
Molecular Detection of Nosema spp. in Three Eco Regions of Slovakia
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Sample Preparation for Microscopic Examination of Nosema spp.
2.3. PCR-Based Diagnostics of Nosema spp.
2.4. Reaction Mixture Preparation
2.5. Statistical Analysis
3. Results
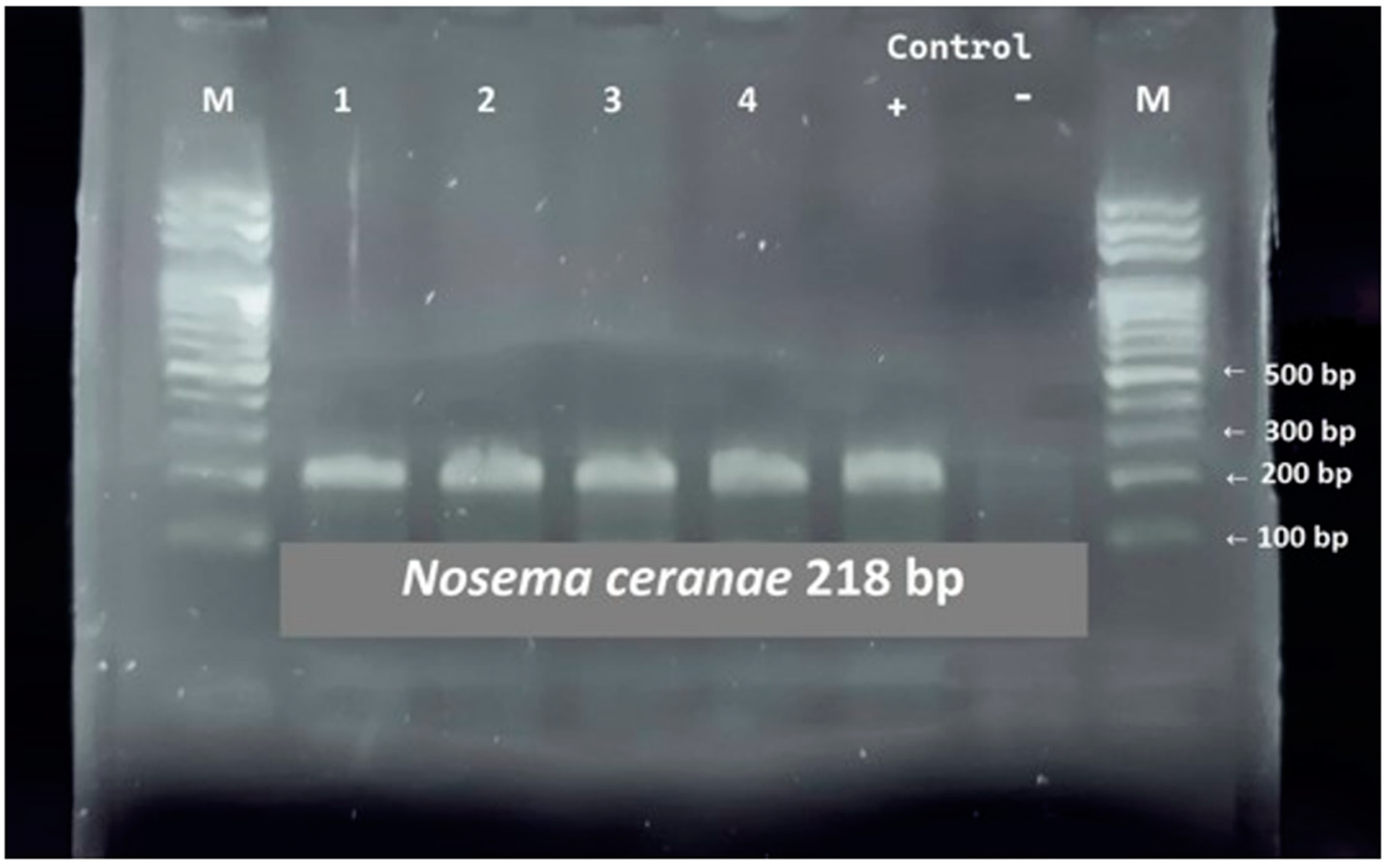
3.1. PCR Diagnostics of Nosema spp.
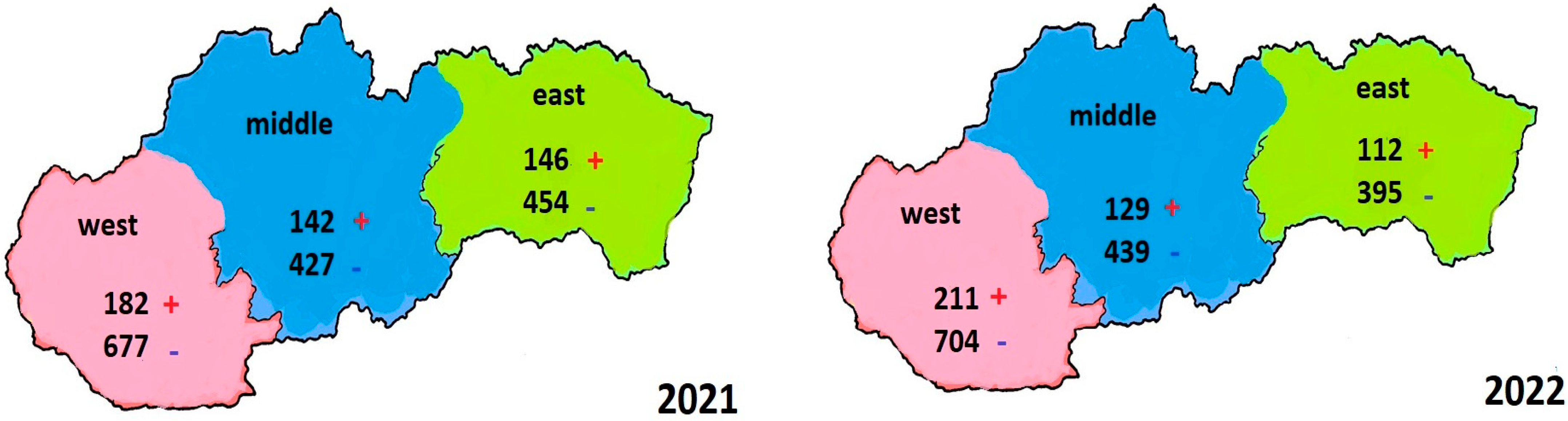
3.2. Evaluation of the Results of Examined Samples in 2021 in Three Ecoregions of Slovakia
3.3. Evaluation of the Results of Examined Samples in 2022 in Three Ecoregions of Slovakia
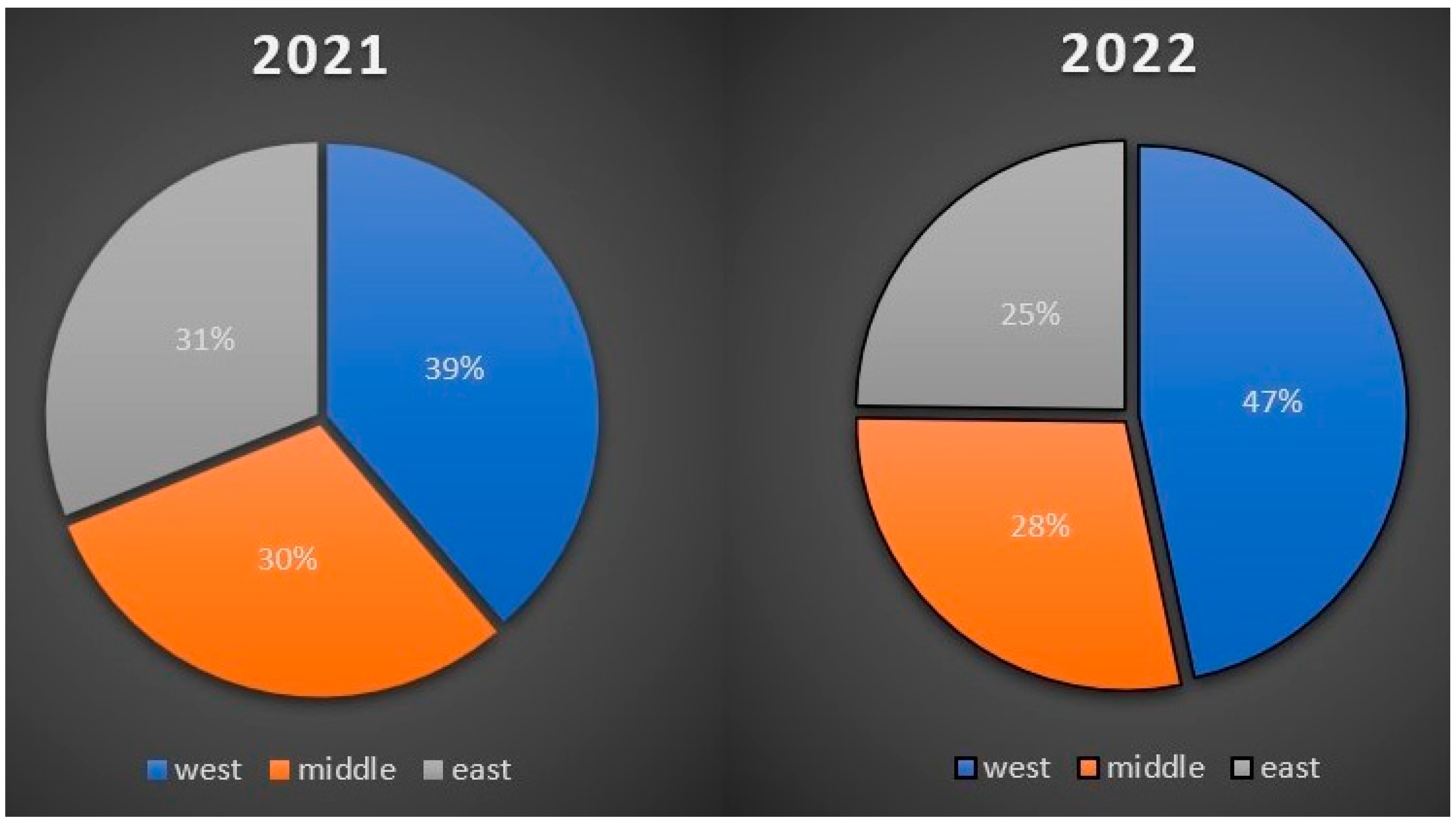
3.4. Comparison of the Number of Positive Cases in Three Slovak Ecoregions from Samples Collected in 2021 and 2022
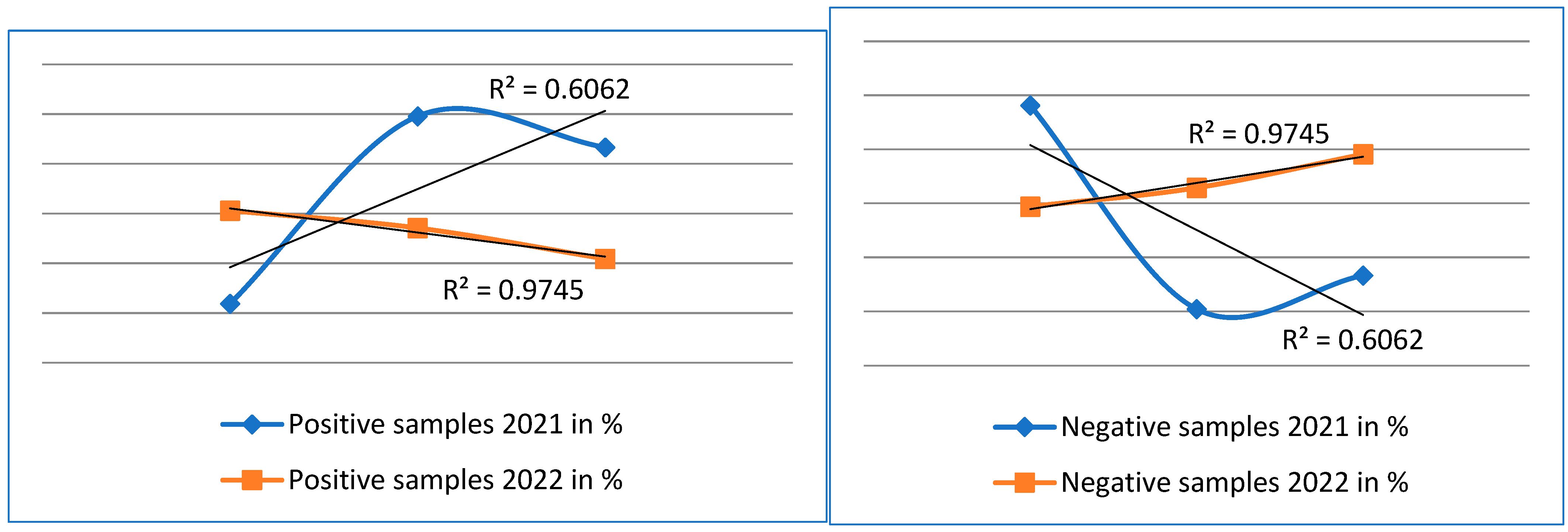
3.5. Evaluation of Statistical Analysis of Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethics Statement
References
- Dong, Z.; Wu, Q.; Long, J.; Lu, B.; Zheng, N.; Hu, C.; Chen, P.; Hu, N.; Lu, C.; Pan, M. Silver nanoparticles are effective in controlling microsporidia. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 125, 112106. [Google Scholar] [CrossRef]
- Urbieta-Margo, A.; Higes, M.; Meana, A.; Goméz-Moracho, T.; Rodríguez- García, C.; Barrios, L.; Martín-Hernández, R. The levels of natural Nosema spp. infection in Apis mellifera iberiensis brood stages. Int. J. Parasitol. 2019, 49, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, A.P.; Lazzarini, K.R.G.; Freitas, N.H.A.; Message, D.; Bitondi, M.M.G.; Simoes, Z.; Teixeira, É. Immunity and physiological changes in adult honey bees (Apis mellifera) infected with Nosema ceranae: The natural colony environment. J. Insect Physiol. 2021, 131, 104237. [Google Scholar] [CrossRef]
- Murareanu, B.M.; Sukhdeo, R.; Qu, R.; Jiang, J.; Reinke, A.W. Generation of a microsporidia species attribute database and analysis of the extensive ecological and phenotypic diversity of microsporidia. mBio 2021, 12, e0149021. [Google Scholar] [CrossRef] [PubMed]
- Duquesne, V.; Gastaldi, C.; Cont, A.D.; Cougoule, N.; Bober, A.; Brunain, M.; Chioveanu, G.; Demicoli, N.; Paulus, P.D.; Somalo, P.F.; et al. An international inter-laboratory study on Nosema spp. spore detection and quantification through microscopic examination of crushed honey bee abdomens. J. Microbiol. Methods 2021, 184, 106183. [Google Scholar] [CrossRef] [PubMed]
- Cilia, G.; Cabbri, R.; Maiorana, G.; Cardaio, I.; Dall´olio, R.; Nanetti, A. A novel TaqMan® assay for Nosema ceranae quantification in honey bee, based on the protein coding gene Hsp70. Eur. J. Protistol. 2018, 63, 44–50. [Google Scholar] [CrossRef]
- Kuzyšinová, K.; Molnár, L.; Toporčák, J.; Mudroňová, D.; Sabo, R.; Staroň, M.; Bielik, B. Atlas of Bee Diseases; The Univerzity of Veterinary Medicine and Pharmacy in Košice: Košice, Slovakia, 2016; pp. 44–48. [Google Scholar]
- Staroň, M.; Jurovčíková, J.; Čermáková, T.; Staroňová, D. A scientific note on incidence of Nosema apis and Nosema ceranae in Slovakia during the years 2009 and 2010. 2012. Available online: https://office.sjas-journal.org/index.php/sjas/article/view/269/257 (accessed on 15 March 2012).
- Abd-El-Samie, E.M.; Basuny, N.K.; Seyam, H. Molecular characterization of viruses found in honeybee (Apis mellifera) colonies infested with Varroa destructor and Nosema cerana in Egypt. Mol. Cell. Probes 2021, 57, 101731. [Google Scholar] [CrossRef]
- Panek, J.; Paris, L.; Roriz, D.; Mone, A.; Dubuffet, A.; Delbac, F.; Diogon, M.; Alaoui, H.E. Impact of the microsporidian Nosema ceranae on the gut epithelium renewal of the honeybee, Apis mellifera. J. Invertebr. Pathol. 2018, 159, 121–128. [Google Scholar] [CrossRef]
- Naree, S.; Benbow, M.E.; Suwannapong, G.; Ellis, J.D. Mitigating Nosema ceranae infection in western honey bee (Apis mellifera) workers using propolis collected from honey bee and stingless bee (Tetrigona apicalis) hives. J. Invertebr. Pathol. 2021, 185, 107666. [Google Scholar] [CrossRef]
- Paris, L.; Alaoui, H.E.; Delbac, F.; Diogon, M. Effects of the gut parasite Nosema ceranae on honey bee physiology and behavior. Curr. Opin. Insect Sci. 2018, 26, 149–154. [Google Scholar] [CrossRef]
- Mazur, E.D.; Gajda, A.M. Nosemosis in Honeybees: A Review Guide on Biology and Diagnostic Methods. Appl. Sci. 2022, 12, 5890. [Google Scholar] [CrossRef]
- Ptaszyńska, A.A.; Gancarz, M. Microsporidiosis Causing Necrotic Changes in the Honeybee Intestine. Appl. Sci. 2023, 13, 4957. [Google Scholar] [CrossRef]
- Sinpoo, C.; Paxton, R.J.; Disayathanoowat, T.; Krongdang, S.; Chantawannakul, P. Impact of Nosema ceranae and Nosema apis on individual worker bees of the two host species (Apis ceranae and Apis mellifera) and regulation of host immune response. J. Insect Physiol. 2018, 105, 1–8. [Google Scholar] [CrossRef]
- Kurze, C.; Conte, I.L.; Kryger, P.; Lewkovski, O.; Muller, T.; Moritz, R. Infection dynamics of Nosema ceranae in honey bee midgut and host cell apoptosis. J. Invertebr. Pathol. 2018, 154, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, N.; Eberl, H.J. Two routes of transmission for Nosema infections in a honey bee population model with polyethism 307 and time-periodic parameters can lead to drastically different qualitative model behavior. Commun. Nonlinear Sci. Numer. Simul. 2020, 84, 105207. [Google Scholar] [CrossRef]
- Valizadeh, P.; Guzman-Novoa, E.; Petukhova, T.; Goodwin, P.H. Effect of feeding chitosan or peptidoglycan on Nosema ceranae infection and gene expression related to stress and the innate immune response of honey bees (Apis mellifera). J. Invertebr. Pathol. 2021, 185, 107671. [Google Scholar] [CrossRef] [PubMed]
- Pachla, A.; Ptaszyńska, A.; Wicha, M.; Kunat, M.; Wydrych, J.; Olénska, E.; Malek, W. Insight into probiotic properties of lactic acid bacterial endosymbionts of Apis mellifera L. derived from the Polish apiary. Saudi J. Biol. Sci. 2021, 28, 1890–1899. [Google Scholar] [CrossRef]
- Titèra, D. Healthy and Sick Bees; Brázda: Praha, Czech Republic, 2017; pp. 32–41. [Google Scholar]
- Fries, I.; Chauzat, M.P.; Chen, Y.P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.; Martín-Hernández, R.; Natsopoulou, M.; et al. Standars methods for Nosema research. J. Apic. Res. 2013, 41, 375–392. [Google Scholar]
- Gisder, S.; Mockel, N.; Linde, A.; Genersch, E. A cell culture model for Nosema ceranae and Nosema apis allows new insights into the life cycle of these important honey bee-pathogenic microsporidia. Environ. Microbiol. 2011, 13, 404–413. [Google Scholar] [CrossRef]
- Maiolino, P.; Iafigliola, L.; Rinaldi, L.; Leva, G.D.; Restucci, B.; Martano, M. Histopathological findings of the midgut in European honey bee (Apis mellifera L.) naturally infected by Nosema spp. Vet. Med. Anim. Sci. 2014, 2, 4. [Google Scholar] [CrossRef]
- Babin, A.; Schurr, F.; Riviére, M.-P.; Chauzat, M.-P.; Dubois, E. Specific detection and quantification of three microsporidia infecting bees, Nosema apis, Nosema cerenae, and Nosema bombi, using probe- based real-time PCR. Eur. J. Protistol. 2022, 86, 125935. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Huang, Z.Y. Nosema ceranae, a newly identified pathogen of Apis mellifera in the USA and Asia. Apidologie 2010, 41, 364–374. [Google Scholar] [CrossRef]
- Chupia, V.; Patchanee, P.; Krutmuang, P.; Pikulkaew, S. Development and evaluation of loop-mediated isothermal amplification for rapid detection of Nosema ceranae in honeybee. Asian Pac. J. Trop. Dis. 2016, 6, 952–956. [Google Scholar] [CrossRef]
- Lannutti, L.; Mira, A.; Basualdo, M.; Rodriguez, G.; Erler, S.; Silva, V.; Gisder, S.; Genersch, E.; Florin-Christensen, M.; Schnittger, L. Development of a loop-mediated isothermal amplification (LAMP) and a direct LAMP for the specific detection of Nosema ceranae, a parasite of honey bees. Parasitol. Res. 2020, 119, 3947–3956. [Google Scholar] [CrossRef]
- Aronstein, K.A.; Webster, T.C.; Saldivar, E. A serological method for detection of Nosema ceranae. J. Appl. Microbiol. 2012, 114, 621–625. [Google Scholar] [CrossRef]
- Ostroverkhova, N.V.; Konusova, O.L.; Kucher, A.N.; Kireeva, T.N.; Rosseykina, S.A. Prevalence of the 318 Microsporidian Nosema spp. in Honey Bee Populations (Apis mellifera) in Some Ecological Regions of North Asia. Vet. Sci. 2020, 7, 111. [Google Scholar] [CrossRef]
- Valenčáková, A.; Luptáková, L.; Danišová, O.; Balent, P. Methods of Molecular Biology-Instructions for Practical Exercises for the Pharmacy Study Program; The University Veterinary Medicine and Pharmacy in Košice: Košice, Slovakia, 2015; pp. 14–15. [Google Scholar]
- Sulborska, A.; Horecka, B.; Cebrat, M.; Kowalczyk, M.; Skrzypek, T.H.; Kazimierczak, W.; Trytek, M.; Borsuk, G. Microsporidia Nosema spp.- obligate bee parasites are transmitted by air. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Hurná, B.; Valenčákova, A.; Súčik, M.; Staroň, M.; Maková, Z.; Galajda, R. The incidence of Nosema spp. in the honey bee colonies in Slovakia during 2021. In Proceedings of the ICOPA, Copenhagen, Denmark, 21–26 August 2022; p. 148 (#712). [Google Scholar]
- Pacini, A.; Mira, A.; Molineri, A.; Giacobino, A.; Cagnolo, N.B.; Aignasse, A.; Zago, L.; Izaguirre, M.; Merke, J.; Orellano, E.; et al. Distribution and prevalence of Nosema apis and N. ceranae in temperate and subtropical eco-regions of Argentina. J. Invertebr. Pathol. 2016, 141, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Matović, K.; Vidanović, D.; Manić, M.; Stojiljković, S.; Radojičić, S.; Debeljak, Z.; Šekler, M.; Ćirić, J. Twenty-five-year study of Nosema spp. in honey bees (Apis mellifera) in Serbia. Saudi J. Biol. Sci. 2020, 27, 518–523. [Google Scholar] [CrossRef]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Intebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef]
- Salkova, D.; Shumkova, R.; Palova, N.; Neov, B.; Radoslavov, G.; Hristov, P. Molecular Detection of Nosema spp. in Honey in Bulgaria. Vet. Sci. 2021, 9, 10. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Bee Queen Breeder | ||
|---|---|---|
| YEAR | 2021 | 2022 |
| west | 23 | 19 |
| middle | 6 | 5 |
| east | 10 | 8 |
| TOTAL: | 39 | 32 |
| Microscopic Diagnostics in 2021 | |||
|---|---|---|---|
| Region | number of samples | Positive samples | Negative samples |
| west | 859 | 182 | 677 |
| middle | 569 | 142 | 427 |
| east | 600 | 146 | 454 |
| TOTAL: | 2028 | 470 | 1558 |
| Molecular Diagnostics in 2021 | |||
| Region | number of samples | Positive samples | Negative samples |
| west | 40 | 34 | 6 |
| middle | 19 | 18 | 1 |
| east | 22 | 12 | 1 |
| TOTAL: | 81 | 73 | 8 |
| Microscopic Diagnostics in 2022 | |||
| Region | number of samples | Positive samples | Negative samples |
| west | 915 | 211 | 704 |
| middle | 568 | 129 | 439 |
| east | 507 | 112 | 395 |
| TOTAL: | 1990 | 452 | 1538 |
| Molecular Diagnostics in 2022 | |||
| Region | number of samples | Positive samples | Negative samples |
| west | 195 | 193 | 2 |
| middle | 123 | 118 | 5 |
| east | 108 | 104 | 4 |
| TOTAL: | 426 | 415 | 11 |
| Microscopic Diagnostics | ||||
|---|---|---|---|---|
| Region | Samples | |||
| 2021 | 2022 | |||
| Positive | Negative | Positive | Negative | |
| west | 21.19% | 78.81% | 23.06% | 76.94% |
| middle | 24.96% | 75.04% | 22.71% | 77.29% |
| east | 24.33% | 75.67% | 22.09% | 77.91% |
| Mean | 23.49% | 76.51% | 22.62% | 77.38% |
| Median | 24.33% | 75.67% | 22.71% | 77.29% |
| Min | 21.19% | 75.04% | 22.09% | 76.94% |
| Max | 24.96% | 78.81% | 23.06% | 77.91% |
| R2 | 0.6062 | 0.6062 | 0.9745 | 0.9745 |
| R | 0.7785 | 0.7785 | 0.9871 | 0.9871 |
| Molecular Diagnostics | ||||
| Region | Samples | |||
| 2021 | 2022 | |||
| Positive | Negative | Positive | Negative | |
| west | 85.00% | 15.00% | 98.97% | 1.03% |
| middle | 94.74% | 5.26% | 95.93% | 4.07% |
| east | 95.45% | 4.55% | 96.30% | 3.70% |
| Mean | 91.73% | 8.27% | 97.07% | 2.93% |
| Median | 94.74% | 5.26% | 96.30% | 3.70% |
| Minimum Value | 85.00% | 15.00% | 95.93% | 1.03% |
| Maximum Value | 95.45% | 94.74% | 98.97% | 4.07% |
| R2 | 0.8012 | 0.6504 | 0.6504 | 0.8012 |
| R | 0.8950 | 0.8064 | 0.8064 | 0.8950 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurná, B.; Sučik, M.; Staroň, M.; Tutka, Š.; Maková, Z.; Galajda, R.; Valenčáková, A. Molecular Detection of Nosema spp. in Three Eco Regions of Slovakia. Curr. Issues Mol. Biol. 2023, 45, 4814-4825. https://doi.org/10.3390/cimb45060306
Hurná B, Sučik M, Staroň M, Tutka Š, Maková Z, Galajda R, Valenčáková A. Molecular Detection of Nosema spp. in Three Eco Regions of Slovakia. Current Issues in Molecular Biology. 2023; 45(6):4814-4825. https://doi.org/10.3390/cimb45060306
Chicago/Turabian StyleHurná, Beáta, Monika Sučik, Martin Staroň, Štefan Tutka, Zuzana Maková, Richard Galajda, and Alexandra Valenčáková. 2023. "Molecular Detection of Nosema spp. in Three Eco Regions of Slovakia" Current Issues in Molecular Biology 45, no. 6: 4814-4825. https://doi.org/10.3390/cimb45060306