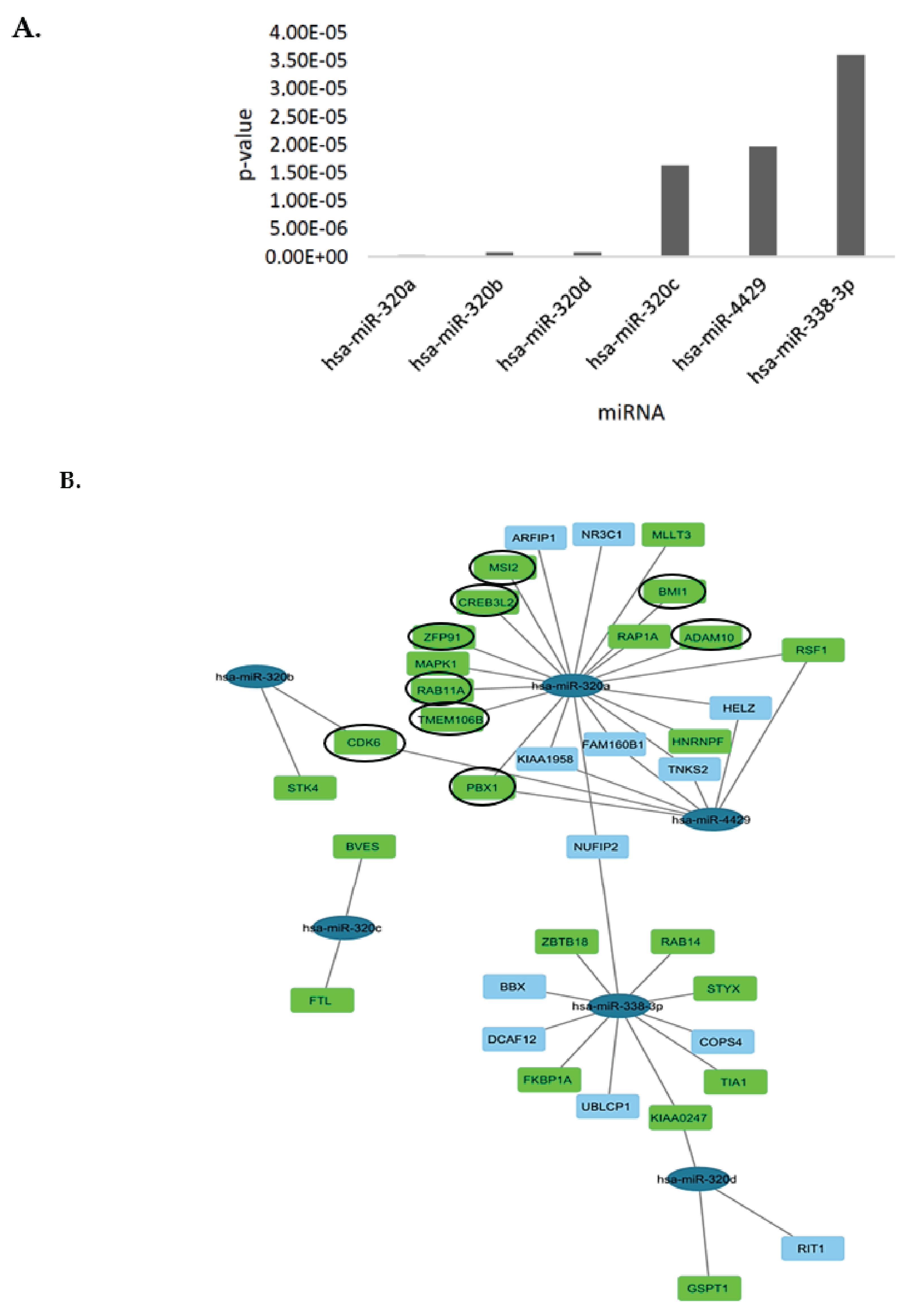
3.4. Bioinformatics Analysis and lncRNA–miRNA-mRNA Competing Endogenous RNA (ceRNA) Network
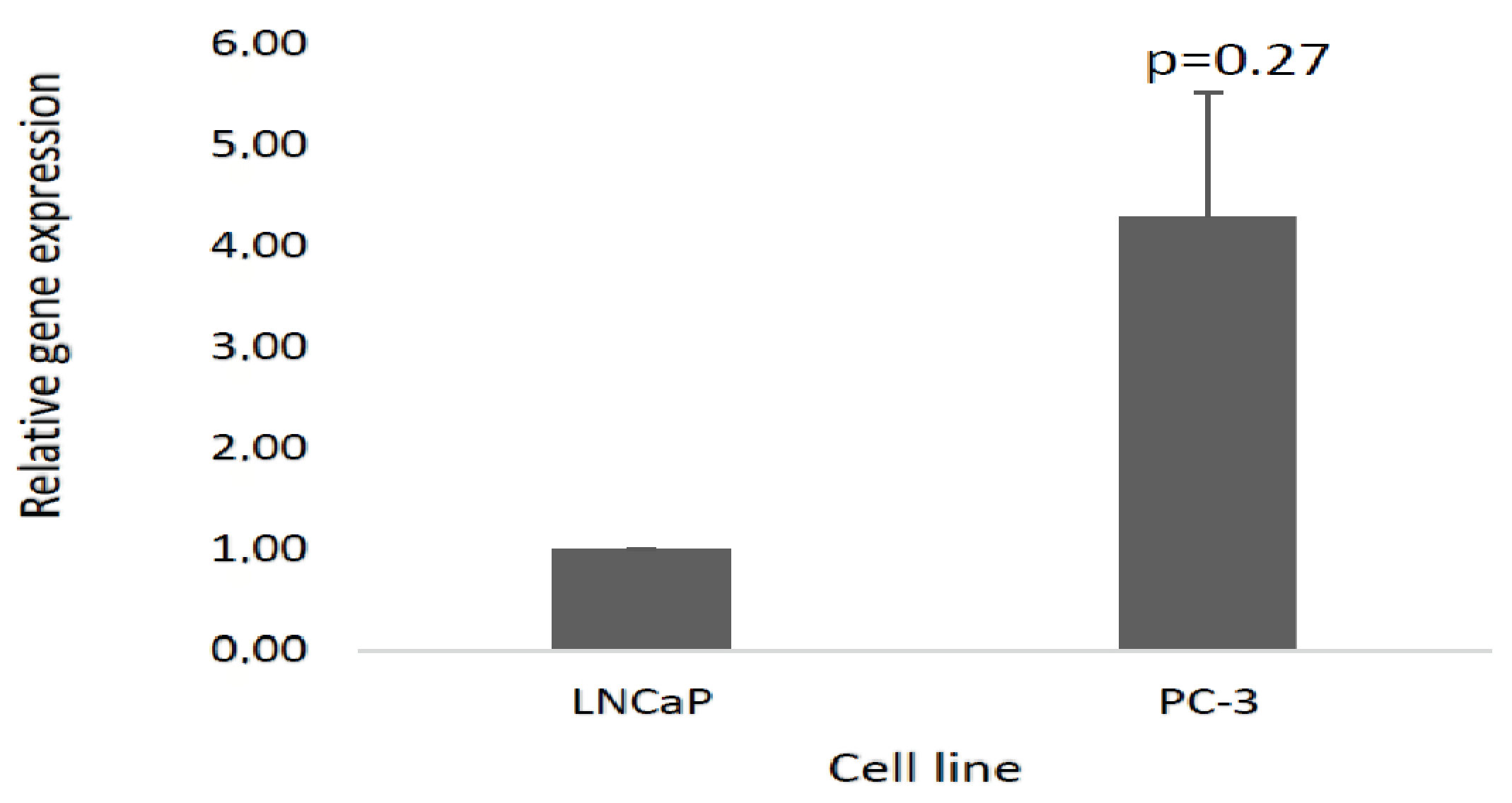
3.5. Validation of the Selected lncRNA Using Real-Time Quantitative PCR (RT-qPCR)
4. Discussion
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, S.; Prajapati, K.S.; Singh, A.K.; Kushwaha, P.P.; Shuaib, M.; Gupta, S. Long non-coding RNA regulating androgen receptor signaling in breast and prostate cancer. Cancer Lett. 2021, 504, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Bolton, E.C.; Jones, J.O. Androgens and androgen receptor signaling in prostate tumorigenesis. J. Mol. Endocrinol. 2015, 54, R15–R29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhao, J.C.; Kim, J.; Fong, K.W.; Yang, Y.A.; Chakravarti, D.; Mo, Y.Y.; Yu, J. LncRNA HOTAIR enhances the androgen-receptor-mediated transcriptional program and drives castration-resistant prostate cancer. Cell Rep. 2015, 13, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, E.S.; Meryet-Figuiere, M.; Sabzalipoor, H.; Kashani, H.H.; Nikzad, H.; Asemi, Z. Dysregulated expression of long noncoding RNAs in gynecologic cancers. Mol. Cancer 2017, 16, 107. [Google Scholar] [CrossRef] [PubMed]
- Walters, K.; Sarsenov, R.; Too, W.S.; Hare, R.K.; Paterson, I.C.; Lambert, D.W.; Brown, S.; Bradford, J.R. Comprehensive functional profiling of long non-coding RNAs through a novel pan-cancer integration approach and modular analysis of their protein-coding gene association networks. BMC Genom. 2019, 20, 454. [Google Scholar] [CrossRef]
- Kashi, K.; Henderson, L.; Bonetti, A.; Carninci, P. Discovery and functional analysis of lncRNAs: Methodologies to investigate an uncharacterized transcriptome. Biochim. Biophys. Acta 2015, 1859, 3–15. [Google Scholar] [CrossRef]
- Petrov, T.L.D.; Riddle, N.C. The evolution of new technologies and methods in clinical epigenetics research. In Epigenetic Biomarkers and Diagnostics; García-Giménez, J.L., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 67–89. [Google Scholar]
- Alimirah, F.; Chen, J.; Basrawala, Z.; Xin, H.; Choubey, D. DU-145 and PC-3 human prostate cancer cell lines express androgen receptor: Implications for the androgen receptor functions and regulation. FEBS Lett. 2006, 580, 2294–2300. [Google Scholar] [CrossRef]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.K.; Liang, C.-Z.; Huang, J. PC3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef]
- Dozmorov, M.G.; Hurst, R.E.; Culkin, D.J.; Kropp, B.P.; Frank, M.B.; Osban, J.; Penning, T.M.; Lin, H.K. Unique patterns of molecular profiling between human prostate cancer LNCaP and PC-3 cells. Prostate 2009, 69, 1077–1090. [Google Scholar] [CrossRef]
- Wang, H.; Wei, X.; Zhang, D.; Li, W.; Hu, Y. LNCaP-AI prostate cancer cell line establishment by Flutamide and androgen-free environment to promote cell adherent. BMC Mol. Cell Biol. 2022, 23, 51. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, J.; Gao, Y.; Li, Y.; Feng, C.; Song, C.; Ning, Z.; Zhou, X.; Zhao, J.; Feng, M.; et al. LncSEA: A platform for long non-coding RNA related sets and enrichment analysis. Nucleic Acids Res 2021, 49, D969–D980. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Cullen, B.R. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA 2007, 13, 313–316. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA-gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Sakurai, K.; Reon, B.J.; Anaya, J.; Dutta, A. The lncRNA DRAIC/PCAT29 locus constitutes a tumor-suppressive nexus. Mol. Cancer Res. 2015, 13, 828. [Google Scholar] [CrossRef]
- Nanashima, N.; Horie, K.; Yamada, T.; Shimizu, T.; Tsuchida, S. Hair keratin KRT81 is expressed in normal and breast cancer cells and contributes to their invasiveness. Oncol. Rep. 2017, 37, 2964–2970. [Google Scholar] [CrossRef]
- Xin, X.; Li, Q.; Fang, J.; Zhao, T. LncRNA HOTAIR: A potential prognostic factor and therapeutic target in human cancers. Front. Oncol. 2021, 11, 679244. [Google Scholar] [CrossRef]
- Cao, Y.; Bryan, T.M.; Reddel, R.R. Increased copy number of the TERT and TERC telomerase subunit genes in cancer cells. Cancer Sci. 2008, 99, 1092–1099. [Google Scholar] [CrossRef]
- Lieb, V.; Weigelt, K.; Scheinost, L.; Fischer, K.; Greither, T.; Marcou, M.; Theil, G.; Klocker, H.; Holzhausen, H.J.; Lai, X.; et al. Serum levels of miR-320 family members are associated with clinical parameters and diagnosis in prostate cancer patients. Oncotarget 2018, 9, 10402–10416. [Google Scholar] [CrossRef]
- Antony, H.; Wiegmans, A.P.; Wei, M.Q.; Chernoff, Y.O.; Khanna, K.K.; Munn, A.L. Potential roles for prions and protein-only inheritance in cancer. Cancer Metastasis Rev. 2012, 31, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Baena-Del Valle, J.A.; Zheng, Q.; Esopi, D.M.; Rubenstein, M.; Hubbard, G.K.; Moncaliano, M.C.; Hruszkewycz, A.; Vaghasia, A.; Yegnasubramanian, S.; Wheelan, S.J.; et al. MYC drives overexpression of telomerase RNA (hTR/TERC) in prostate cancer. J. Pathol. 2018, 244, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liang, Y.; Zhang, W.; Zeng, N.; Tang, S.; Tian, R. KRT81 knockdown inhibits malignant progression of melanoma through regulating interleukin-8. DNA Cell Biol. 2021, 40, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Sun, Y.; Liu, L.; Yu, J.; Ji, J.; Wang, Y.; Yang, J. HOTAIR mediates cisplatin resistance in nasopharyngeal carcinoma by regulating miR-106a-5p/SOX4 axis. Bioengineered 2022, 13, 6567–6578. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.-z.; Li, C.-x.; Zhang, Y.; Weng, M.-z.; Zhang, M.-d.; Qin, Y.-y.; Gong, W.; Quan, Z.-w. Long non-coding RNA HOTAIR, a c-Myc activated driver of malignancy, negatively regulates miRNA-130a in gallbladder cancer. Mol. Cancer 2014, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, R.; Li, L.W.; Liu, X.; Wang, Y.F.; Wang, Q.X.; Zhang, Q. Long non-coding RNA HOTAIR mediates the switching of histone H3 lysine 27 acetylation to methylation to promote epithelial-to-mesenchymal transition in gastric cancer. Int. J. Oncol. 2019, 54, 77–86. [Google Scholar] [CrossRef]
- Da, M.; Zhuang, J.; Zhou, Y.; Qi, Q.; Han, S. Role of long noncoding RNA taurine-upregulated gene 1 in cancers. Mol. Med. 2021, 27, 51. [Google Scholar] [CrossRef]
- Ding, Y.-M.; Chan, E.C.; Liu, L.-C.; Liu, Z.-W.; Wang, Q.; Wang, J.-L.; Cui, X.-P.; Jiang, F.; Guo, X.-S. Long noncoding RNAs: Important participants and potential therapeutic targets for myocardial ischaemia reperfusion injury. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Tang, X.; Wang, Z.; Sun, D.; Wei, X.; Ding, Y. TUG1 promotes prostate cancer progression by acting as a ceRNA of miR-26a. Biosci. Rep. 2018, 38, BSR20180677. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Huang, J.; Mo, J.; Da, X.; Li, Q.; Fan, M.; Lu, H. Exosomal lncRNA TUG1 from cancer-associated fibroblasts promotes liver cancer cell migration, invasion, and glycolysis by regulating the miR-524-5p/SIX1 axis. Cell. Mol. Biol. Lett. 2022, 27, 17. [Google Scholar] [CrossRef]
- Roshani, M.; Baniebrahimi, G.; Mousavi, M.; Zare, N.; Sadeghi, R.; Salarinia, R.; Sheida, A.; Molavizadeh, D.; Sadeghi, S.; Moammer, F.; et al. Exosomal long non-coding RNAs: Novel molecules in gastrointestinal cancers’ progression and diagnosis. Front. Oncol. 2022, 12, 1014949. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.; Wen, C.; Huo, Z.; Wang, W.; Zhan, Q.; Cheng, D.; Chen, H.; Deng, X.; Peng, C.; et al. Long noncoding RNA NORAD, a novel competing endogenous RNA, enhances the hypoxia-induced epithelial-mesenchymal transition to promote metastasis in pancreatic cancer. Mol. Cancer 2017, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Y.; Peng, H.; Zhu, Q.; Wu, Y.X.; Wu, F.; Han, C.R.; Yan, B.; Li, Q.; Xiang, H.G. Silencing the long noncoding RNA NORAD inhibits gastric cancer cell proliferation and invasion by the RhoA/ROCK1 pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3760–3770. [Google Scholar] [CrossRef]
- Chu, L.; Yu, L.; Liu, J.; Song, S.; Yang, H.; Han, F.; Liu, F.; Hu, Y. Long intergenic non-coding LINC00657 regulates tumorigenesis of glioblastoma by acting as a molecular sponge of miR-190a-3p. Aging 2019, 11, 1456–1470. [Google Scholar] [CrossRef]
- Yang, L.; Lin, C.; Jin, C.; Yang, J.C.; Tanasa, B.; Li, W.; Merkurjev, D.; Ohgi, K.A.; Meng, D.; Zhang, J.; et al. lncRNA-dependent mechanisms of androgen-receptor-regulated gene activation programs. Nature 2013, 500, 598–602. [Google Scholar] [CrossRef]
- Srikantan, V.; Zou, Z.; Petrovics, G.; Xu, L.; Augustus, M.; Davis, L.; Livezey, J.R.; Connell, T.; Sesterhenn, I.A.; Yoshino, K.; et al. PCGEM1, a prostate-specific gene, is overexpressed in prostate cancer. Proc. Natl. Acad. Sci. USA 2000, 97, 12216–12221. [Google Scholar] [CrossRef]
- Ehrlich, M. DNA hypermethylation in disease: Mechanisms and clinical relevance. Epigenetics 2019, 14, 1141–1163. [Google Scholar] [CrossRef]
- Wang, J.; Wu, M.; Chang, L.; Jin, Z.; Yang, X.; Li, D.; Wang, J.; Qu, J.; Hou, Q.; Huang, X.; et al. The lncRNA TERC promotes gastric cancer cell proliferation, migration, and invasion by sponging miR-423-5p to regulate SOX12 expression. Ann. Transl. Med. 2022, 10, 963. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, L.; Roebuck, P.; Diao, L.; Liu, L.; Yuan, Y.; Weinstein, J.N.; Liang, H. TANRIC: An interactive open platform to explore the function of lncRNAs in cancer. Cancer Res. 2015, 75, 3728–3737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Wang, J.; Liu, Z.L.; Liu, H.; Cheng, Y.F.; Wang, T. LINC00657/miR-26a-5p/CKS2 ceRNA network promotes the growth of esophageal cancer cells via the MDM2/p53/Bcl2/Bax pathway. Biosci. Rep. 2020, 40, BSR20200525. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Liu, C.; Li, H.; Zhang, L.; Luo, G.; Liang, S.; Lü, M. Research progress on the interactions between long non-coding RNAs and microRNAs in human cancer (Review). Oncol. Lett. 2020, 19, 595–605. [Google Scholar] [CrossRef]
- Liang, Y.; Li, S.; Tang, L. MicroRNA 320, an anti-oncogene target miRNA for cancer therapy. Biomedicines 2021, 9, 591. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Zhu, H.; Zhao, S.; Wang, K.; Zhang, N.; Tian, Y.; Li, Y.; Wang, Y.; Lv, X.; Wei, T.; et al. The novel circCLK3/miR-320a/FoxM1 axis promotes cervical cancer progression. Cell Death Dis. 2019, 10, 950. [Google Scholar] [CrossRef]
- Lv, Q.L.; Du, H.; Liu, Y.L.; Huang, Y.T.; Wang, G.H.; Zhang, X.; Chen, S.H.; Zhou, H.H. Low expression of microRNA-320b correlates with tumorigenesis and unfavorable prognosis in glioma. Oncol. Rep. 2017, 38, 959–966. [Google Scholar] [CrossRef]
- Wang, W.Y.; Lu, W.C. Reduced expression of hsa-miR-338-3p contributes to the development of glioma cells by targeting mitochondrial 3-Oxoacyl-ACP Synthase (OXSM) in glioblastoma (GBM). Onco Targets Ther. 2020, 13, 9513–9523. [Google Scholar] [CrossRef]
- Sun, H.; Fan, G.; Deng, C.; Wu, L. miR-4429 sensitized cervical cancer cells to irradiation by targeting RAD51. J. Cell. Physiol. 2020, 235, 185–193. [Google Scholar] [CrossRef]
- Wang, J.; Xie, S.; Liu, J.; Li, T.; Wang, W.; Xie, Z. MicroRNA-4429 suppresses proliferation of prostate cancer cells by targeting distal-less homeobox 1 and inactivating the Wnt/β-catenin pathway. BMC Urol. 2021, 21, 40. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, H. miR-338-3p targets RAB23 and suppresses tumorigenicity of prostate cancer cells. Am. J. Cancer Res. 2018, 8, 2564–2574. [Google Scholar] [PubMed]
- Zhang, J.; Zhao, X.; Luan, Z.; Wang, A. RAB14 overexpression promotes proliferation and invasion through YAP signaling in non-small cell lung cancers. Onco Targets Ther. 2020, 13, 9269–9280. [Google Scholar] [CrossRef] [PubMed]
- Philip, M.; Chen, T.; Tyagi, S. A survey of current resources to study lncRNA-protein interactions. Non-Coding RNA 2021, 7, 33. [Google Scholar] [CrossRef]
- Ganesan, A.; Coote, M.L.; Barakat, K. Molecular dynamics-driven drug discovery: Leaping forward with confidence. Drug Discov. Today 2017, 22, 249–269. [Google Scholar] [CrossRef] [PubMed]
- De Summa, S.; Palazzo, A.; Caputo, M.; Iacobazzi, R.M.; Pilato, B.; Porcelli, L.; Tommasi, S.; Paradiso, A.V.; Azzariti, A. Long non-coding RNA landscape in prostate cancer molecular subtypes: A feature selection approach. Int. J. Mol. Sci. 2021, 22, 2227. [Google Scholar] [CrossRef]
- Słomka, A.; Kornek, M.; Cho, W.C. Small extracellular vesicles and their involvement in cancer resistance: An up-to-date review. Cells 2022, 11, 2913. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mbeje, M.; Kandhavelu, J.; Penny, C.; Kgoebane-Maseko, M.; Dlamini, Z.; Marima, R. In Silico Bioinformatics Analysis on the Role of Long Non-Coding RNAs as Drivers and Gatekeepers of Androgen-Independent Prostate Cancer Using LNCaP and PC-3 Cells. Curr. Issues Mol. Biol. 2023, 45, 7257-7274. https://doi.org/10.3390/cimb45090459
Mbeje M, Kandhavelu J, Penny C, Kgoebane-Maseko M, Dlamini Z, Marima R. In Silico Bioinformatics Analysis on the Role of Long Non-Coding RNAs as Drivers and Gatekeepers of Androgen-Independent Prostate Cancer Using LNCaP and PC-3 Cells. Current Issues in Molecular Biology. 2023; 45(9):7257-7274. https://doi.org/10.3390/cimb45090459
Chicago/Turabian StyleMbeje, Mandisa, Jeyalakshmi Kandhavelu, Clement Penny, Mmamoletla Kgoebane-Maseko, Zodwa Dlamini, and Rahaba Marima. 2023. "In Silico Bioinformatics Analysis on the Role of Long Non-Coding RNAs as Drivers and Gatekeepers of Androgen-Independent Prostate Cancer Using LNCaP and PC-3 Cells" Current Issues in Molecular Biology 45, no. 9: 7257-7274. https://doi.org/10.3390/cimb45090459