
Effects of Whole-Body Vibration and Manually Assisted Locomotor Therapy on Neurotrophin-3 Expression and Microglia/Macrophage Mobilization Following Thoracic Spinal Cord Injury in Rats

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Groups
2.2. Pre-Injury Conditioning
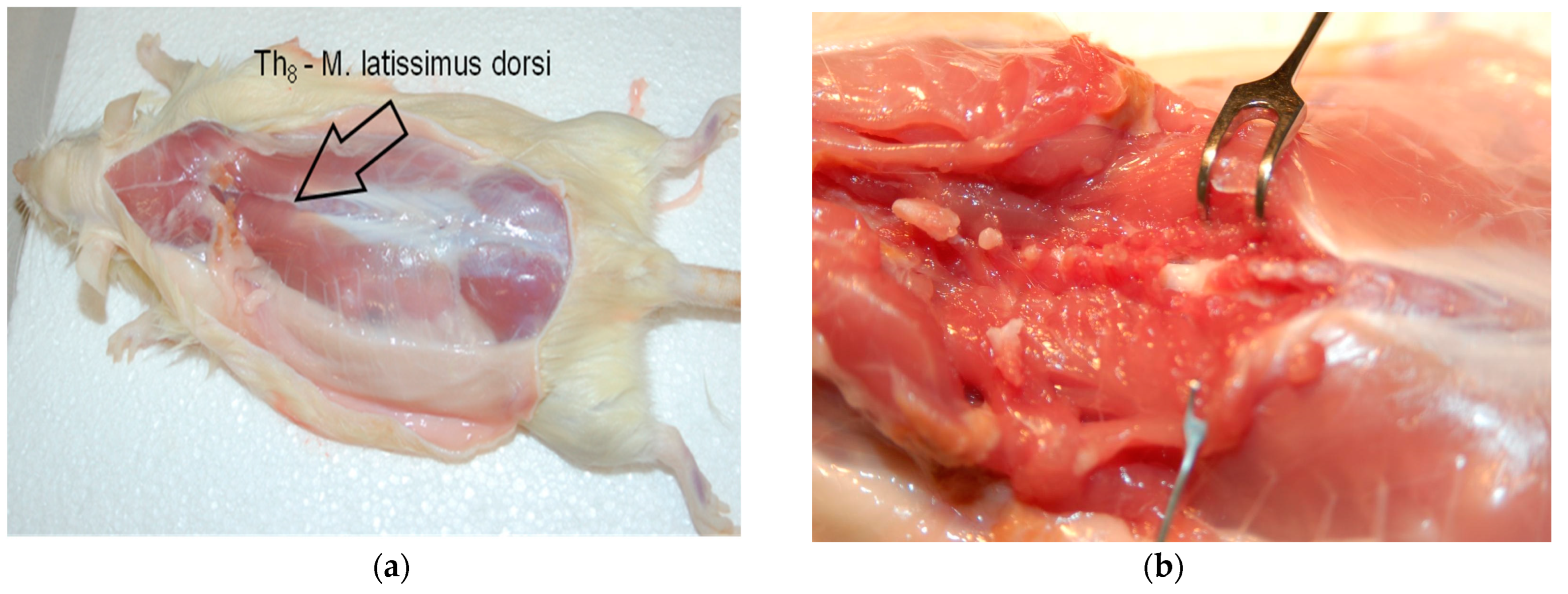
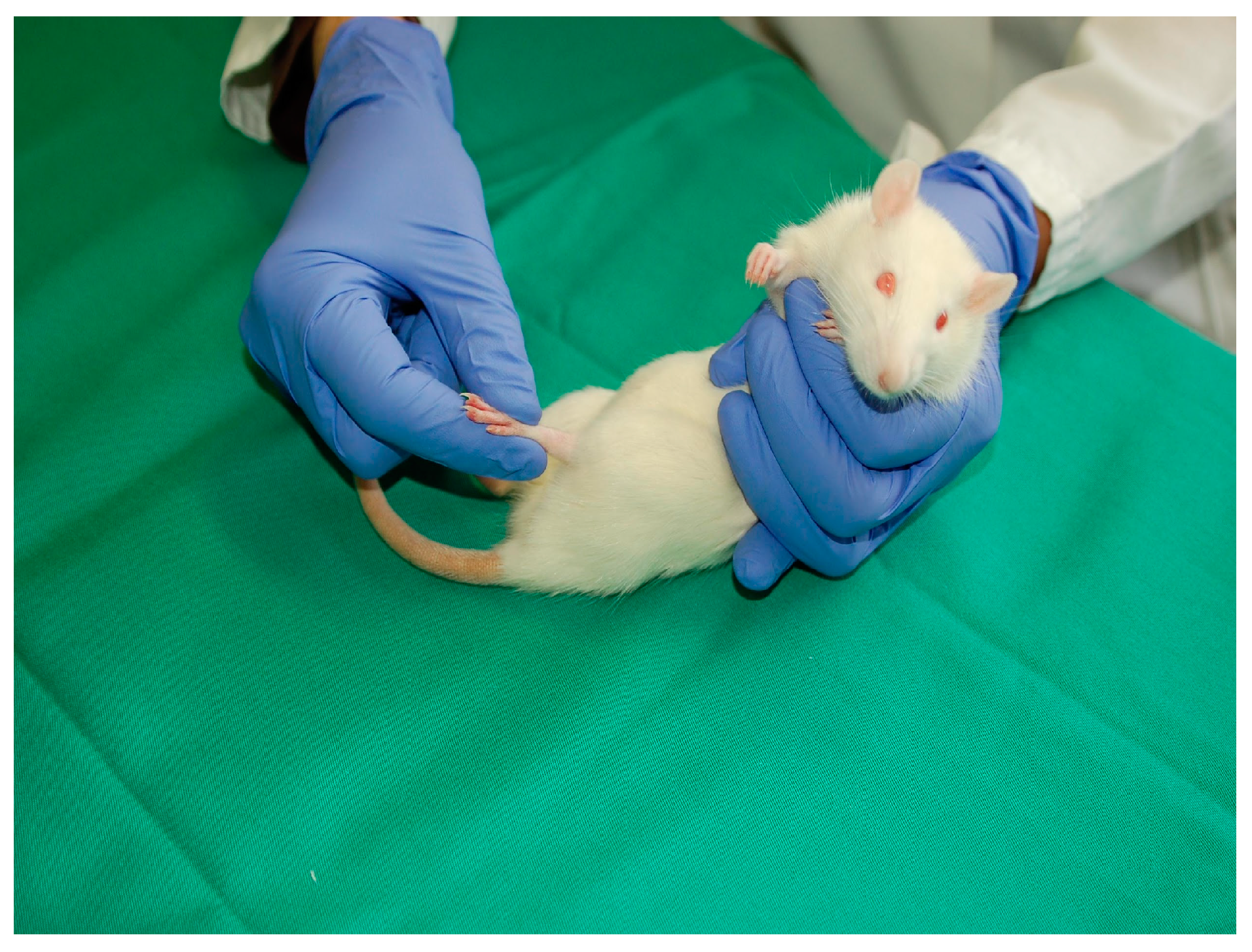
2.3. Spinal Cord Injury
2.4. Whole-Body Vibration (WBV) Therapy
2.5. Passive Flexion–Extension (PFE)
2.6. Locomotor Performance
2.7. Tissue Preparation
2.8. Immunohistochemistry
2.9. Statistical Analyses
3. Results
3.1. Effects of WBV and PFE Therapy on Functional Recovery after SCI
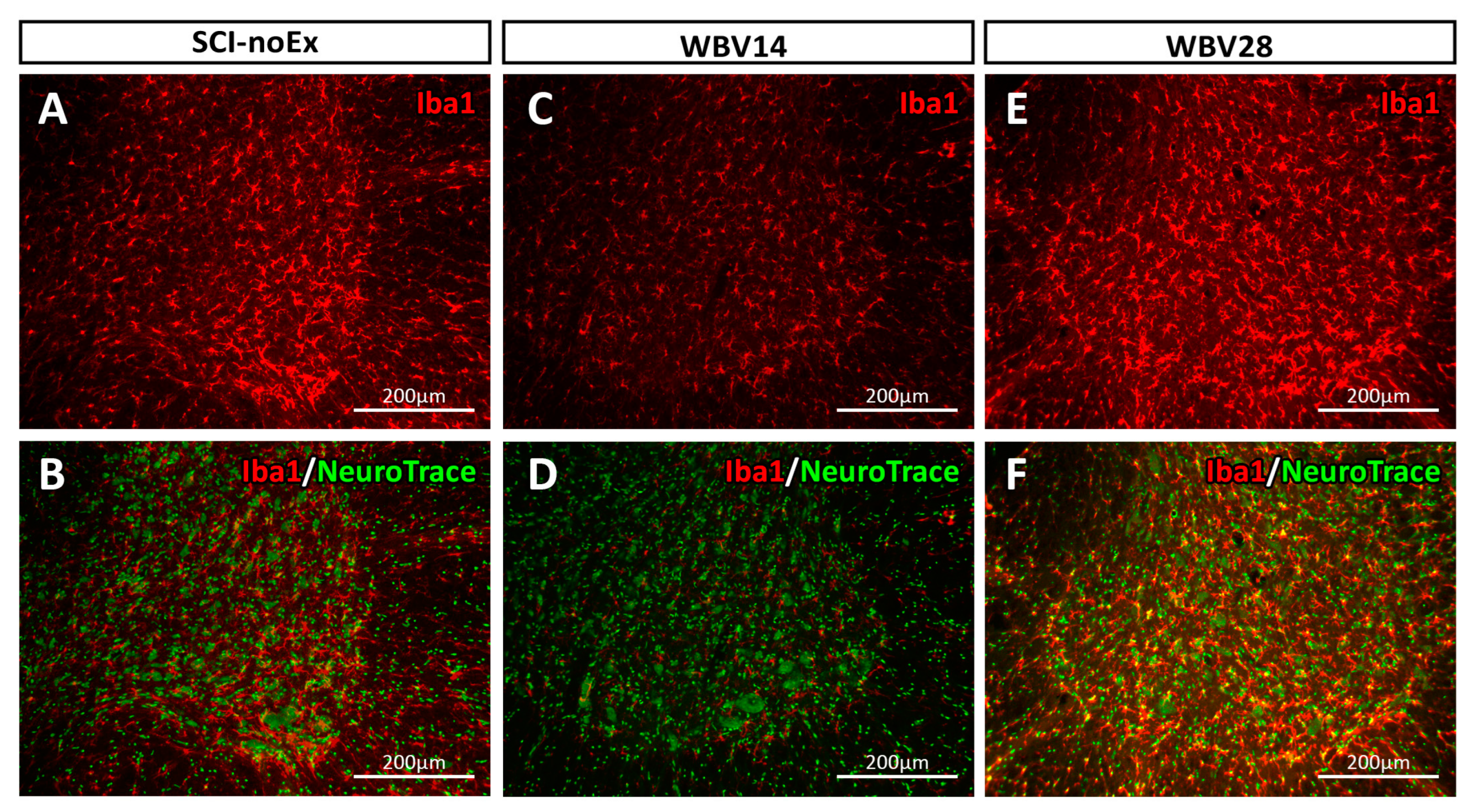
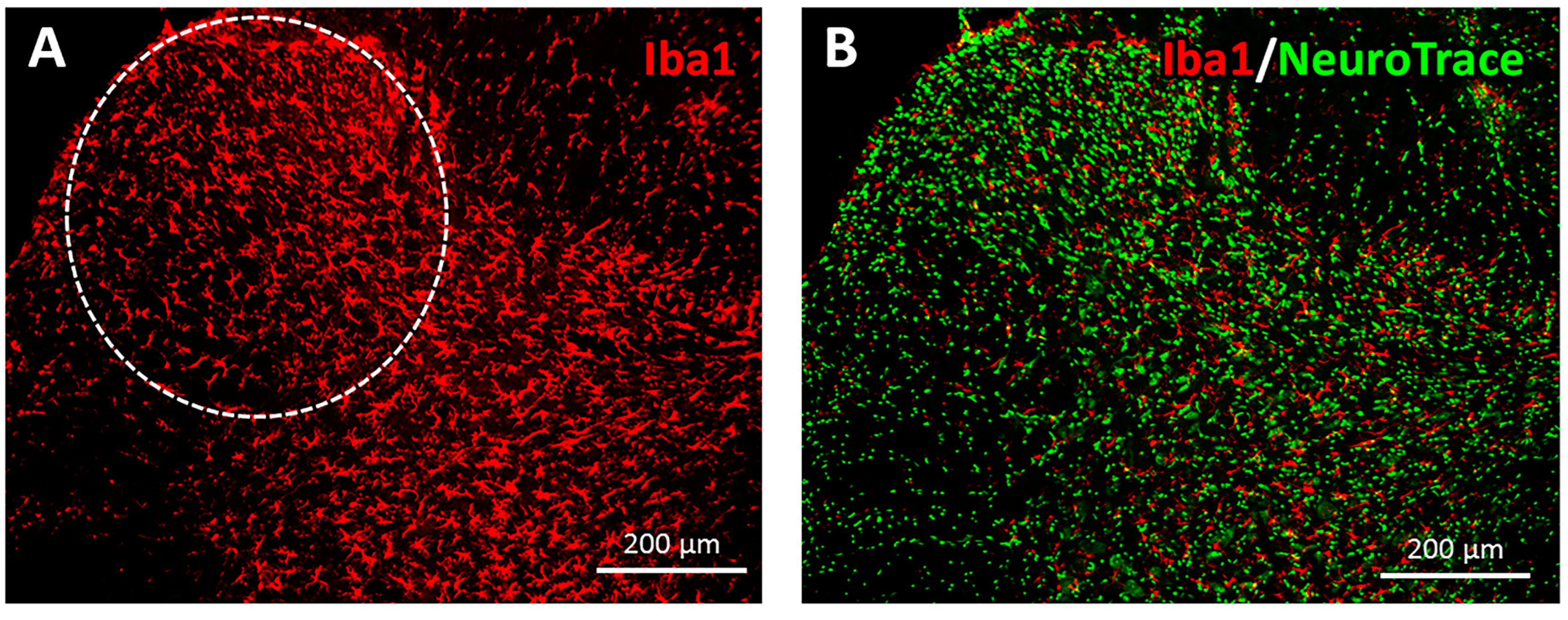
3.2. Immunohistochemical Analysis of Iba1 Expression
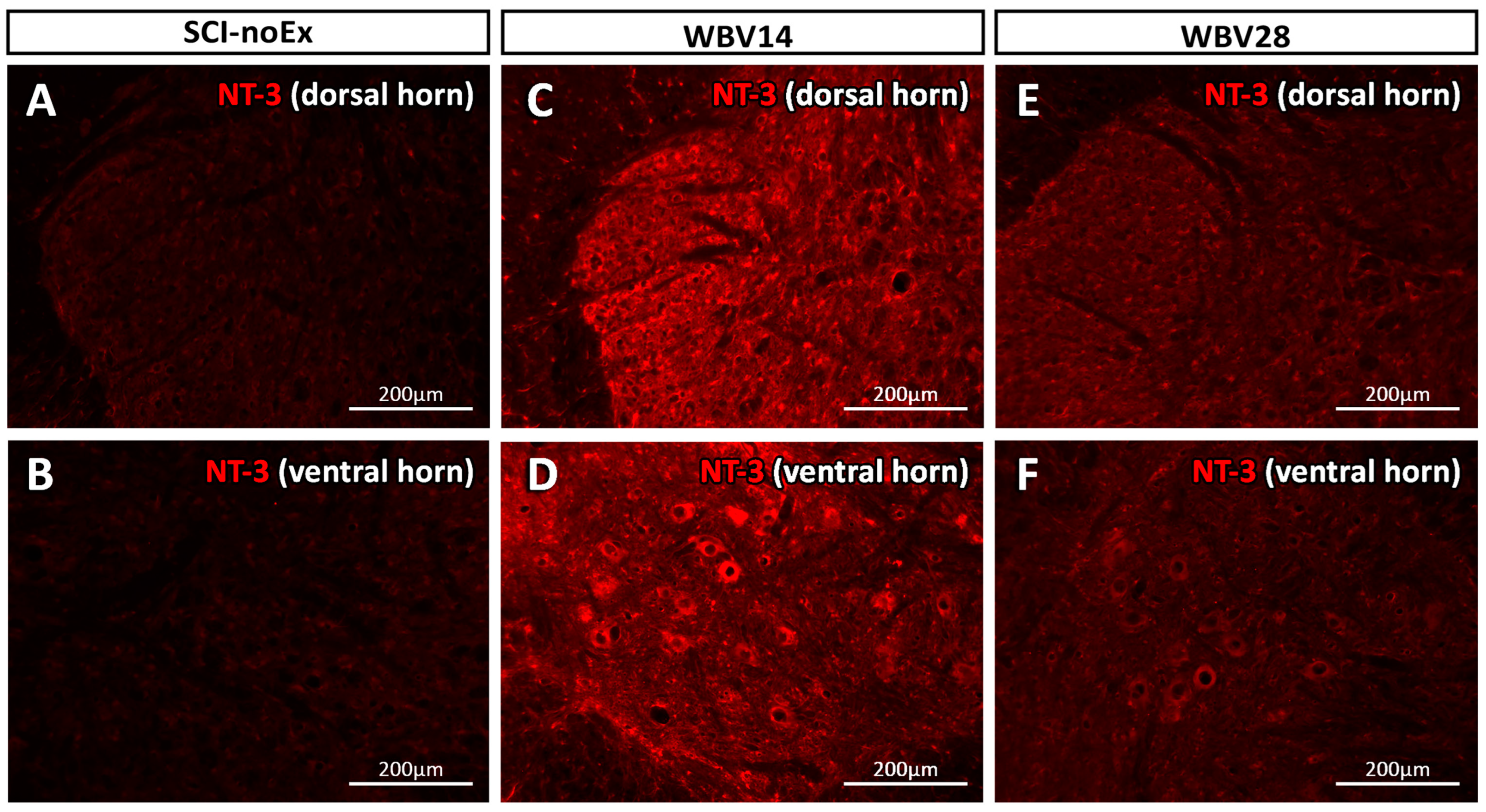
3.3. Immunohistochemical Analysis of NT-3 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bills, K.B.; Clarke, T.; Major, G.H.; Jacobson, C.B.; Blotter, J.D.; Feland, J.B.; Steffensen, S.C. Targeted Subcutaneous Vibration With Single-Neuron Electrophysiology As a Novel Method for Understanding the Central Effects of Peripheral Vibrational Therapy in a Rodent Model. Dose Response 2019, 17. [Google Scholar] [CrossRef] [PubMed]
- Herren, K.; Schmid, S.; Rogan, S.; Radlinger, L. Effects of Stochastic Resonance Whole-Body Vibration in Individuals with Unilateral Brain Lesion: A Single-Blind Randomized Controlled Trial: Whole-Body Vibration and Neuromuscular Function. Rehabil. Res. Pract. 2018, 2018, 9319258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Li, N.; Zhu, L.; Lin, Y.; Cheng, H. The Microglial Activation Profile and Associated Factors after Experimental Spinal Cord Injury in Rats. Neuropsychiatr. Dis. Treat. 2018, 14, 2401–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroner, A.; Rosas Almanza, J. Role of Microglia in Spinal Cord Injury. Neurosci. Lett. 2019, 709, 134370. [Google Scholar] [CrossRef]
- Bermudez, S.; Khayrullina, G.; Zhao, Y.; Byrnes, K.R. NADPH Oxidase Isoform Expression Is Temporally Regulated and May Contribute to Microglial/Macrophage Polarization after Spinal Cord Injury. Mol. Cell. Neurosci. 2016, 77, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Wang, J.; Ding, Y.; Wang, L.; Zhu, Y.-J. Current Knowledge of Microglia in Traumatic Spinal Cord Injury. Front. Neurol. 2021, 12, 796704. [Google Scholar] [CrossRef]
- David, S.; Kroner, A. Repertoire of Microglial and Macrophage Responses after Spinal Cord Injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef]
- Magaki, S.D.; Williams, C.K.; Vinters, H. V Glial Function (and Dysfunction) in the Normal & Ischemic Brain. Neuropharmacology 2018, 134, 218–225. [Google Scholar] [CrossRef]
- Alizadeh, A.; Karimi-Abdolrezaee, S. Microenvironmental Regulation of Oligodendrocyte Replacement and Remyelination in Spinal Cord Injury. J. Physiol. 2016, 594, 3539–3552. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, O.N. Post-Traumatic Inflammation Following Spinal Cord Injury. Spinal Cord 2003, 41, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hu, B.; Wong, W.M.; Lu, P.; Wu, W.; Xu, X.-M. Glial and Axonal Responses in Areas of Wallerian Degeneration of the Corticospinal and Dorsal Ascending Tracts after Spinal Cord Dorsal Funiculotomy. Neuropathology 2009, 29, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, L.H.; Fehlings, M.G. Epidemiology, Demographics, and Pathophysiology of Acute Spinal Cord Injury. Spine 2001, 26, S2–S12. [Google Scholar] [CrossRef] [PubMed]
- Gwak, Y.S.; Kang, J.; Unabia, G.C.; Hulsebosch, C.E. Spatial and Temporal Activation of Spinal Glial Cells: Role of Gliopathy in Central Neuropathic Pain Following Spinal Cord Injury in Rats. Exp. Neurol. 2012, 234, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Chadi, G.; Andrade, M.S.; Leme, R.J.; Gomide, V.C. Experimental Models of Partial Lesion of Rat Spinal Cord to Investigate Neurodegeneration, Glial Activation, and Behavior Impairments. Int. J. Neurosci. 2001, 111, 137–165. [Google Scholar] [CrossRef]
- Leme, R.J.; Chadi, G. Distant Microglial and Astroglial Activation Secondary to Experimental Spinal Cord Lesion. Arq Neuropsiquiatr 2001, 59, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Abdanipour, A.; Tiraihi, T.; Taheri, T.; Kazemi, H. Microglial Activation in Rat Experimental Spinal Cord Injury Model. Iran. Biomed. J. 2013, 17, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.C.; Goetz, B.D.; Carré, J.L.; Duncan, I.D. Reactive Microglia in Dysmyelination and Demyelination. Glia 2001, 34, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Yu, D.; Bi, Y.; Cao, Y. A Disintegrin and Metalloprotease 17 Promotes Microglial Cell Survival via Epidermal Growth Factor Receptor Signalling Following Spinal Cord Injury. Mol. Med. Rep. 2015, 12, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting Neurotrophins to Specific Populations of Neurons: NGF, BDNF, and NT-3 and Their Relevance for Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef] [Green Version]
- Neumann, H.; Misgeld, T.; Matsumuro, K.; Wekerle, H. Neurotrophins Inhibit Major Histocompatibility Class II Inducibility of Microglia: Involvement of the P75 Neurotrophin Receptor. Proc. Natl. Acad. Sci. USA 1998, 95, 5779–5784. [Google Scholar] [CrossRef] [Green Version]
- Bessis, A.; Béchade, C.; Bernard, D.; Roumier, A. Microglial Control of Neuronal Death and Synaptic Properties. Glia 2007, 55, 233–238. [Google Scholar] [CrossRef]
- Tian, L.; Ma, L.; Kaarela, T.; Li, Z. Neuroimmune Crosstalk in the Central Nervous System and Its Significance for Neurological Diseases. J. Neuroinflammation 2012, 9, 155. [Google Scholar] [CrossRef] [Green Version]
- Manthou, M.; Abdulla, D.S.Y.; Pavlov, S.P.; Jansen, R.; Bendella, H.; Nohroudi, K.; Stein, G.; Meyer, C.; Ozsoy, O.; Ozsoy, U.; et al. Whole Body Vibration (WBV) Following Spinal Cord Injury (SCI) in Rats: Timing of Intervention. Restor. Neurol. Neurosci. 2017, 35, 185–216. [Google Scholar] [CrossRef] [PubMed]
- Ozsoy, O.; Ozsoy, U.; Stein, G.; Semler, O.; Skouras, E.; Schempf, G.; Wellmann, K.; Wirth, F.; Angelova, S.; Ankerne, J.; et al. Functional Deficits and Morphological Changes in the Neurogenic Bladder Match the Severity of Spinal Cord Compression. Restor. Neurol. Neurosci. 2012, 30, 363–381. [Google Scholar] [CrossRef] [PubMed]
- Semler, J.; Wellmann, K.; Wirth, F.; Stein, G.; Angelova, S.; Ashrafi, M.; Schempf, G.; Ankerne, J.; Ozsoy, O.; Ozsoy, U.; et al. Objective Measures of Motor Dysfunction after Compression Spinal Cord Injury in Adult Rats: Correlations with Locomotor Rating Scores. J. Neurotrauma 2011, 28, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Curtis, R.; Green, D.; Lindsay, R.M.; Wilkin, G.P. Up-Regulation of GAP-43 and Growth of Axons in Rat Spinal Cord after Compression Injury. J. Neurocytol. 1993, 22, 51–64. [Google Scholar] [CrossRef]
- Apostolova, I.; Irintchev, A.; Schachner, M. Tenascin-R Restricts Posttraumatic Remodeling of Motoneuron Innervation and Functional Recovery after Spinal Cord Injury in Adult Mice. J. Neurosci. 2006, 26, 7849–7859. [Google Scholar] [CrossRef] [Green Version]
- Manthou, M.; Nohroudi, K.; Moscarino, S.; Rehberg, F.; Stein, G.; Jansen, R.; Abdulla, D.; Jaminet, P.; Semler, O.; Schoenau, E.; et al. Functional Recovery after Experimental Spinal Cord Compression and Whole Body Vibration Therapy Requires a Balanced Revascularization of the Injured Site. Restor. Neurol. Neurosci. 2015, 33, 233–249. [Google Scholar] [CrossRef]
- Wirth, F.; Schempf, G.; Stein, G.; Wellmann, K.; Manthou, M.; Scholl, C.; Sidorenko, M.; Semler, O.; Eisel, L.; Harrach, R.; et al. Whole-Body Vibration Improves Functional Recovery in Spinal Cord Injured Rats. J. Neurotrauma 2013, 30, 453–468. [Google Scholar] [CrossRef]
- Ganor, I.; Golani, I. Coordination and Integration in the Hindleg Step Cycle of the Rat: Kinematic Synergies. Brain Res. 1980, 195, 57–67. [Google Scholar] [CrossRef]
- Stasiak, M.E.; Wiznia, D.; Alzoobaee, S.; Ciccotti, M.C.; Imhauser, C.W.; Voigt, C.; Torzilli, P.A.; Deng, X.-H.; Rodeo, S.A. A Novel Device to Apply Controlled Flexion and Extension to the Rat Knee Following Anterior Cruciate Ligament Reconstruction. J. Biomech. Eng. 2012, 134, 41008. [Google Scholar] [CrossRef] [PubMed]
- Hubli, M.; Dietz, V. The Physiological Basis of Neurorehabilitation--Locomotor Training after Spinal Cord Injury. J. NeuroEngineering Rehabil. 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, K.; Kerasidis, H.; Wrathall, J.R. Spinal Cord Contusion in the Rat: Behavioral Analysis of Functional Neurologic Impairment. Exp. Neurol. 1985, 88, 123–134. [Google Scholar] [CrossRef]
- Kerasidis, H.; Wrathall, J.R.; Gale, K. Behavioral Assessment of Functional Deficit in Rats with Contusive Spinal Cord Injury. J. Neurosci. Methods 1987, 20, 167–179. [Google Scholar] [CrossRef]
- Kim, Y.; Youm, Y.; Wu, M.; Schmit, B.D. Modulation of Flexor Reflexes by Static and Dynamic Hip Proprioceptors in Chronic Human Spinal Cord Injury. J. Clin. Neurosci. 2007, 14, 1078–1088. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, K.; Imai, Y.; Kanazawa, H.; Sasaki, Y.; Kohsaka, S. Involvement of Iba1 in Membrane Ruffling and Phagocytosis of Macrophages/Microglia. J. Cell Sci. 2000, 113 Pt 1, 3073–3084. [Google Scholar] [CrossRef]
- Ohsawa, K.; Imai, Y.; Sasaki, Y.; Kohsaka, S. Microglia/Macrophage-Specific Protein Iba1 Binds to Fimbrin and Enhances Its Actin-Bundling Activity. J. Neurochem. 2004, 88, 844–856. [Google Scholar] [CrossRef]
- Ahmed, Z.; Shaw, G.; Sharma, V.P.; Yang, C.; McGowan, E.; Dickson, D.W. Actin-Binding Proteins Coronin-1a and IBA-1 Are Effective Microglial Markers for Immunohistochemistry. J. Histochem. Cytochem. 2007, 55, 687–700. [Google Scholar] [CrossRef]
- Venkatesan, C.; Chrzaszcz, M.; Choi, N.; Wainwright, M.S. Chronic Upregulation of Activated Microglia Immunoreactive for Galectin-3/Mac-2 and Nerve Growth Factor Following Diffuse Axonal Injury. J. Neuroinflammation 2010, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Bellver-Landete, V.; Bretheau, F.; Mailhot, B.; Vallières, N.; Lessard, M.; Janelle, M.-E.; Vernoux, N.; Tremblay, M.-È.; Fuehrmann, T.; Shoichet, M.S.; et al. Microglia Are an Essential Component of the Neuroprotective Scar That Forms after Spinal Cord Injury. Nat. Commun. 2019, 10, 518. [Google Scholar] [CrossRef] [Green Version]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of Two Distinct Macrophage Subsets with Divergent Effects Causing Either Neurotoxicity or Regeneration in the Injured Mouse Spinal Cord. J. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponomarev, E.D.; Maresz, K.; Tan, Y.; Dittel, B.N. CNS-Derived Interleukin-4 Is Essential for the Regulation of Autoimmune Inflammation and Induces a State of Alternative Activation in Microglial Cells. J. Neurosci. 2007, 27, 10714–10721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mautes, A.E.; Weinzierl, M.R.; Donovan, F.; Noble, L.J. Vascular Events after Spinal Cord Injury: Contribution to Secondary Pathogenesis. Phys. Ther. 2000, 80, 673–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whetstone, W.D.; Hsu, J.-Y.C.; Eisenberg, M.; Werb, Z.; Noble-Haeusslein, L.J. Blood-Spinal Cord Barrier after Spinal Cord Injury: Relation to Revascularization and Wound Healing. J. Neurosci. Res. 2003, 74, 227–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noble, L.J.; Wrathall, J.R. Distribution and Time Course of Protein Extravasation in the Rat Spinal Cord after Contusive Injury. Brain Res. 1989, 482, 57–66. [Google Scholar] [CrossRef]
- Popovich, P.G.; Horner, P.J.; Mullin, B.B.; Stokes, B.T. A Quantitative Spatial Analysis of the Blood-Spinal Cord Barrier. I. Permeability Changes after Experimental Spinal Contusion Injury. Exp. Neurol. 1996, 142, 258–275. [Google Scholar] [CrossRef]
- Beck, K.D.; Nguyen, H.X.; Galvan, M.D.; Salazar, D.L.; Woodruff, T.M.; Anderson, A.J. Quantitative Analysis of Cellular Inflammation after Traumatic Spinal Cord Injury: Evidence for a Multiphasic Inflammatory Response in the Acute to Chronic Environment. Brain 2010, 133, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Ernfors, P.; Lee, K.F.; Kucera, J.; Jaenisch, R. Lack of Neurotrophin-3 Leads to Deficiencies in the Peripheral Nervous System and Loss of Limb Proprioceptive Afferents. Cell 1994, 77, 503–512. [Google Scholar] [CrossRef]
- Tessarollo, L.; Vogel, K.S.; Palko, M.E.; Reid, S.W.; Parada, L.F. Targeted Mutation in the Neurotrophin-3 Gene Results in Loss of Muscle Sensory Neurons. Proc. Natl. Acad. Sci. USA 1994, 91, 11844–11848. [Google Scholar] [CrossRef] [Green Version]
- Elkabes, S.; DiCicco-Bloom, E.M.; Black, I.B. Brain Microglia/Macrophages Express Neurotrophins That Selectively Regulate Microglial Proliferation and Function. J. Neurosci. 1996, 16, 2508–2521. [Google Scholar] [CrossRef] [Green Version]
- Elkabes, S.; Peng, L.; Black, I.B. Lipopolysaccharide Differentially Regulates Microglial Trk Receptor and Neurotrophin Expression. J. Neurosci. Res. 1998, 54, 117–122. [Google Scholar] [CrossRef]
- Ye, J.H.; Houle, J.D. Treatment of the Chronically Injured Spinal Cord with Neurotrophic Factors Can Promote Axonal Regeneration from Supraspinal Neurons. Exp. Neurol. 1997, 143, 70–81. [Google Scholar] [CrossRef]
- Coumans, J.V.; Lin, T.T.; Dai, H.N.; MacArthur, L.; McAtee, M.; Nash, C.; Bregman, B.S. Axonal Regeneration and Functional Recovery after Complete Spinal Cord Transection in Rats by Delayed Treatment with Transplants and Neurotrophins. J. Neurosci. 2001, 21, 9334–9344. [Google Scholar] [CrossRef] [Green Version]
- Narazaki, D.K.; de Barros Filho, T.E.P.; de Oliveira, C.R.G.C.M.; Cristante, A.F.; Iutaka, A.S.; Marcon, R.M.; Oliveira, R.P. Spinal Cord Regeneration: The Action of Neurotrophin-3 in Spinal Cord Injury in Rats. Clinics 2006, 61, 453–460. [Google Scholar] [CrossRef] [Green Version]
- Fitch, M.T.; Silver, J. CNS Injury, Glial Scars, and Inflammation: Inhibitory Extracellular Matrices and Regeneration Failure. Exp. Neurol. 2008, 209, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Mendell, L.M.; Munson, J.B.; Arvanian, V.L. Neurotrophins and Synaptic Plasticity in the Mammalian Spinal Cord. J. Physiol. 2001, 533, 91–97. [Google Scholar] [CrossRef]
- Arvanian, V. Role of Neurotrophins in Spinal Plasticity and Locomotion. Curr. Pharm. Des. 2013, 19, 4509–4516. [Google Scholar] [CrossRef]
- Kettenmann, H.; Kirchhoff, F.; Verkhratsky, A. Microglia: New Roles for the Synaptic Stripper. Neuron 2013, 77, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Pinilla, F.; Ying, Z.; Opazo, P.; Roy, R.R.; Edgerton, V.R. Differential Regulation by Exercise of BDNF and NT-3 in Rat Spinal Cord and Skeletal Muscle. Eur. J. Neurosci. 2001, 13, 1078–1084. [Google Scholar] [CrossRef]
- Hutchinson, K.J.; Gómez-Pinilla, F.; Crowe, M.J.; Ying, Z.; Basso, D.M. Three Exercise Paradigms Differentially Improve Sensory Recovery after Spinal Cord Contusion in Rats. Brain 2004, 127, 1403–1414. [Google Scholar] [CrossRef] [Green Version]
- Côté, M.-P.; Azzam, G.A.; Lemay, M.A.; Zhukareva, V.; Houlé, J.D. Activity-Dependent Increase in Neurotrophic Factors Is Associated with an Enhanced Modulation of Spinal Reflexes after Spinal Cord Injury. J. Neurotrauma 2011, 28, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Ying, Z.; Roy, R.R.; Edgerton, V.R.; Gómez-Pinilla, F. Voluntary Exercise Increases Neurotrophin-3 and Its Receptor TrkC in the Spinal Cord. Brain Res. 2003, 987, 93–99. [Google Scholar] [CrossRef]
- Donaghue, I.; Tator, C.; Shoichet, M. Sustained Delivery of Bioactive Neurotrophin-3 to the Injured Spinal Cord. Biomater. Sci. 2014, 3, 65–72. [Google Scholar] [CrossRef]
- Zhou, L.; Baumgartner, B.J.; Hill-Felberg, S.J.; McGowen, L.R.; Shine, H.D. Neurotrophin-3 Expressed in Situ Induces Axonal Plasticity in the Adult Injured Spinal Cord. J. Neurosci. 2003, 23, 1424–1431. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Chung, W.; Lee, J.-H.; Chung, D.-J.; Yang, W.-J.; Lee, A.-J.; Choi, C.-B.; Chang, H.-S.; Kim, D.-H.; Suh, H.J.; et al. Expression of Neurotrophic Factors in Injured Spinal Cord after Transplantation of Human-Umbilical Cord Blood Stem Cells in Rats. J. Vet. Sci. 2016, 17, 97–102. [Google Scholar] [CrossRef]
- Helgren, M.E.; Cliffer, K.D.; Torrento, K.; Cavnor, C.; Curtis, R.; DiStefano, P.S.; Wiegand, S.J.; Lindsay, R.M. Neurotrophin-3 Administration Attenuates Deficits of Pyridoxine-Induced Large-Fiber Sensory Neuropathy. J. Neurosci. 1997, 17, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Gao, W.Q.; Dybdal, N.; Shinsky, N.; Murnane, A.; Schmelzer, C.; Siegel, M.; Keller, G.; Hefti, F.; Phillips, H.S.; Winslow, J.W. Neurotrophin-3 Reverses Experimental Cisplatin-Induced Peripheral Sensory Neuropathy. Ann. Neurol. 1995, 38, 30–37. [Google Scholar] [CrossRef]
- Sahenk, Z.; Nagaraja, H.N.; McCracken, B.S.; King, W.M.; Freimer, M.L.; Cedarbaum, J.M.; Mendell, J.R. NT-3 Promotes Nerve Regeneration and Sensory Improvement in CMT1A Mouse Models and in Patients. Neurology 2005, 65, 681–689. [Google Scholar] [CrossRef]
- Ying, Z.; Roy, R.R.; Edgerton, V.R.; Gómez-Pinilla, F. Exercise Restores Levels of Neurotrophins and Synaptic Plasticity Following Spinal Cord Injury. Exp. Neurol. 2005, 193, 411–419. [Google Scholar] [CrossRef]
- Bouyer, L.J.G.; Rossignol, S. Contribution of Cutaneous Inputs from the Hindpaw to the Control of Locomotion. II. Spinal Cats. J. Neurophysiol. 2003, 90, 3640–3653. [Google Scholar] [CrossRef]
- Dietz, V.; Harkema, S.J. Locomotor Activity in Spinal Cord-Injured Persons. J. Appl. Physiol. 2004, 96, 1954–1960. [Google Scholar] [CrossRef]
- Kuerzi, J.; Brown, E.H.; Shum-Siu, A.; Siu, A.; Burke, D.; Morehouse, J.; Smith, R.R.; Magnuson, D.S.K. Task-Specificity vs. Ceiling Effect: Step-Training in Shallow Water after Spinal Cord Injury. Exp. Neurol. 2010, 224, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.R.; Shum-Siu, A.; Baltzley, R.; Bunger, M.; Baldini, A.; Burke, D.A.; Magnuson, D.S.K. Effects of Swimming on Functional Recovery after Incomplete Spinal Cord Injury in Rats. J. Neurotrauma 2006, 23, 908–919. [Google Scholar] [CrossRef]
- Roll, J.P.; Vedel, J.P.; Ribot, E. Alteration of Proprioceptive Messages Induced by Tendon Vibration in Man: A Microneurographic Study. Exp. Brain Res. 1989, 76, 213–222. [Google Scholar] [CrossRef]
- Bove, M.; Nardone, A.; Schieppati, M. Effects of Leg Muscle Tendon Vibration on Group Ia and Group II Reflex Responses to Stance Perturbation in Humans. J. Physiol. 2003, 550, 617–630. [Google Scholar] [CrossRef]
- Cardinale, M.; Bosco, C. The Use of Vibration as an Exercise Intervention. Exerc. Sport Sci. Rev. 2003, 31, 3–7. [Google Scholar] [CrossRef]
- Cardinale, M.; Lim, J. Electromyography Activity of Vastus Lateralis Muscle during Whole-Body Vibrations of Different Frequencies. J. Strength Cond. Res. 2003, 17, 621–624. [Google Scholar]
- Cardinale, M.; Rittweger, J. Vibration Exercise Makes Your Muscles and Bones Stronger: Fact or Fiction? J. Br. Menopause Soc. 2006, 12, 12–18. [Google Scholar] [CrossRef]
- Behrman, A.L.; Harkema, S.J. Locomotor Training after Human Spinal Cord Injury: A Series of Case Studies. Phys. Ther. 2000, 80, 688–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, V.; Müller, R.; Colombo, G. Locomotor Activity in Spinal Man: Significance of Afferent Input from Joint and Load Receptors. Brain 2002, 125, 2626–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Day 0 | Day 7 | Day 14 | Day 28 | Day 90 |
|---|---|---|---|---|---|
| Intact | endpoint | ||||
| SCI | endpoint | ||||
| SCI | WBV | endpoint | |||
| SCI | WBV | endpoint | |||
| SCI | WBV | endpoint | |||
| SCI | PFE | endpoint |
| Group | 0 Weeks after SCI | 1 Week after SCI | 3 Weeks after SCI | 6 Weeks after SCI | 9 Weeks after SCI | 12 Weeks after SCI |
|---|---|---|---|---|---|---|
| SCI-noEx | 18 ± 1.0 | 130 ± 20 | 97 ± 47 | 88 ± 41 | 72 ± 41 | 66 ± 42 |
| WBV7 | 19 ± 1.0 | 123 ± 22 | 103 ± 43 | 93 ± 47 | 85 ± 40 | 93 ± 35 |
| WBV14 | 18 ± 1.0 | 98 ± 27 | 28 ± 21 | 28 ± 24 | 26 ± 11 | 26 ± 11 |
| WBV28 | 17 ± 1.6 | 114 ± 49 | 111 ± 24 | 86 ± 38 | 103 ± 39 | 118 ± 29 |
| ML (PFE) | 16 ± 1.0 | 145 ± 6 | 132 ± 12 | 106± 27 | 103 ± 22 | 105 ± 36 |
| Group | Iba1_Ventral Horn | Iba1_Dorsal Horn |
|---|---|---|
| Intact (n = 3) | 12,263 ± 1530 | 13,194 ± 3113 |
| SCI-noEx (n = 5) | 4535 ± 4234 | 3868 ±2959 |
| WBV7 (n = 5) | 12,211 ± 4467 | 5593 ± 2541 |
| WBV14 (n = 9) | 6477 ± 2800 | 5317 ± 2739 |
| WBV28 (n = 6) | 22,645 ± 11,121 * | 12,924 ± 5657 |
| ML (PFE) (n = 7) | 12,229 ± 6923 | 7593 ± 4133 |
| Group | NT-3_Ventral Horn | NT-3_Dorsal Horn |
|---|---|---|
| Intact (n = 3) | 162,483± 9224 | 481,178 ± 27,187 |
| SCI-noEx (n = 5) | 176,548 ± 26,117 | 505,795 ± 53,792 |
| WBV7 (n = 5) | 334,924 ± 51,345 | 595,022 ± 61,196 |
| WBV14 (n = 9) | 525,418 ± 197885 * | 739,444 ± 90,572 * |
| WBV28 (n = 6) | 318,238 ± 61485 | 644,613 ± 95,059 |
| ML (PFE) (n = 7) | 388,171 ± 141,477 | 624,435 ± 97,267 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schaufler, D.; Manthou, M.E.; Theotokis, P.; Rink-Notzon, S.; Angelov, D.N. Effects of Whole-Body Vibration and Manually Assisted Locomotor Therapy on Neurotrophin-3 Expression and Microglia/Macrophage Mobilization Following Thoracic Spinal Cord Injury in Rats. Curr. Issues Mol. Biol. 2023, 45, 3238-3254. https://doi.org/10.3390/cimb45040211
Schaufler D, Manthou ME, Theotokis P, Rink-Notzon S, Angelov DN. Effects of Whole-Body Vibration and Manually Assisted Locomotor Therapy on Neurotrophin-3 Expression and Microglia/Macrophage Mobilization Following Thoracic Spinal Cord Injury in Rats. Current Issues in Molecular Biology. 2023; 45(4):3238-3254. https://doi.org/10.3390/cimb45040211
Chicago/Turabian StyleSchaufler, Diana, Maria Eleni Manthou, Paschalis Theotokis, Svenja Rink-Notzon, and Doychin N. Angelov. 2023. "Effects of Whole-Body Vibration and Manually Assisted Locomotor Therapy on Neurotrophin-3 Expression and Microglia/Macrophage Mobilization Following Thoracic Spinal Cord Injury in Rats" Current Issues in Molecular Biology 45, no. 4: 3238-3254. https://doi.org/10.3390/cimb45040211