
Genetic Dissection of Salt Tolerance and Yield Traits of Geng (japonica) Rice by Selective Subspecific Introgression

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Selection Efficiencies for Improving ST
2.2. Phenological Mechanisms Underlying Rice ST
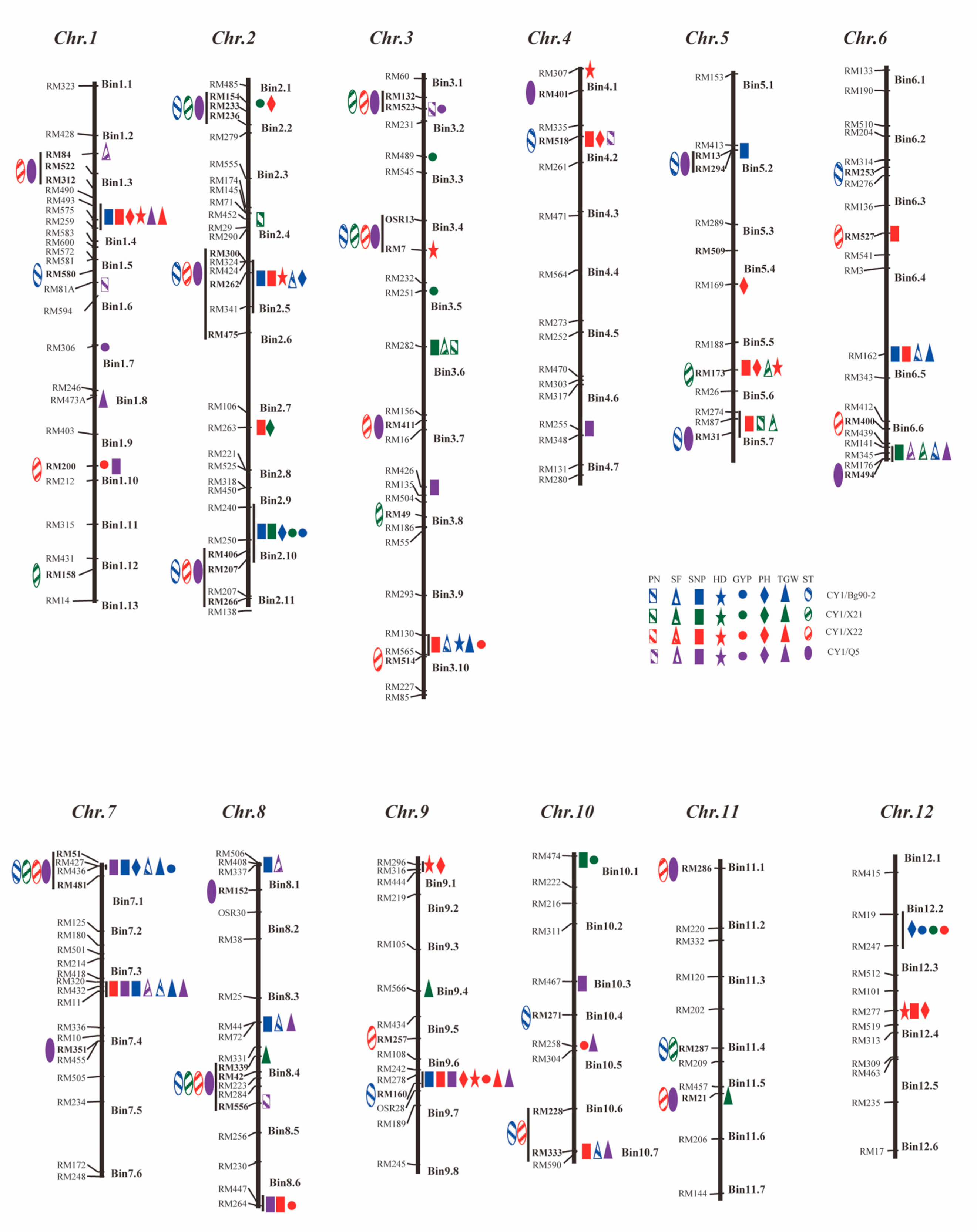
2.3. Identification and Mapping of ST QTLs
2.4. Correlated Phenotypic Responses from Selection for ST and Mapping of QTLs for ‘Non-Target’ Traits
2.5. Identification and Mapping of QTLs Affecting the Yield Traits under Normal Conditions
3. Discussion
3.1. The Phenological Mechanisms of Rice ST at the Vegetative and Reproductive Stages
3.2. Genetic Basis of Rice ST and Its Associated Yield Traits
3.3. Implications for Improving Rice ST at The Vegetative and Reproductive Stages
4. Materials and Methods
4.1. Plant Materials
4.2. Screening BC2F4 Progeny for Salt Tolerance at the Vegetative and Reproductive Stages
4.3. The Genotyping Experiment
4.4. Statistical Analyses
4.5. Identifying QTLs Affecting ST and Yield Traits under Non-Stress Normal Conditions
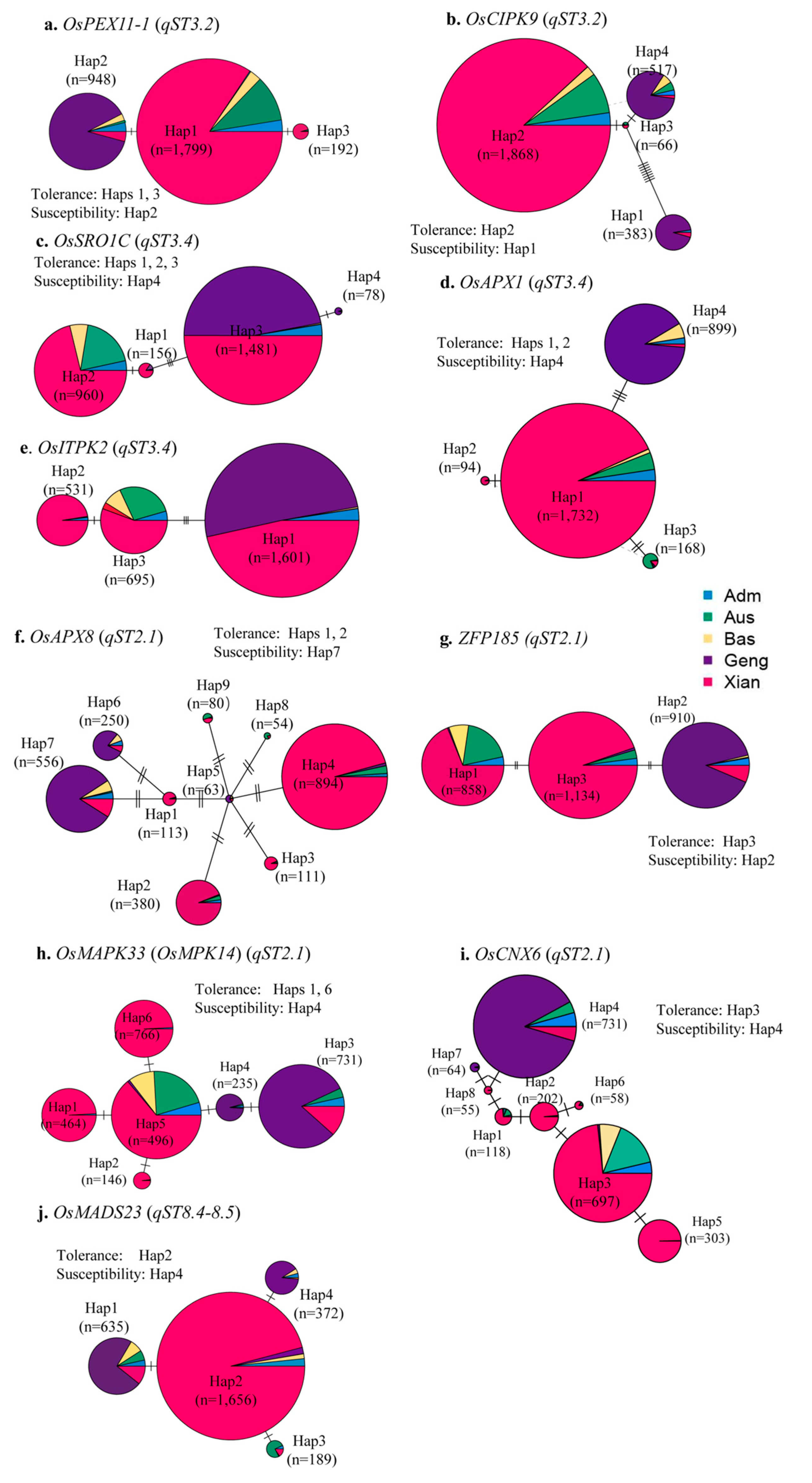
4.6. Candidate Genes for the ST QTLs and Their Gene CDS Haplotype (gcHap) Diversity in Rice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ganie, S.A.; Molla, K.A.; Henry, R.J.; Bhat, K.; Mondal, T.K. Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Li, Y.; Huang, R. Advances and challenges in the breeding of salt-tolerant rice. Int. J. Mol. Sci. 2020, 21, 8385. [Google Scholar] [CrossRef] [PubMed]
- Peltier, W.R.; Tushingham, A. Global sea level rise and the greenhouse effect: Might they be connected? Science 1989, 244, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Thomson, M.; Vergara, G.; Rahman, M.; Singh, R.; Gregorio, G.; Mackill, D. Designing resilient rice varieties for coastal deltas using modern breeding tools. In Tropical Deltas and Coastal Zones: Food Production, Communities and Environment at the Land and Water Interface; CABI: Wallingford, UK, 2010; pp. 154–165. [Google Scholar]
- Quan, R.; Wang, J.; Yang, D.; Zhang, H.; Zhang, Z.; Huang, R. EIN3 and SOS2 synergistically modulate plant salt tolerance. Sci. Rep. 2017, 7, 44637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Xu, P.; Huang, J.; Ma, S.; Xie, X.; Tao, D.; Chen, L.; Liu, Y.-G. Interspecific hybrid sterility in rice is mediated by OgTPR1 at the S1 locus encoding a peptidase-like protein. Mol. Plant 2017, 10, 1137–1140. [Google Scholar] [CrossRef]
- Pearson, G.A.; Ayers, A.; Eberhard, D. Relative salt tolerance of rice during germination and early seedling development. Soil Sci. 1966, 102, 151–156. [Google Scholar] [CrossRef]
- Noor, A.U.Z.; Nurnabi Azad Jewel, G.; Haque, T.; Elias, S.M.; Biswas, S.; Rahman, M.S.; Seraj, Z.I. Validation of QTLs in Bangladeshi rice landrace Horkuch responsible for salt tolerance in seedling stage and maturation. Acta Physiol. Plant. 2019, 41, 173. [Google Scholar] [CrossRef]
- Wing, R.A.; Purugganan, M.D.; Zhang, Q. The rice genome revolution: From an ancient grain to Green Super Rice. Nat. Rev. Genet. 2018, 19, 505–517. [Google Scholar] [CrossRef]
- Li, Z.-K.; Xu, J.-L. Breeding for drought and salt tolerant rice (Oryza sativa L.): Progress and perspectives. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Springer: Dordrecht, The Netherlands, 2007; pp. 531–564. [Google Scholar]
- Mondal, S.; Septiningsih, E.M.; Singh, R.K.; Thomson, M.J. Mapping QTLs for Reproductive Stage Salinity Tolerance in Rice Using a Cross between Hasawi and BRRI dhan28. Int. J. Mol. Sci. 2022, 23, 11376. [Google Scholar] [CrossRef]
- Batayeva, D.; Labaco, B.; Ye, C.; Li, X.; Usenbekov, B.; Rysbekova, A.; Dyuskalieva, G.; Vergara, G.; Reinke, R.; Leung, H. Genome-wide association study of seedling stage salinity tolerance in temperate japonica rice germplasm. BMC Genet. 2018, 19, 2. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Choudhary, M.; Halder, T.; Prakash, N.R.; Singh, V.; Vineeth, V.T.; Sheoran, S.; Longmei, N.; Rakshit, S.; Siddique, K.H. Salinity stress tolerance and omics approaches: Revisiting the progress and achievements in major cereal crops. Heredity 2022, 128, 497–518. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Mishra, B.; Chauhan, M.; Yeo, A.; Flowers, S.; Flowers, T. Solution culture for screening rice varieties for sodicity tolerance. J. Agric. Sci. 2002, 139, 327–333. [Google Scholar] [CrossRef]
- Jena, K.; Mackill, D. Molecular markers and their use in marker-assisted selection in rice. Crop Sci. 2008, 48, 1266–1276. [Google Scholar] [CrossRef]
- Calapit-Palao, C.D.O. Identification of QTL for salinity tolerance at reproductive stage in rice (Oryza sativa L.). Master’s Thesis, University of the Philippines Los Baños, Los Baños, Philippines, 2010; p. 116. [Google Scholar]
- Singh, R.K.; Kota, S.; Flowers, T.J. Salt tolerance in rice: Seedling and reproductive stage QTL mapping come of age. Theor. Appl. Genet. 2021, 134, 3495–3533. [Google Scholar] [CrossRef]
- Mohammadi, R.; Mendioro, M.S.; Diaz, G.Q.; Gregorio, G.B.; Singh, R.K. Mapping quantitative trait loci associated with yield and yield components under reproductive stage salinity stress in rice (Oryza sativa L.). J. Genet. 2013, 92, 433–443. [Google Scholar] [CrossRef]
- Chen, C.; Norton, G.J.; Price, A.H. Genome-wide association mapping for salt tolerance of rice seedlings grown in hydroponic and soil systems using the Bengal and Assam Aus panel. Front. Plant Sci. 2020, 11, 576479. [Google Scholar] [CrossRef]
- Takehisa, H.; Shimodate, T.; Fukuta, Y.; Ueda, T.; Yano, M.; Yamaya, T.; Kameya, T.; Sato, T. Identification of quantitative trait loci for plant growth of rice in paddy field flooded with salt water. Field Crops Res. 2004, 89, 85–95. [Google Scholar] [CrossRef]
- Ammar, M.; Pandit, A.; Singh, R.; Sameena, S.; Chauhan, M.; Singh, A.; Sharma, P.; Gaikwad, K.; Sharma, T.; Mohapatra, T. Mapping of QTLs controlling Na+, K+ and CI− ion concentrations in salt tolerant indica rice variety CSR27. J. Plant Biochem. Biotechnol. 2009, 18, 139–150. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, Y.; Gui, R.; Wang, Z.; Li, Z.; Han, Y.; Guo, X.; Sun, J. Comparative multi-omics analysis of hypoxic germination tolerance in weedy rice embryos and coleoptiles. Genomics 2021, 113, 3337–3348. [Google Scholar] [CrossRef]
- Murray, M.; Thompson, W. Protocol of DNA isolation. Nucl. Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zhou, X.-G.; Yan, Z.; Tabien, R.E.; Wilson, L.T.; Wang, L. Hybrid rice outperforms inbred rice in resistance to sheath blight and narrow brown leaf spot. Plant Dis. 2021, 105, 2981–2989. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M. Genomic variation in 3,010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartl, D.L.; Clark, A.G.; Clark, A.G. Principles of Population Genetics; Sinauer associates Sunderland: Sunderland, MA, USA, 1997; Volume 116. [Google Scholar]
- Falconer, D.; Mackay, T. Introduction to Quantitative Genetics; Longman Group: Harlow, UK, 1996; Volume 12. [Google Scholar]
- Li, Z.-K.; Fu, B.-Y.; Gao, Y.-M.; Xu, J.-L.; Ali, J.; Lafitte, H.; Jiang, Y.-Z.; Rey, J.D.; Vijayakumar, C.; Maghirang, R. Genome-wide introgression lines and their use in genetic and molecular dissection of complex phenotypes in rice (Oryza sativa L.). Plant Mol. Biol. 2005, 59, 33–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhai, H.-Q.; Paterson, A.H.; Xu, J.-L.; Gao, Y.-M.; Zheng, T.-Q.; Wu, R.-L.; Fu, B.-Y.; Ali, J.; Li, Z.-K. Dissecting genetic networks underlying complex phenotypes: The theoretical framework. PLoS ONE 2011, 6, e14541. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, H.; Sun, C.; Ma, Q.; Bu, H.; Chong, K.; Xu, Y. A C2H2 zinc-finger protein OsZFP213 interacts with OsMAPK3 to enhance salt tolerance in rice. J. Plant Physiol. 2018, 229, 100–110. [Google Scholar] [CrossRef]
- Shabala, S.; Alnayef, M.; Bose, J.; Chen, Z.-H.; Venkataraman, G.; Zhou, M.; Shabala, L.; Yu, M. Revealing the role of the Calcineurin B-like protein-interacting protein kinase 9 (CIPK9) in Rice adaptive responses to salinity, osmotic stress, and K+ deficiency. Plants 2021, 10, 1513. [Google Scholar] [CrossRef]
- You, J.; Zong, W.; Li, X.; Ning, J.; Hu, H.; Li, X.; Xiao, J.; Xiong, L. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Qingjie, G.; Qiuxiang, L. Expression of rice OsAPX1 gene and its salt tolerance in tobacco. Mol. Plant Breed. 2007, 5, 1–7. [Google Scholar]
- Fang, T.; Dong, Y.; Li, Y.; Chen, D.; Chen, X. The role of rice chloroplast ascorbate peroxidase in drought and high salt stress. Plant Physiol. 2015, 51, 2207–2213. [Google Scholar] [CrossRef]
- Lee, S.-K.; Kim, B.-G.; Kwon, T.-R.; Jeong, M.-J.; Park, S.-R.; Lee, J.-W.; Byun, M.-O.; Kwon, H.-B.; Matthews, B.F.; Hong, C.-B. Overexpression of the mitogen-activated protein kinase gene OsMAPK33 enhances sensitivity to salt stress in rice (Oryza sativa L.). J. Biosci. 2011, 36, 139–151. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Yu, Y.; Kong, L.; Liu, Y.; Liu, Z.; Li, H.; Wei, P.; Liu, M.; Zhou, H. Identification and characterization of the rice pre-harvest sprouting mutants involved in molybdenum cofactor biosynthesis. New Phytol. 2019, 222, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yu, B.; Wu, Q.; Min, Q.; Zeng, R.; Xie, Z.; Huang, J. OsMADS23 phosphorylated by SAPK9 confers drought and salt tolerance by regulating ABA biosynthesis in rice. PLoS Genet. 2021, 17, e1009699. [Google Scholar] [CrossRef] [PubMed]
- Akbar, M.; Ponnamperuma, F. Saline soils of South and Southeast Asia as potential rice lands. In Rice Research Strategies for the Future; IRRI: Makati, Philippines, 1982; pp. 265–281. [Google Scholar]
- Lafitte, H.; Li, Z.; Vijayakumar, C.; Gao, Y.; Shi, Y.; Xu, J.; Fu, B.; Yu, S.; Ali, A.; Domingo, J. Improvement of rice drought tolerance through backcross breeding: Evaluation of donors and selection in drought nurseries. Field Crops Res. 2006, 97, 77–86. [Google Scholar] [CrossRef]
- Ali, A.; Xu, J.; Ismail, A.; Fu, B.; Vijaykumar, C.; Gao, Y.; Domingo, J.; Maghirang, R.; Yu, S.; Gregorio, G. Hidden diversity for abiotic and biotic stress tolerances in the primary gene pool of rice revealed by a large backcross breeding program. Field Crops Res. 2006, 97, 66–76. [Google Scholar] [CrossRef]
- He, Y.; Zheng, T.; Hao, X.; Wang, L.; Gao, Y.; Hua, Z.; Zhai, H.; Xu, J.; Xu, Z.; Zhu, L. Yield performances of japonica introgression lines selected for drought tolerance in a BC breeding programme. Plant Breed. 2010, 129, 167–175. [Google Scholar] [CrossRef]
- Meng, L.; Lin, X.; Wang, J.; Chen, K.; Cui, Y.; Xu, J.; Li, Z. Simultaneous improvement in cold tolerance and yield of temperate japonica rice (Oryza sativa L.) by introgression breeding. Plant Breed. 2013, 132, 604–612. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, Q.; Wang, S.; Hong, Y.; Wang, Z. Rice and cold stress: Methods for its evaluation and summary of cold tolerance-related quantitative trait loci. Rice 2014, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Meng, L.; Lin, X.; Cui, Y.; Pang, Y.; Xu, J.; Li, Z. QTL and QTL networks for cold tolerance at the reproductive stage detected using selective introgression in rice. PLoS ONE 2018, 13, e0200846. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Zhang, W.; Lin, X.; Xu, S.; Xu, J.; Li, Z. Simultaneous improvement and genetic dissection of drought tolerance using selected breeding populations of rice. Front. Plant Sci. 2018, 9, 320. [Google Scholar] [CrossRef] [Green Version]
- Jagadish, S.K.; Craufurd, P.Q.; Wheeler, T. High temperature stress and spikelet fertility in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 1627–1635. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chen, K.; Zhao, X.; Wang, X.; Shen, C.; Zhu, Y.; Dai, M.; Qiu, X.; Yang, R.; Xing, D.; et al. Identification of genes for salt tolerance and yield-related traits in rice plants grown hydroponically and under saline field conditions by genome-wide association study. Rice 2019, 12, 24. [Google Scholar] [CrossRef] [PubMed]
- Khatun, S.; Flowers, T. Effects of salinity on seed set in rice. Plant Cell Environ. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Fan, X.; Jiang, H.; Meng, L.; Chen, J. Gene mapping, cloning and association analysis for salt tolerance in rice. Inter Natl. J. Mol. Sci. 2021, 22, 11674. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Abdullah, Z. Salinity–sodicity induced changes in reproductive physiology of rice (Oryza sativa L.) under dense soil conditions. Environ. Exp. Bot. 2003, 49, 145–157. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Gautam, R.; Singh, R.; Krishnamurthy, S.L.; Ali, S.; Sakthivel, K.; Iquebal, M.A.; Rai, A. Harmonizing technological advances in phenomics and genomics for enhanced salt tolerance in rice from a practical perspective. Rice 2019, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Weber, D. Adaptive mechanisms of halophytes in desert regions. In Salinity and Water Stress; Springer: Dordrecht, The Netherlands, 2009; pp. 179–185. [Google Scholar]
- Li, Z.-K.; Zhang, F. Rice breeding in the post-genomics era: From concept to practice. Curr. Opin. Plant Biol. 2013, 16, 261–269. [Google Scholar] [CrossRef]
- Kim, J.H.; Jang, C.S. E3 ligase, the Oryza sativa salt-induced RING finger protein 4 (OsSIRP4), negatively regulates salt stress responses via degradation of the OsPEX11-1 protein. Plant Mol. Biol. 2021, 105, 231–245. [Google Scholar] [CrossRef]
- Ren, Z.H.; Gao, J.P.; Li, L.G. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Lines | Donor | GYP (g) 1 | SF(%) | FGN | SNP | TGW (g) | PN | PH (cm) | HD (Days) |
|---|---|---|---|---|---|---|---|---|---|
| A-11 | Bg90-2 | 29.3 | 81.7 | 150.5 | 184.3 | 22.6 | 7.7 | 126.1 | 108.0 |
| A-12 | Bg90-2 | 40.1 | 92.9 | 163.5 | 176.1 | 22.5 | 9.7 | 117.7 | 108.0 |
| A-21 | Bg90-2 | 25.9 | 92.5 | 153.4 | 165.9 | 24.3 | 6.7 | 124.5 | 95.0 |
| A-3 | Bg90-2 | 27.3 | 95.3 | 153.6 | 161.2 | 26.2 | 7.0 | 127.3 | 112.0 |
| B-5 | X21 | 29.5 | 92.0 | 120.3 | 130.8 | 23.8 | 9.7 | 115.0 | 108.0 |
| C-2 | X22 | 28.0 | 87.0 | 132.5 | 152.3 | 26.7 | 8.3 | 127.1 | 104.0 |
| C-30 | X22 | 34.1 | 83.1 | 161.0 | 193.7 | 26.6 | 7.7 | 105.7 | 103.0 |
| D-10 | Q5 | 32.9 | 92.5 | 149.6 | 161.7 | 25.4 | 8.7 | 130.8 | 108.0 |
| D-9 | Q5 | 20.0 | 93.0 | 106.8 | 114.8 | 22.5 | 7.7 | 110.6 | 102.0 |
| Mean | 29.7 | 90.0 | 143.5 | 160.1 | 24.5 | 8.1 | 120.5 | 105.3 | |
| CY1 | 22.6 | 87.5 | 124.0 | 134.2 | 24.2 | 7.2 | 124.3 | 105.3 | |
| A-11 | Bg90-2 | 12.8 | 63.0 | 473.0 | 187.8 | 23.5 | 4.0 | 96.0 | 107.0 |
| A-12 | Bg90-2 | 10.9 | 68.6 | 386.0 | 140.8 | 26.5 | 4.0 | 89.0 | 108.0 |
| A-21 | Bg90-2 | 14.6 | 73.5 | 476.0 | 162.0 | 27.5 | 4.0 | 93.0 | 107.0 |
| A-3 | Bg90-2 | 8.4 | 83.9 | 224.0 | 133.5 | 27.0 | 2.0 | 85.0 | 108.0 |
| B-5 | X21 | 9.4 | 66.0 | 285.0 | 72.0 | 23.3 | 6.0 | 76.0 | 118.0 |
| C-2 | X22 | 11.5 | 78.4 | 373.0 | 95.2 | 22.0 | 5.0 | 84.0 | 115.0 |
| C-30 | X22 | 13.9 | 73.2 | 583.0 | 132.7 | 23.3 | 6.0 | 118.0 | 106.0 |
| D-10 | Q5 | 11.1 | 81.1 | 344.0 | 84.8 | 23.3 | 5.0 | 98.0 | 110.0 |
| D-9 | Q5 | 11.7 | 63.1 | 352.0 | 93.0 | 26.3 | 6.0 | 91.0 | 112.0 |
| Mean | 11.6 | 72.3 | 388.4 | 122.4 | 24.7 | 4.7 | 92.2 | 110.1 | |
| CY1 | 3.5 | 54 | 48.8 | 99.4 | 20.3 | 3.6 | 85.5 | 117.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Feng, T.; Zhang, C.; Zhang, F.; Li, H.; Chen, Y.; Liang, L.; Zhang, C.; Zeng, W.; Liu, E.; et al. Genetic Dissection of Salt Tolerance and Yield Traits of Geng (japonica) Rice by Selective Subspecific Introgression. Curr. Issues Mol. Biol. 2023, 45, 4796-4813. https://doi.org/10.3390/cimb45060305
Li S, Feng T, Zhang C, Zhang F, Li H, Chen Y, Liang L, Zhang C, Zeng W, Liu E, et al. Genetic Dissection of Salt Tolerance and Yield Traits of Geng (japonica) Rice by Selective Subspecific Introgression. Current Issues in Molecular Biology. 2023; 45(6):4796-4813. https://doi.org/10.3390/cimb45060305
Chicago/Turabian StyleLi, Simin, Ting Feng, Chenyang Zhang, Fanlin Zhang, Hua Li, Yanjun Chen, Lunping Liang, Chaopu Zhang, Wei Zeng, Erbao Liu, and et al. 2023. "Genetic Dissection of Salt Tolerance and Yield Traits of Geng (japonica) Rice by Selective Subspecific Introgression" Current Issues in Molecular Biology 45, no. 6: 4796-4813. https://doi.org/10.3390/cimb45060305