
Knockdown of KDM5B Leads to DNA Damage and Cell Cycle Arrest in Granulosa Cells via MTF1
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Granulosa Cell Culture
2.3. Transfection of siRNA against KDM5B
2.4. Quantitative Real-Time PCR
2.5. Immunofluorescence Staining
2.6. Cell Proliferation Analysis
2.7. Flow Cytometry Analysis
2.8. Western Blot Analysis
2.9. Intracellular Reactive Oxygen Species Detection
2.10. Mitochondrial Membrane Potential Detection
2.11. RNA Sequence Data Analysis
2.12. GO, KEGG and GSEA Analysis
2.13. Statistical Analysis
3. Results
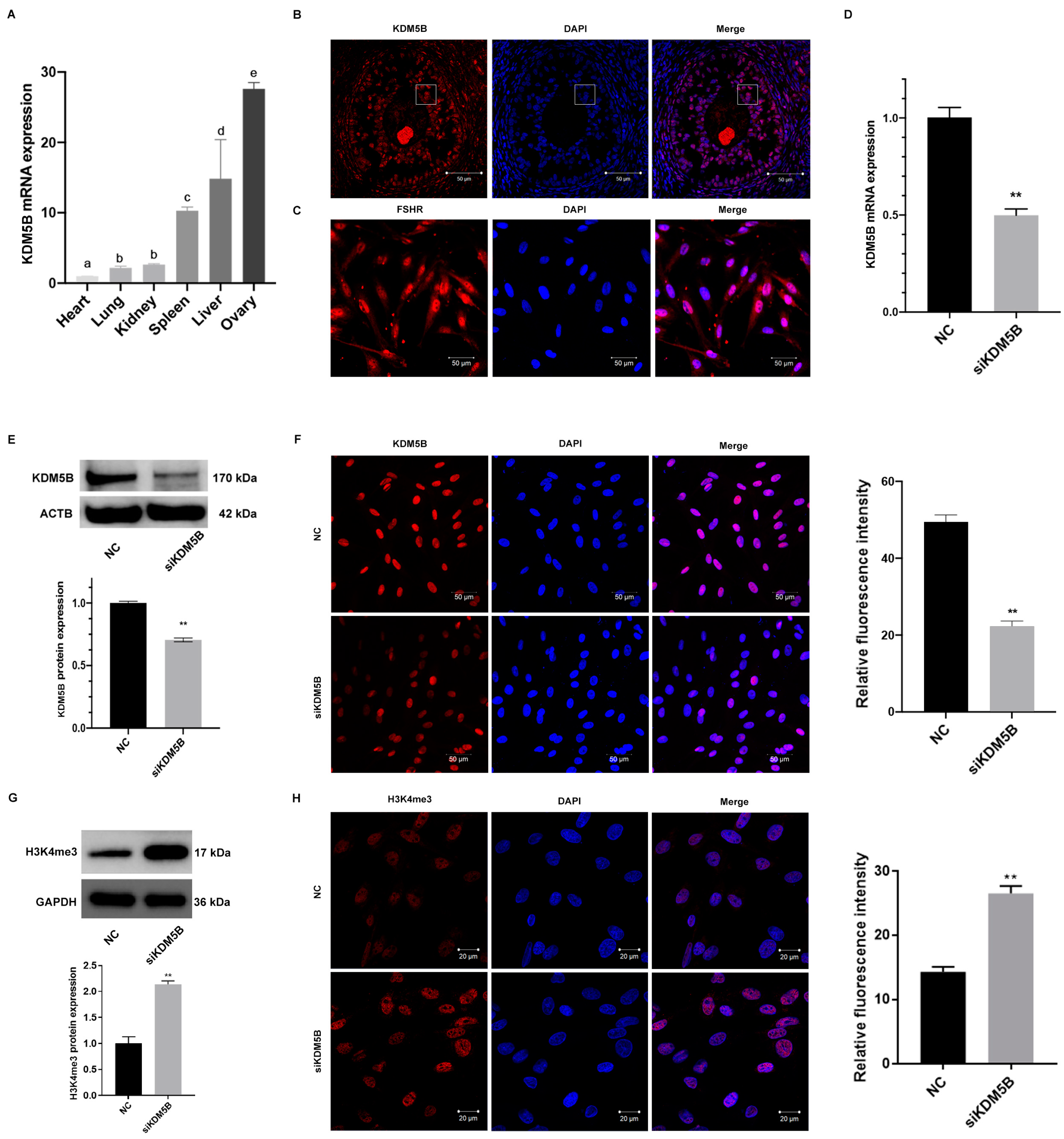
3.1. High Expression of KDM5B in Goat GCs
3.2. Knockdown of KDM5B in Goat GCs
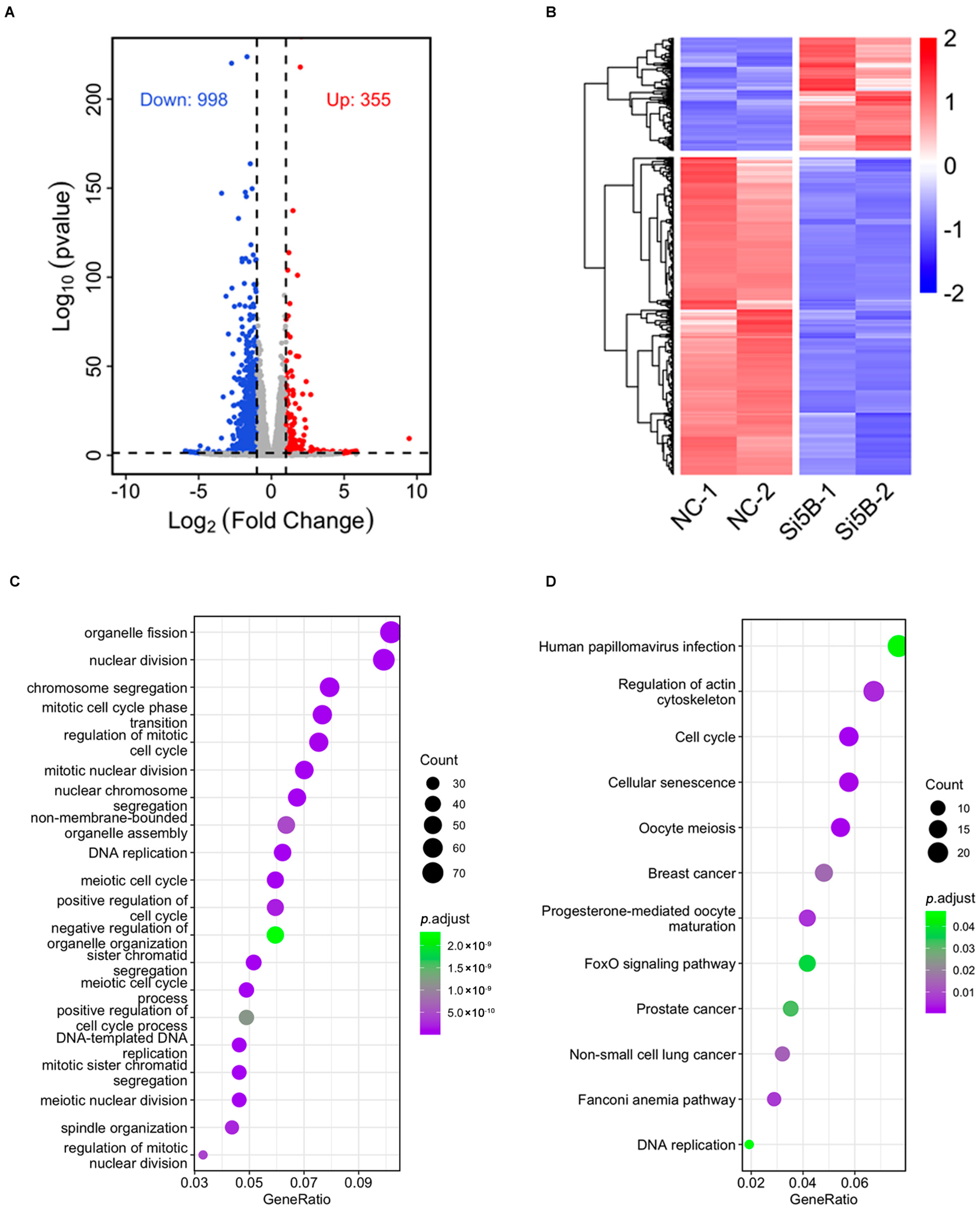
3.3. Compromise of Cell Cycle and DNA Replication-Related Functions in KDM5B Knockdown GCs
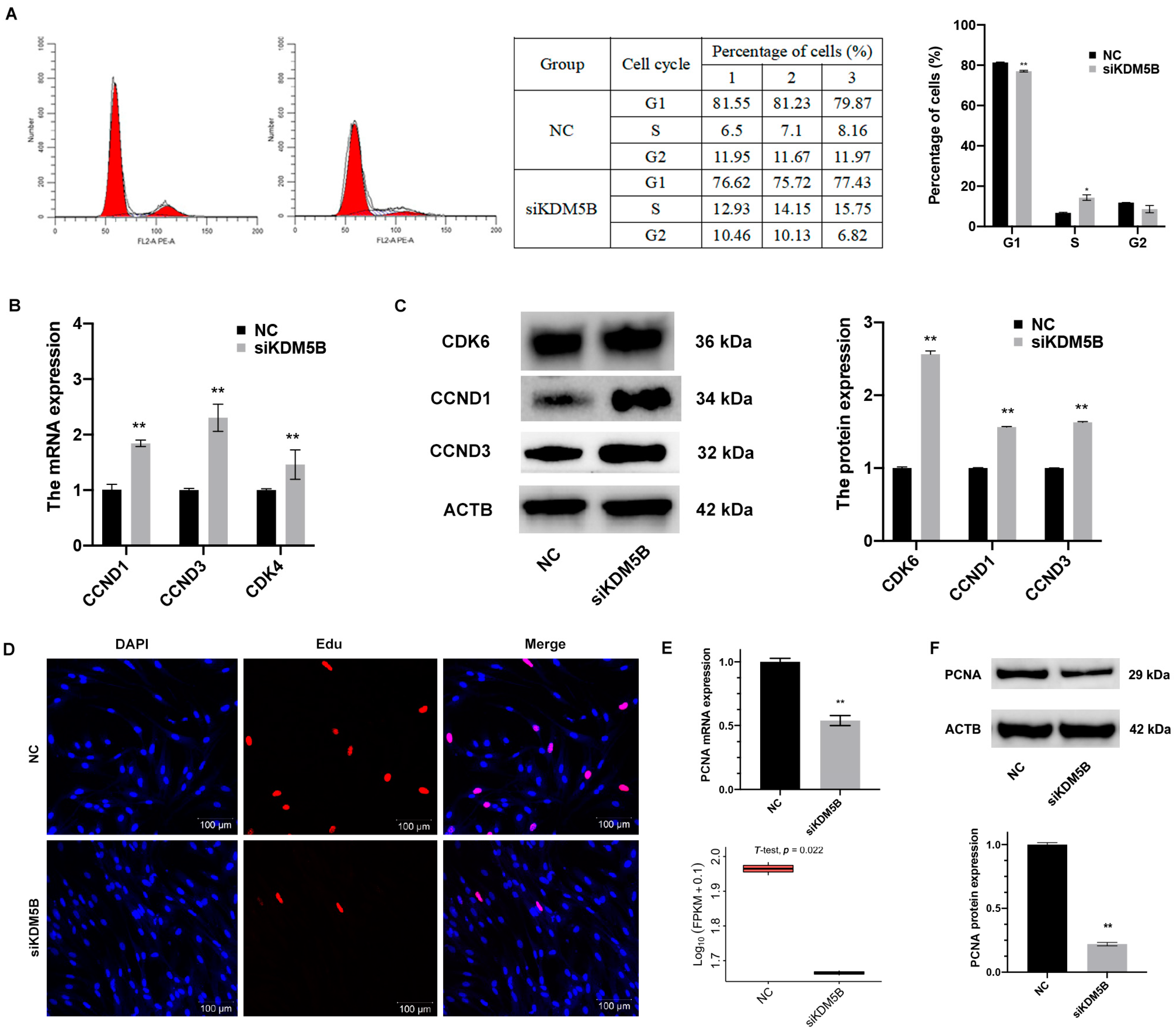
3.4. Blocked S Phase and Restrained Proliferation in KDM5B Knockdown GCs
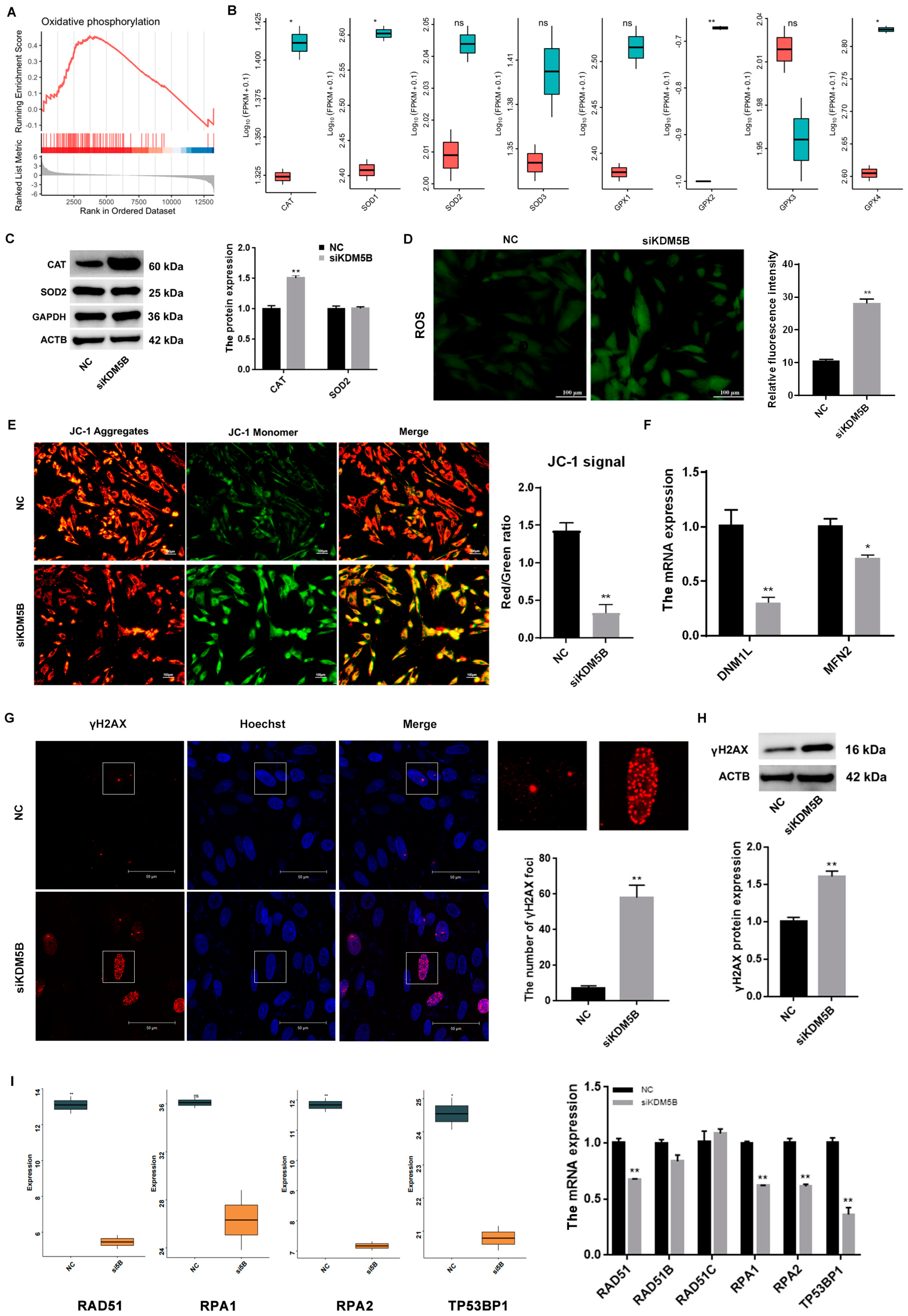
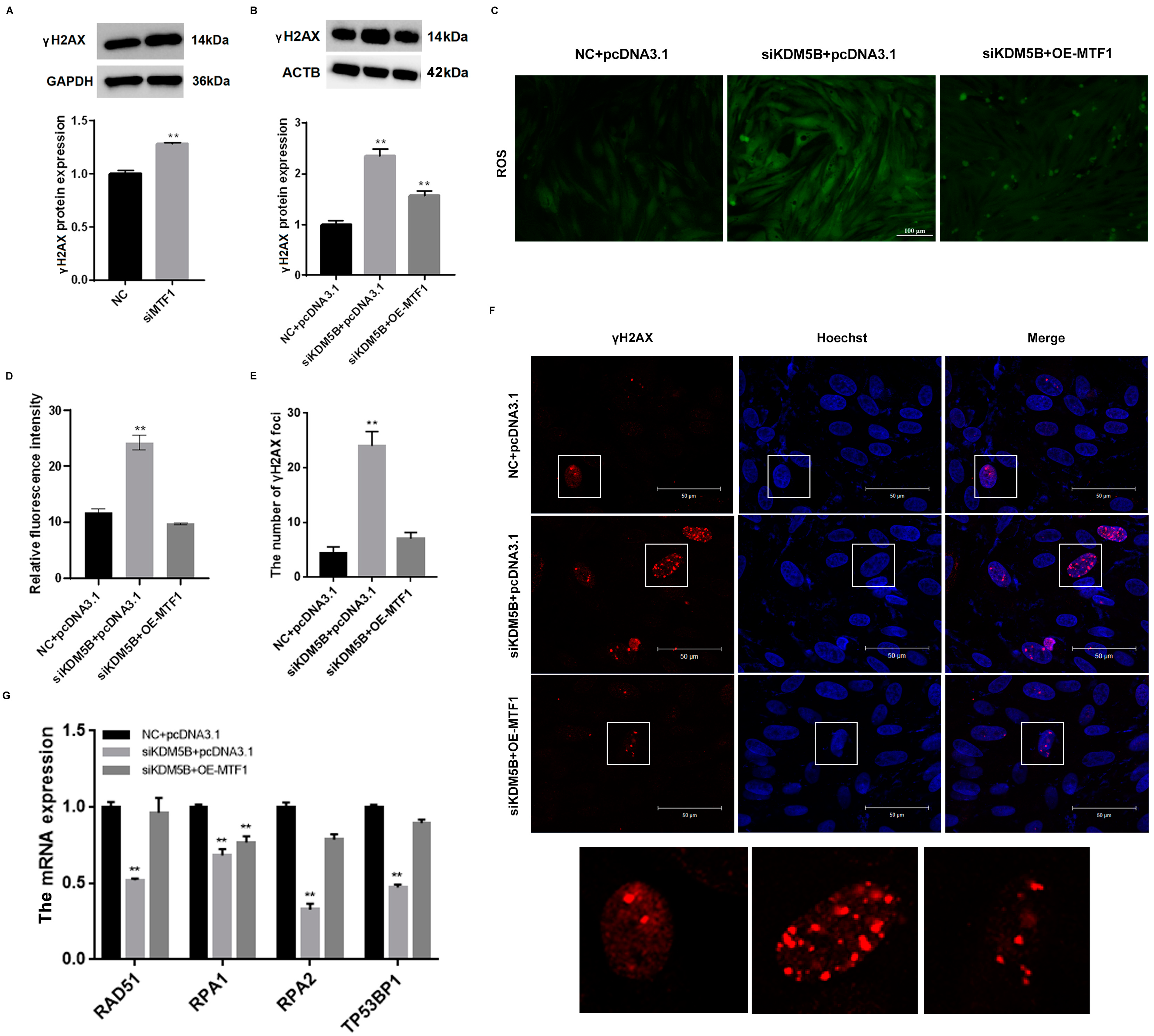
3.5. DNA DSBs in KDM5B Knockdown GCs
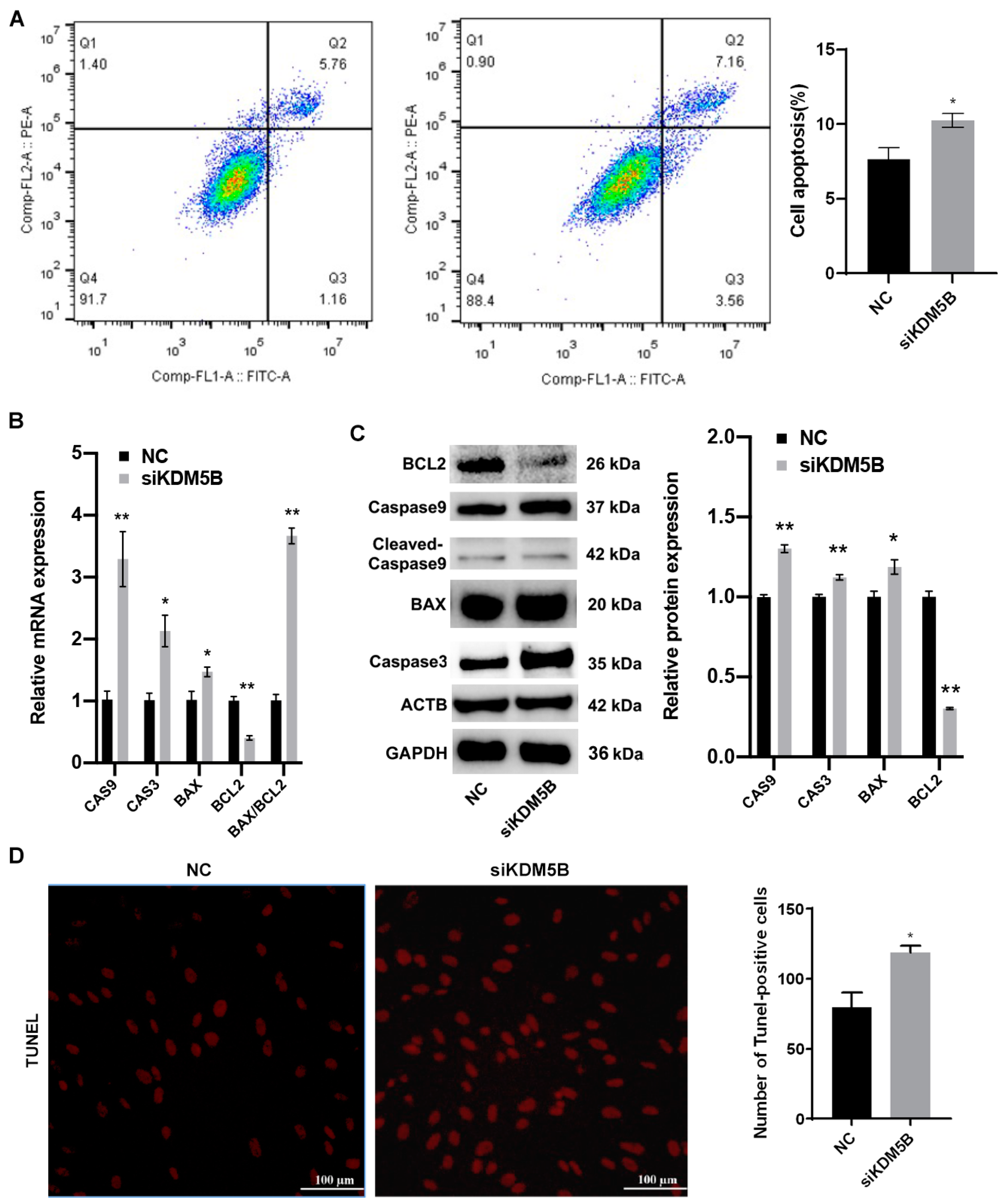
3.6. Promotion of Apoptosis in KDM5B Knockdown GCs
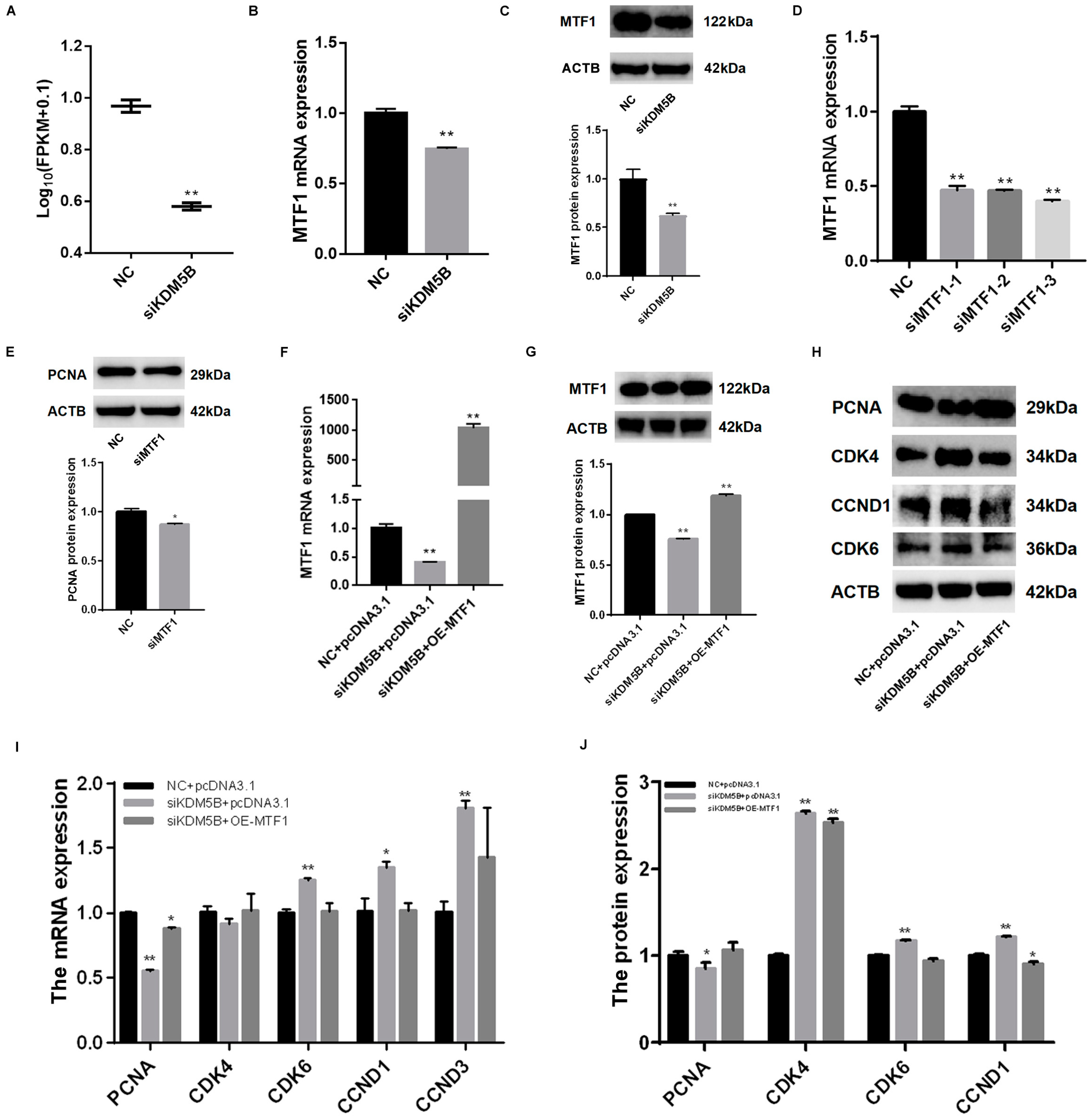
3.7. Rescued Cell Proliferation and Cell Cycle in KDM5B Knockdown and MTF1 Overexpression GCs
3.8. Decreased DNA Damage in KDM5B Knockdown and MTF1 Overexpression GCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Solovova, O.A.; Chernykh, V.B. Genetics of Oocyte Maturation Defects and Early Embryo Development Arrest. Genes 2022, 13, 1920. [Google Scholar] [CrossRef]
- Nivet, A.L.; Leveille, M.C.; Leader, A.; Sirard, M.A. Transcriptional characteristics of different sized follicles in relation to embryo transferability: Potential role of hepatocyte growth factor signalling. Mol. Hum. Reprod. 2016, 22, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, R.J.; Lavranos, T.C.; van Wezel, I.L.; Irving-Rodgers, H.F. Development of the ovarian follicular epithelium. Mol. Cell. Endocrinol. 1999, 151, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Safdar, M.; Liang, A.; Rajput, S.A.; Abbas, N.; Zubair, M.; Shaukat, A.; Rehman, A.U.; Jamil, H.; Guo, Y.; Ullah, F.; et al. Orexin-A Regulates Follicular Growth, Proliferation, Cell Cycle and Apoptosis in Mouse Primary Granulosa Cells via the AKT/ERK Signaling Pathway. Molecules 2021, 26, 5635. [Google Scholar] [CrossRef]
- Fan, Y.; Chang, Y.; Wei, L.; Chen, J.; Li, J.; Goldsmith, S.; Silber, S.; Liang, X. Apoptosis of mural granulosa cells is increased in women with diminished ovarian reserve. J. Assist. Reprod. Genet. 2019, 36, 1225–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, J.; Li, J.; Li, X.; Wang, X.; Xiao, Y.; Sun, Z.; Yu, Q. Downregulating lncRNA NEAT1 induces proliferation and represses apoptosis of ovarian granulosa cells in polycystic ovary syndrome via microRNA-381/IGF1 axis. J. Biomed. Sci. 2021, 28, 53. [Google Scholar] [CrossRef]
- Lee, J.H.; Berger, J.M. Cell Cycle-Dependent Control and Roles of DNA Topoisomerase II. Genes 2019, 10, 859. [Google Scholar] [CrossRef] [Green Version]
- Turrero García, M.; Chang, Y.; Arai, Y.; Huttner, W.B. S-phase duration is the main target of cell cycle regulation in neural progenitors of developing ferret neocortex. J. Comp. Neurol. 2016, 524, 456–470. [Google Scholar] [CrossRef] [Green Version]
- Neganova, I.; Lako, M. G1 to S phase cell cycle transition in somatic and embryonic stem cells. J. Anat. 2008, 213, 30–44. [Google Scholar] [CrossRef]
- Blackford, A.N.; Stucki, M. How Cells Respond to DNA Breaks in Mitosis. Trends Biochem. Sci. 2020, 45, 321–331. [Google Scholar] [CrossRef]
- Williams, G.H.; Stoeber, K. The cell cycle and cancer. J. Pathol. 2012, 226, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Hume, S.; Dianov, G.L.; Ramadan, K. A unified model for the G1/S cell cycle transition. Nucleic Acids Res. 2020, 48, 12483–12501. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.Y.; Lin, S.W.; Wu, Y.C.; Chan, N.L.; Chi, P. Functional characterization of the meiosis-specific DNA double-strand break inducing factor SPO-11 from C. elegans. Sci. Rep. 2017, 7, 2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takanami, T.; Mori, A.; Takahashi, H.; Horiuchi, S.; Higashitani, A. Caenorhabditis elegans Ce-rdh-1/rad-51 functions after double-strand break formation of meiotic recombination. Chromosome Res. 2003, 11, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Somfai, T.; Haraguchi, S.; Dang-Nguyen, T.Q.; Kaneko, H.; Kikuchi, K. Vitrification of porcine immature oocytes and zygotes results in different levels of DNA damage which reflects developmental competence to the blastocyst stage. PLoS ONE 2023, 18, e0282959. [Google Scholar] [CrossRef] [PubMed]
- Pedroza-Garcia, J.A.; Xiang, Y.; De Veylder, L. Cell cycle checkpoint control in response to DNA damage by environmental stresses. Plant J. 2022, 109, 490–507. [Google Scholar] [CrossRef] [PubMed]
- Ovejero-Sánchez, M.; Rubio-Heras, J.; Vicente de la Peña, M.D.C.; San-Segundo, L.; Pérez-Losada, J.; González-Sarmiento, R.; Herrero, A.B. Chloroquine-Induced DNA Damage Synergizes with Nonhomologous End Joining Inhibition to Cause Ovarian Cancer Cell Cytotoxicity. Int. J. Mol. Sci. 2022, 23, 7518. [Google Scholar] [CrossRef]
- Angelis, K.J.; Zaveska Drabkova, L.; Vagnerova, R.; Hola, M. RAD51 and RAD51B Play Diverse Roles in the Repair of DNA Double Strand Breaks in Physcomitrium patens. Genes 2023, 14, 305. [Google Scholar] [CrossRef]
- Liu, J.B.; Zhang, J.B.; Yan, X.M.; Xie, P.G.; Fu, Y.; Fu, X.H.; Sun, X.L.; Han, D.X.; Li, S.P.; Zheng, Y.; et al. DNA Double-Strand Break-Related Competitive Endogenous RNA Network of Noncoding RNA in Bovine Cumulus Cells. Genes 2023, 14, 290. [Google Scholar] [CrossRef]
- Ui, A.; Chiba, N.; Yasui, A. Relationship among DNA double-strand break (DSB), DSB repair, and transcription prevents genome instability and cancer. Cancer Sci. 2020, 111, 1443–1451. [Google Scholar] [CrossRef]
- Gong, Y.; Luo, S.; Fan, P.; Zhu, H.; Li, Y.; Huang, W. Growth hormone activates PI3K/Akt signaling and inhibits ROS accumulation and apoptosis in granulosa cells of patients with polycystic ovary syndrome. Reprod. Biol. Endocrinol. 2020, 18, 121. [Google Scholar] [CrossRef] [PubMed]
- Guru, A.; Sudhakaran, G.; Almutairi, M.H.; Almutairi, B.O.; Juliet, A.; Arockiaraj, J. β-cells regeneration by WL15 of cysteine and glycine-rich protein 2 which reduces alloxan induced β-cell dysfunction and oxidative stress through phosphoenolpyruvate carboxykinase and insulin pathway in zebrafish in-vivo larval model. Mol. Biol. Rep. 2022, 49, 11867–11879. [Google Scholar] [CrossRef] [PubMed]
- Forte, A.; Lessa, P.H.C.; Chaves Filho, A.J.M.; Aquino, P.E.A.; Brito, L.M.; Pinheiro, L.C.; Juruena, M.F.; Lucena, D.F.; de Rezende, P.H.F.; de Vasconcelos, S.M.M. Oxidative stress and inflammatory process in borderline personality disorder (BPD): A narrative review. Braz. J. Med. Biol. Res. 2023, 56, e12484. [Google Scholar] [CrossRef]
- Glanzner, W.G.; Gutierrez, K.; Rissi, V.B.; de Macedo, M.P.; Lopez, R.; Currin, L.; Dicks, N.; Baldassarre, H.; Agellon, L.B.; Bordignon, V. Histone Lysine Demethylases KDM5B and KDM5C Modulate Genome Activation and Stability in Porcine Embryos. Front. Cell Dev. Biol. 2020, 8, 151. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, H.; Wang, X.; Dobbs, K.B.; Yao, J.; Qin, G.; Whitworth, K.; Walters, E.M.; Prather, R.S.; Zhao, J. Impairment of preimplantation porcine embryo development by histone demethylase KDM5B knockdown through disturbance of bivalent H3K4me3-H3K27me3 modifications. Biol. Reprod. 2015, 92, 72. [Google Scholar] [CrossRef] [Green Version]
- Di Nisio, E.; Licursi, V.; Mannironi, C.; Buglioni, V.; Paiardini, A.; Robusti, G.; Noberini, R.; Bonaldi, T.; Negri, R. A truncated and catalytically inactive isoform of KDM5B histone demethylase accumulates in breast cancer cells and regulates H3K4 tri-methylation and gene expression. Cancer Gene Ther. 2023, 1–11. [Google Scholar] [CrossRef]
- Lu, P.J.; Sundquist, K.; Baeckstrom, D.; Poulsom, R.; Hanby, A.; Meier-Ewert, S.; Jones, T.; Mitchell, M.; Pitha-Rowe, P.; Freemont, P.; et al. A novel gene (PLU-1) containing highly conserved putative DNA/chromatin binding motifs is specifically up-regulated in breast cancer. J. Biol. Chem. 1999, 274, 15633–15645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayami, S.; Yoshimatsu, M.; Veerakumarasivam, A.; Unoki, M.; Iwai, Y.; Tsunoda, T.; Field, H.I.; Kelly, J.D.; Neal, D.E.; Yamaue, H.; et al. Overexpression of the JmjC histone demethylase KDM5B in human carcinogenesis: Involvement in the proliferation of cancer cells through the E2F/RB pathway. Mol. Cancer 2010, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Han, S.; Peng, R.; Jiao, C.; Wang, X.; Yang, X.; Yang, R.; Li, X. Depletion of histone demethylase KDM5B inhibits cell proliferation of hepatocellular carcinoma by regulation of cell cycle checkpoint proteins p15 and p27. J. Exp. Clin. Cancer Res. 2016, 35, 37. [Google Scholar] [CrossRef] [Green Version]
- Barrett, A.; Madsen, B.; Copier, J.; Lu, P.J.; Cooper, L.; Scibetta, A.G.; Burchell, J.; Taylor-Papadimitriou, J. PLU-1 nuclear protein, which is upregulated in breast cancer, shows restricted expression in normal human adult tissues: A new cancer/testis antigen? Int. J. Cancer 2002, 101, 581–588. [Google Scholar] [CrossRef]
- McCann, C.; Quinteros, M.; Adelugba, I.; Morgada, M.N.; Castelblanco, A.R.; Davis, E.J.; Lanzirotti, A.; Hainer, S.J.; Vila, A.J.; Navea, J.G.; et al. The mitochondrial Cu(+) transporter PiC2 (SLC25A3) is a target of MTF1 and contributes to the development of skeletal muscle in vitro. Front. Mol. Biosci. 2022, 9, 1037941. [Google Scholar] [CrossRef]
- Bi, S.S.; Talukder, M.; Jin, H.T.; Lv, M.W.; Ge, J.; Zhang, C.; Li, J.L. Nano-selenium alleviates cadmium-induced cerebellar injury by activating metal regulatory transcription factor 1 mediated metal response. Anim. Nutr. 2022, 11, 402–412. [Google Scholar] [CrossRef]
- Deng, M.; Wan, Y.; Chen, B.; Dai, X.; Liu, Z.; Yang, Y.; Cai, Y.; Zhang, Y.; Wang, F. Long non-coding RNA lnc_3712 impedes nuclear reprogramming via repressing Kdm5b. Mol. Ther. Nucleic Acids 2021, 24, 54–66. [Google Scholar] [CrossRef]
- Sedelnikova, O.A.; Rogakou, E.P.; Panyutin, I.G.; Bonner, W.M. Quantitative detection of (125)IdU-induced DNA double-strand breaks with gamma-H2AX antibody. Radiat. Res. 2002, 158, 486–492. [Google Scholar] [CrossRef]
- Słupianek, A.; Pytel, D.; Majsterek, I. The role of oncogenic tyrosine kinases in the cellular response to anticancer therapy. Postepy Hig. Med. Dosw. 2007, 61, 819–827. [Google Scholar]
- Shahin, W.S.; Ebed, S.O.; Tyler, S.R.; Miljkovic, B.; Choi, S.H.; Zhang, Y.; Zhou, W.; Evans, I.A.; Yeaman, C.; Engelhardt, J.F. Redox-dependent Igfbp2 signaling controls Brca1 DNA damage response to govern neural stem cell fate. Nat. Commun. 2023, 14, 444. [Google Scholar] [CrossRef]
- Ovejero-Sanchez, M.; Gonzalez-Sarmiento, R.; Herrero, A.B. Synergistic effect of Chloroquine and Panobinostat in ovarian cancer through induction of DNA damage and inhibition of DNA repair. Neoplasia 2021, 23, 515–528. [Google Scholar] [CrossRef]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Hübner, C.; Haase, H. Interactions of zinc- and redox-signaling pathways. Redox Biol. 2021, 41, 101916. [Google Scholar] [CrossRef]
- Huang, S.; Cao, B.; Zhang, J.; Feng, Y.; Wang, L.; Chen, X.; Su, H.; Liao, S.; Liu, J.; Yan, J.; et al. Induction of ferroptosis in human nasopharyngeal cancer cells by cucurbitacin B: Molecular mechanism and therapeutic potential. Cell Death Dis. 2021, 12, 237. [Google Scholar] [CrossRef]
- Li, J.; Wan, X.; Qiang, W.; Li, T.; Huang, W.; Huang, S.; Wu, D.; Li, Y. MiR-29a suppresses prostate cell proliferation and induces apoptosis via KDM5B protein regulation. Int. J. Clin. Exp. Med. 2015, 8, 5329–5339. [Google Scholar]
- Yoo, J.; Kim, G.W.; Jeon, Y.H.; Kim, J.Y.; Lee, S.W.; Kwon, S.H. Drawing a line between histone demethylase KDM5A and KDM5B: Their roles in development and tumorigenesis. Exp. Mol. Med. 2022, 54, 2107–2117. [Google Scholar] [CrossRef]
- Cui, G.; Liu, D.; Li, W.; Li, Y.; Liang, Y.; Shi, W.; Zhao, S. Original Research: miR-194 inhibits proliferation and invasion and promotes apoptosis by targeting KDM5B in esophageal squamous cell carcinoma cells. Exp. Biol. Med. 2017, 242, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, W.; Wei, B.; Wu, K.; Liu, D.; Zhu, D.; Zhang, C.; Wen, F.; Fan, Y.; Zhao, S. MicroRNA let-7i Inhibits Histone Lysine Demethylase KDM5B to Halt Esophageal Cancer Progression. Mol. Ther. Nucleic Acids 2020, 22, 846–861. [Google Scholar] [CrossRef]
- Scully, R.; Panday, A.; Elango, R.; Willis, N.A. DNA double-strand break repair-pathway choice in somatic mammalian cells. Nat. Rev. Mol. Cell Biol. 2019, 20, 698–714. [Google Scholar] [CrossRef]
- Tarsounas, M.; Sung, P. The antitumorigenic roles of BRCA1-BARD1 in DNA repair and replication. Nat. Rev. Mol. Cell Biol. 2020, 21, 284–299. [Google Scholar] [CrossRef]
- Li, X.; Heyer, W.D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008, 18, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Awwad, S.W.; Darawshe, M.M.; Machour, F.E.; Arman, I.; Ayoub, N. Recruitment of RBM6 to DNA Double-Strand Breaks Fosters Homologous Recombination Repair. Mol. Cell. Biol. 2023, 43, 130–142. [Google Scholar] [CrossRef]
- Imamura, R.; Saito, M.; Shimada, M.; Kobayashi, J.; Ishiai, M.; Matsumoto, Y. APTX acts in DNA double-strand break repair in a manner distinct from XRCC4. J. Radiat. Res. 2023, rrad007. [Google Scholar] [CrossRef]
- Gale, M.; Sayegh, J.; Cao, J.; Norcia, M.; Gareiss, P.; Hoyer, D.; Merkel, J.S.; Yan, Q. Screen-identified selective inhibitor of lysine demethylase 5A blocks cancer cell growth and drug resistance. Oncotarget 2016, 7, 39931–39944. [Google Scholar] [CrossRef] [Green Version]
- Bonacci, T.; Emanuele, M.J. Dissenting degradation: Deubiquitinases in cell cycle and cancer. Semin. Cancer Biol. 2020, 67, 145–158. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, F.; Qi, G.; Yuan, S.; Zhang, G.; Tang, B.; He, S. KDM5B is overexpressed in gastric cancer and is required for gastric cancer cell proliferation and metastasis. Am. J. Cancer Res. 2015, 5, 87–100. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sense (5′-3′) |
|---|---|
| Negative control (NC) | S:5′-UUCUCCGAACGUGUCACGUTT-3′ A:5′-ACGUGACACGUUCGGAGAATT-3′ |
| KDM5B-goat | S:5′-GCGACUAGUGAGCACUAUUTT-3′ A:5′-AAUAGUGCUCACUAGUCGCTT-3′ |
| MTF1-goat | S:5′-CUCUGUCCUAACUAGUAAUTT-3′ A:5′-AUUACUAGUUAGGACAGAGTT-3′ |
| Gene | Primer Sequence (5′-3′) | Product Size (bp) | Accession Number |
|---|---|---|---|
| KDM5B | F 5′-TCCTTGAGATGGCAGCACTC-3′ R 5′-CCTTCTGACACACGCAGACT-3′ | 134 | XM_018060847.1 |
| PCNA | F 5′-TGGCTCCCAAGATCGAGGAT-3′ R 5′-TAAAACTGCATTTAGAGTCAAGACC-3′ | 381 | NM_002592.2 |
| CASP3 | F 5′-TGAAACATGCCGCCTTCCTA-3′ R 5′-AGTGGCATACCCACATGACTG-3′ | 242 | XM_018041755.1 |
| CASP9 | F 5′-CCAGATGCCGTGTCTAGTCTG-3′ R 5′-ACAGTAAGGTAGGGTGAGGGG-3′ | 296 | XM_005690814.3 |
| BAX | F 5′-GCATCCACCAAGAAGCTGAG-3′ R 5′-CCGCCACTCGGAAAAAGAC-3′ | 130 | XM_002701934.1 |
| BCL2 | F 5′-ATGTGTGTGGAGAGCGTCA-3′ R 5′-AGAGACAGCCAGGAGAAATC-3′ | 182 | NM_001166486.1 |
| CDK4 | F 5′-CCTTCATGCCAACTGCATCG-3′ R 5′-GGCAGTCCGATCAGGTCAAA-3′ | 309 | XM_005680266.3 |
| Ccnd1 | F 5′-AGACCCTCGCTTGTGCTTAC-3′ R 5′-AACGTGCCGGTTACATGTCT-3′ | 117 | XM_018043271.1 |
| Ccne3 | F 5′-GGATGGAGCTGCTGTGCTG-3′ R 5′-ACAGCTTCTCGATGGTCAGG-3′ | 375 | XM_018038733.1 |
| 3β-HSD | F 5′-AGACCAGAAGTTCGGGAGGAA-3′ R 5′-TCTCCCTGTAGGAGTTGGGC-3′ | 292 | XM_013962473.2 |
| STAR | F 5′-AATCCACTTGGGTCTGCGAG-3′ R 5′-CACTTTCGCCACATCGAGAAC-3′ | 262 | XM_013975437.2 |
| CYP11 | F 5′-CACTTTCGCCACATCGAGAAC-3′ R 5′-AGGCTCCTGACTTCTTAAACAGG-3′ | 217 | NM_001287574.1 |
| CYP17 | F 5′-AACGCCATAGCAAGGAACGA-3′ R 5′-TCTGTCACGCTGTGTTGTGT-3′ | 263 | NM_001255003.3 |
| CYP19 | F 5′-TGGGCTATGTGGACGTGTTG-3′ R 5′-TTCACCACGTTTCTCGGCAA-3′ | 188 | NM_001123000.1 |
| ESR | F 5′-ATGCATCCAACACCAATGGC-3′ R 5′-TCTGAGCCCCAACCCATAGA-3′ | 213 | NM_000125.4 |
| ESRβ | F 5′-CTGCTGGCTTTTTGGACACC-3′ R 5′-CCGGCCTTGCCTTCTCTAAA-3′ | 297 | NM_000125.4 |
| PGR | F 5′-CAGCCAGAGCCCACAGTACA-3′ R 5′-TGCAATCGTTTCTTCCAGCA-3′ | 176 | XM_027979146.1 |
| FSHR | F 5′-TGAGCAAGTGTGGCTGCTAT-3′ R 5′-ATGTGTAGAAGCACTGTCAGC-3′ | 320 | NM_000145.4 |
| ACTB | F 5′-GGCCATGA-3′ R 5′-CCACGCTCCGTGAGAATCTT-3′ | 252 | NM_001314342.1 |
| Antibodies | Cat NO. | Source | Dilution | Observed Band (kDa) |
|---|---|---|---|---|
| Anti-KDM5B/PLU1/Jarid1B Antibody | Ab181089 | Abcam | 1:5000 | 170 |
| Anti-PCNA Antibody | Ab18197 | Abcam | 1:500 | 29 |
| BAX Rabbit Polyclonal Antibody | 50599-2-Ig | ProteinTech | 1:2000 | 24 |
| Bcl-2(D17C4) Rabbit mAb | 3498 | Cell Signaling Technology | 1:1000 | 26 |
| Caspase 3 Rabbit Polyclonal Antibody | 19677-1-AP | Abcam | 1:1000 | 35 |
| Caspase 9 Rabbit Polyclonal Antibody | 10380-1-AP | Abcam | 1:500 | 46,35 |
| CDK6 Polyclonal Antibody | 14052-1-AP | Proteintech | 1:1000 | 36 |
| Cyclin D1 Monoclonal Antibody | 60186-1-IG | Proteintech | 1:1000 | 34 |
| Ccnd 3 Antibody | ET1612-4 | Huabio | 1:1000 | 32 |
| MTF1 Rabbit pAb | A10824 | ABclonal | 1:500 | 122 |
| P-Histone H2A.X | sc-517348 | Santa Cruz Biotechnology | 1:1000 | 15 |
| Anti-beta Actin Antibody | ab8227 | Abcam | 1:4000 | 42 |
| Alpha Tubulin Monoclonal Antibody | 66031-1-Ig | Proteintech | 1:10,000 | 52 |
| Pierce goat anti-rabbit IGG | 31460 | Pierce | 1:10,000 | |
| HRP-Conjugated Affinipure Goat Anti-Mouse IgG (H + L) | SA00001-1 | Proteintech | 1:5000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Cai, Y.; Guo, J.; Dai, K.; Liu, L.; Chen, Z.; Wang, F.; Deng, M. Knockdown of KDM5B Leads to DNA Damage and Cell Cycle Arrest in Granulosa Cells via MTF1. Curr. Issues Mol. Biol. 2023, 45, 3219-3237. https://doi.org/10.3390/cimb45040210
Yang Y, Cai Y, Guo J, Dai K, Liu L, Chen Z, Wang F, Deng M. Knockdown of KDM5B Leads to DNA Damage and Cell Cycle Arrest in Granulosa Cells via MTF1. Current Issues in Molecular Biology. 2023; 45(4):3219-3237. https://doi.org/10.3390/cimb45040210
Chicago/Turabian StyleYang, Yingnan, Yu Cai, Jinjing Guo, Keke Dai, Liang Liu, Zili Chen, Feng Wang, and Mingtian Deng. 2023. "Knockdown of KDM5B Leads to DNA Damage and Cell Cycle Arrest in Granulosa Cells via MTF1" Current Issues in Molecular Biology 45, no. 4: 3219-3237. https://doi.org/10.3390/cimb45040210