

Identification of miR-671-5p and Its Related Pathways as General Mechanisms of Both Form-Deprivation and Lens-Induced Myopia in Mice
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
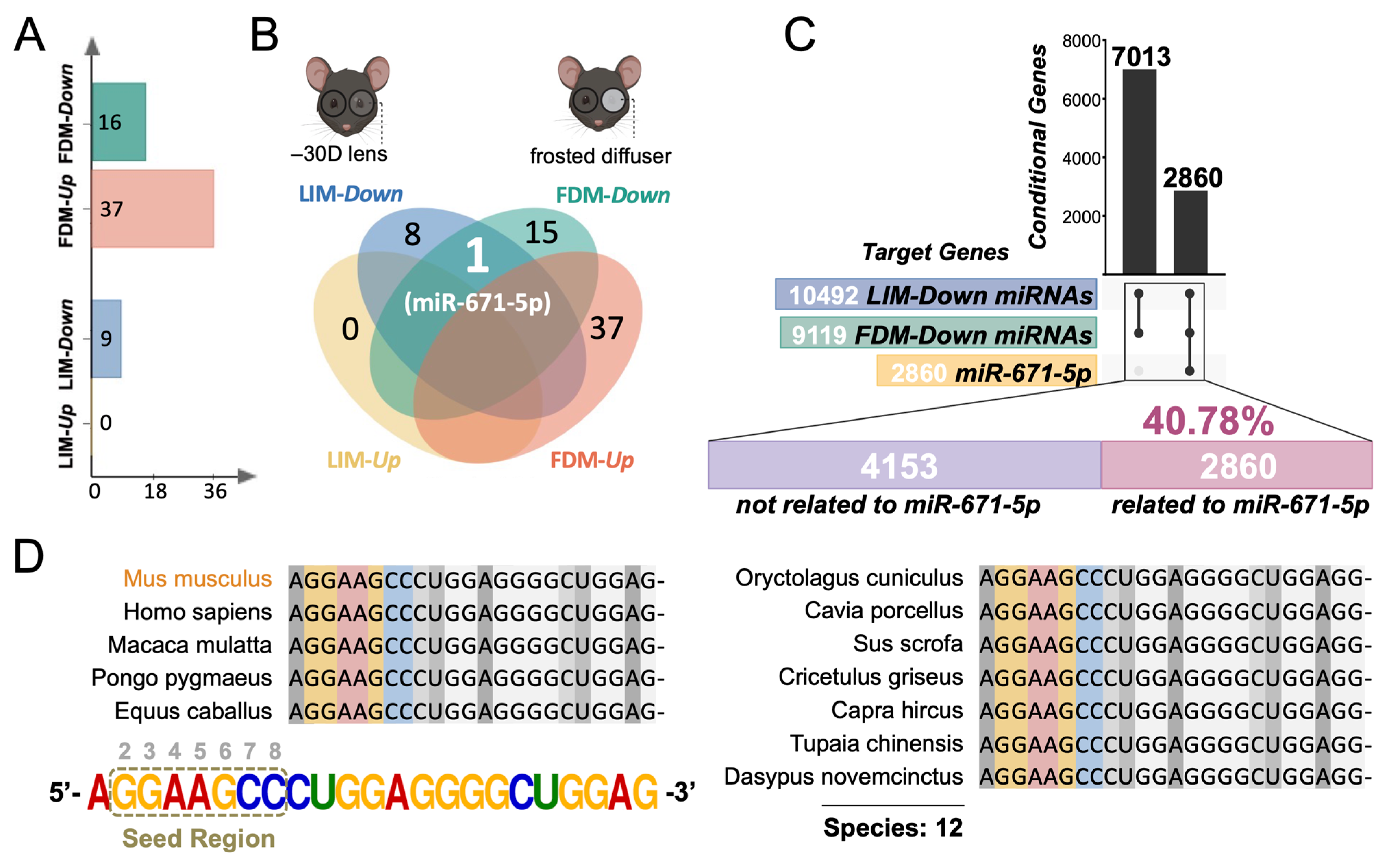
3.1. The Analytical Pipeline Reveals the General Regulatory Role of Retinal miR-671-5p in Both LIM and FDM Mouse Models
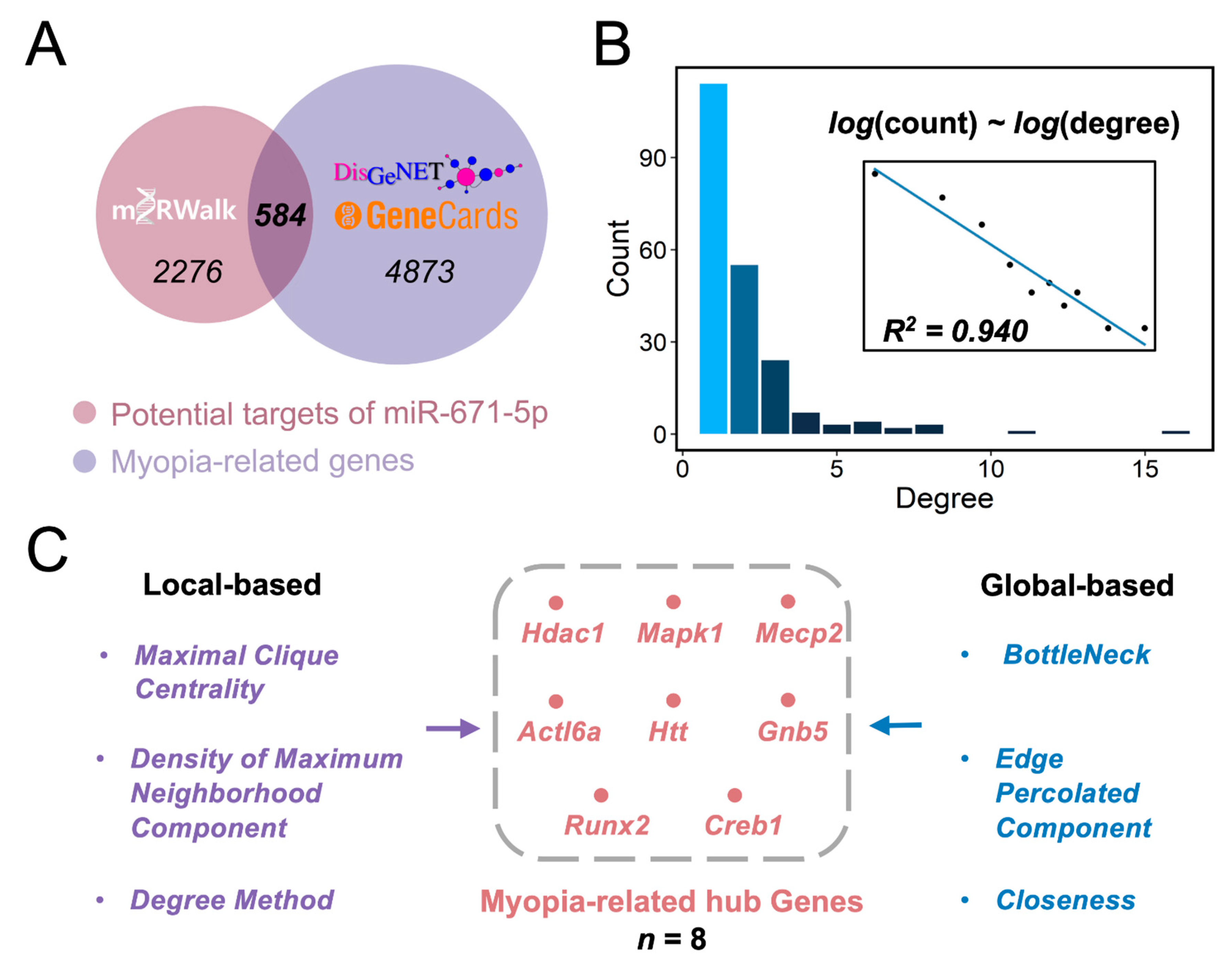
3.2. Identifying Hub Myopia-Related Genes Regulated by miR-671-5p
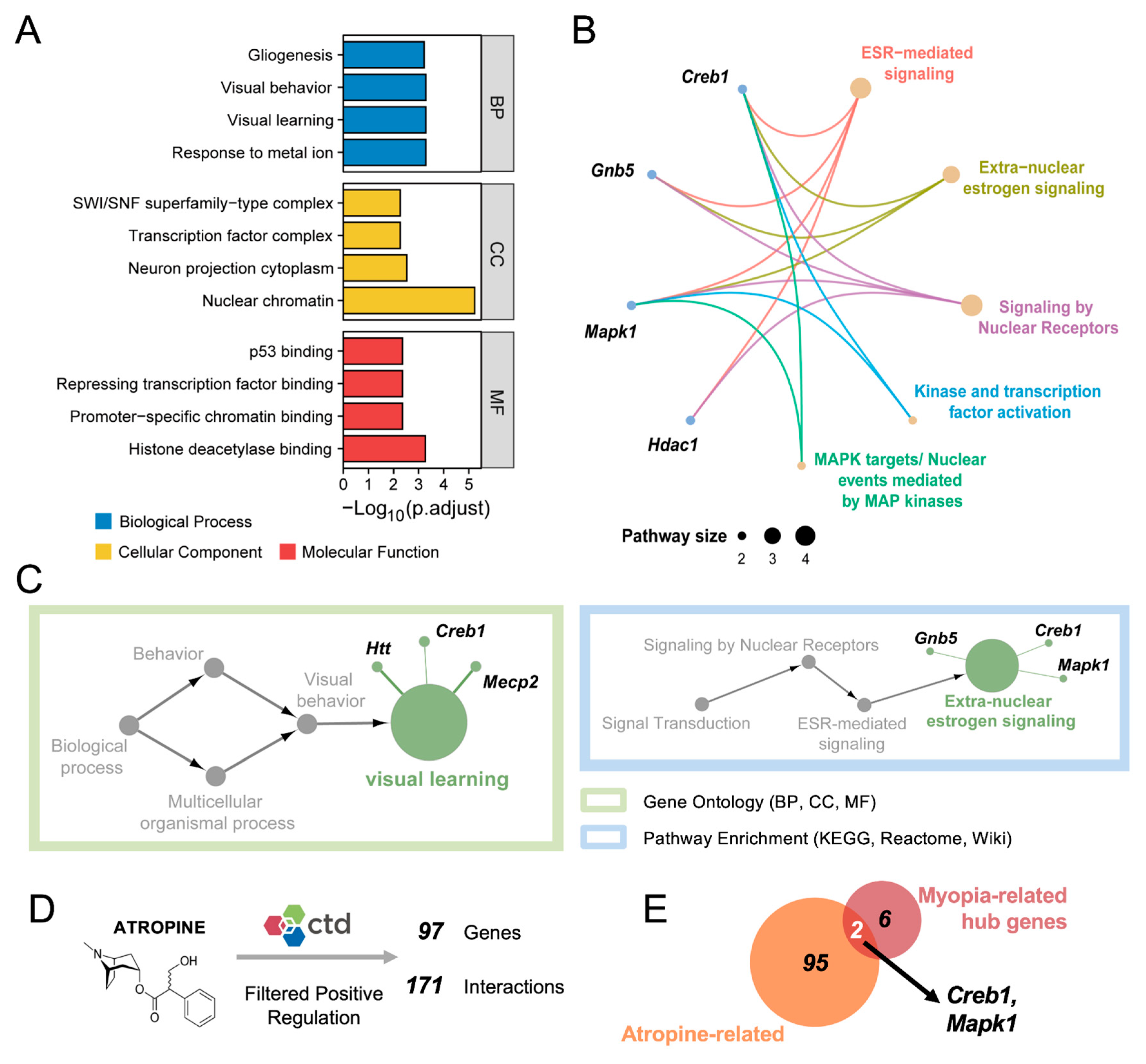
3.3. Integrated Analysis Reveals the Biological Functions of the Hub Genes
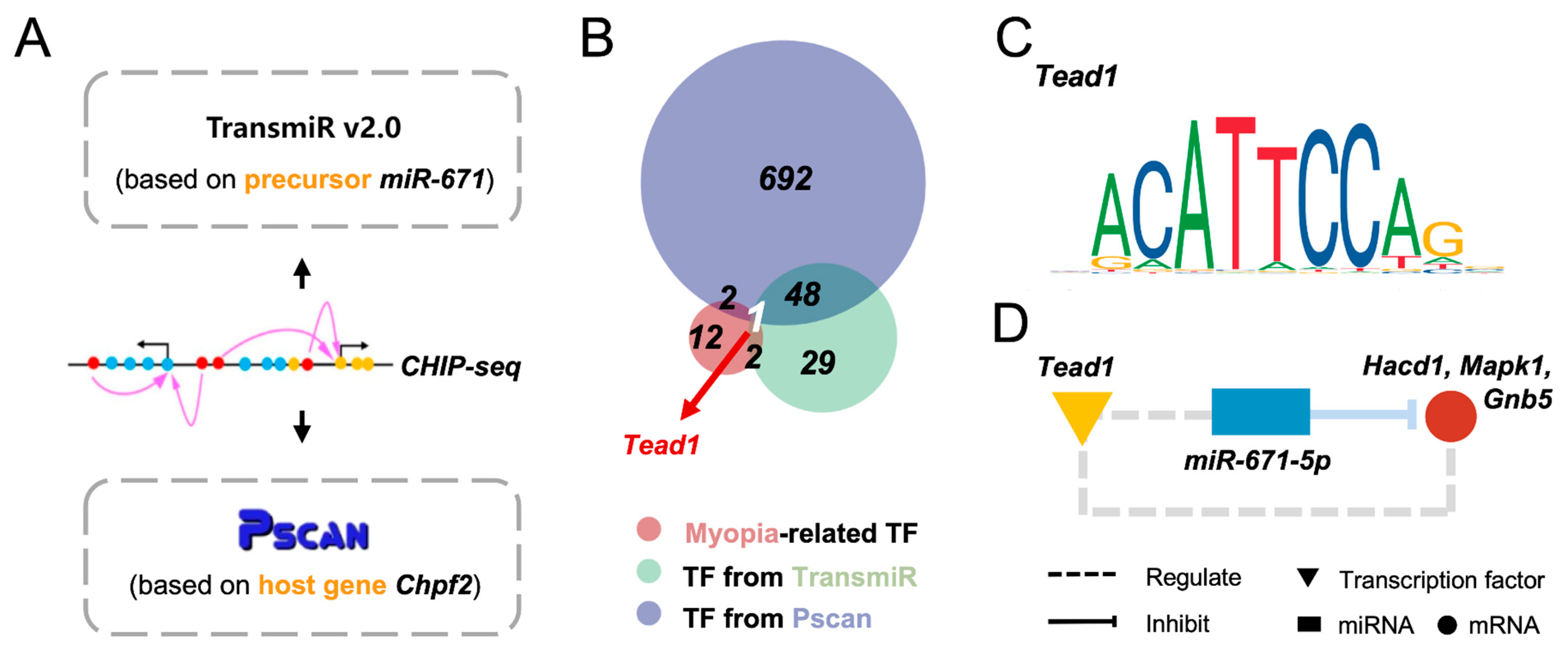
3.4. Tead1 May Act as an Upstream Regulator of miR-671-5p in Myopia Pathogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dolgin, E. The Myopia Boom. Nature 2015, 519, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Holden, B.A.; Fricke, T.R.; Wilson, D.A.; Jong, M.; Naidoo, K.S.; Sankaridurg, P.; Wong, T.Y.; Naduvilath, T.J.; Resnikoff, S. Global Prevalence of Myopia and High Myopia and Temporal Trends from 2000 through 2050. Ophthalmology 2016, 123, 1036–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naidoo, K.S.; Fricke, T.R.; Frick, K.D.; Jong, M.; Naduvilath, T.J.; Resnikoff, S.; Sankaridurg, P. Potential Lost Productivity Resulting from the Global Burden of Myopia: Systematic Review, Meta-Analysis, and Modeling. Ophthalmology 2019, 126, 338–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, I.G.; French, A.N.; Ashby, R.S.; Guo, X.; Ding, X.; He, M.; Rose, K.A. The Epidemics of Myopia: Aetiology and Prevention. Prog. Retin. Eye Res. 2018, 62, 134–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, H.; Fan, Z.-Y.; Tian, X.-D.; Xu, Y.-C. Comparison of Form-Deprived Myopia and Lens-Induced Myopia in Guinea Pigs. Int. J. Ophthalmol. 2014, 7, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Kurihara, T.; Hagiwara, Y.; Ikeda, S.-I.; Mori, K.; Jiang, X.; Torii, H.; Tsubota, K. Ocular-Component-Specific MiRNA Expression in a Murine Model of Lens-Induced Myopia. Int. J. Mol. Sci. 2019, 20, 3629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troilo, D.; Smith, E.L., III; Nickla, D.L.; Ashby, R.; Tkatchenko, A.V.; Ostrin, L.A.; Gawne, T.J.; Pardue, M.T.; Summers, J.A.; Kee, C.; et al. IMI—Report on Experimental Models of Emmetropization and Myopia. Investig. Ophthalmol. Vis. Sci. 2019, 60, M31–M88. [Google Scholar] [CrossRef] [Green Version]
- Morgan, I.G.; Ashby, R.S.; Nickla, D.L.; Guggenheim, J.A. Form Deprivation and Lens-Induced Myopia: Are They Different? Ophthalmic Physiol. Opt. 2013, 33, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Schaeffel, F.; Glasser, A.; Howland, H.C. Accommodation, Refractive Error and Eye Growth in Chickens. Vis. Res. 1988, 28, 639–657. [Google Scholar] [CrossRef]
- Raviola, E.; Wiesel, T.N. An Animal Model of Myopia. N. Engl. J. Med. 1985, 312, 1609–1615. [Google Scholar] [CrossRef]
- Smith, E.L.; Hung, L.-F.; Huang, J. Protective Effects of High Ambient Lighting on the Development of Form-Deprivation Myopia in Rhesus Monkeys. Investig. Ophthalmol. Vis. Sci. 2012, 53, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Ashby, R.S.; Schaeffel, F. The Effect of Bright Light on Lens Compensation in Chicks. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5247–5253. [Google Scholar] [CrossRef] [Green Version]
- Bartmann, M.; Schaeffel, F.; Hagel, G.; Zrenner, E. Constant Light Affects Retinal Dopamine Levels and Blocks Deprivation Myopia but Not Lens-Induced Refractive Errors in Chickens. Vis. Neurosci. 1994, 11, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Nickla, D.L.; Schroedl, F. Parasympathetic Influences on Emmetropization in Chicks: Evidence for Different Mechanisms in Form Deprivation vs. Negative Lens-Induced Myopia. Exp. Eye Res. 2012, 102, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, K.; Karouta, C.; Ashby, R. Form-Deprivation and Lens-Induced Myopia Are Similarly Affected by Pharmacological Manipulation of the Dopaminergic System in Chicks. Investig. Ophthalmol. Vis. Sci. 2020, 61, 4. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Yang, Z.; Feldkaemper, M.; Schaeffel, F. Changes in Dopamine and ZENK during Suppression of Myopia in Chicks by Intense Illuminance. Exp. Eye Res. 2016, 145, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Ashby, R. Animal Studies and the Mechanism of Myopia-Protection by Light? Optom. Vis. Sci. 2016, 93, 1052–1054. [Google Scholar] [CrossRef]
- French, A.N.; Ashby, R.S.; Morgan, I.G.; Rose, K.A. Time Outdoors and the Prevention of Myopia. Exp. Eye Res. 2013, 114, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Thomson, K.; Kelly, T.; Karouta, C.; Morgan, I.; Ashby, R. Insights into the Mechanism by Which Atropine Inhibits Myopia: Evidence against Cholinergic Hyperactivity and Modulation of Dopamine Release. Br. J. Pharmacol. 2021, 178, 4501–4517. [Google Scholar] [CrossRef]
- Qian, K.-W.; Li, Y.-Y.; Wu, X.-H.; Gong, X.; Liu, A.-L.; Chen, W.-H.; Yang, Z.; Cui, L.-J.; Liu, Y.-F.; Ma, Y.-Y.; et al. Altered Retinal Dopamine Levels in a Melatonin-Proficient Mouse Model of Form-Deprivation Myopia. Neurosci. Bull. 2022, 38, 992–1006. [Google Scholar] [CrossRef]
- Alkozi, H.A.; Franco, R.; Pintor, J.J. Epigenetics in the Eye: An Overview of the Most Relevant Ocular Diseases. Front. Genet. 2017, 8, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambros, V. The Functions of Animal MicroRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Kabekkodu, S.P.; Shukla, V.; Varghese, V.K.; D’ Souza, J.; Chakrabarty, S.; Satyamoorthy, K. Clustered MiRNAs and Their Role in Biological Functions and Diseases. Biol. Rev. Camb. Philos. Soc. 2018, 93, 1955–1986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, D.-N.; Zhu, Y.; Sun, H.; Gu, P.; Zhu, D.; Zhou, J. Regulation of Matrix Metalloproteinase-2 Secretion from Scleral Fibroblasts and Retinal Pigment Epithelial Cells by MiR-29a. BioMed Res. Int. 2017, 2017, 2647879. [Google Scholar] [CrossRef] [Green Version]
- Tkatchenko, A.V.; Luo, X.; Tkatchenko, T.V.; Vaz, C.; Tanavde, V.M.; Maurer-Stroh, S.; Zauscher, S.; Gonzalez, P.; Young, T.L. Large-Scale MicroRNA Expression Profiling Identifies Putative Retinal MiRNA-MRNA Signaling Pathways Underlying Form-Deprivation Myopia in Mice. PLoS ONE 2016, 11, e0162541. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.-C.; Hsi, E.; Hu, C.-Y.; Chou, W.-W.; Liang, C.-L.; Juo, S.-H.H. MicroRNA-328 May Influence Myopia Development by Mediating the PAX6 Gene. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2732–2739. [Google Scholar] [CrossRef] [Green Version]
- Mei, F.; Wang, J.; Chen, Z.; Yuan, Z. Potentially Important MicroRNAs in Form-Deprivation Myopia Revealed by Bioinformatics Analysis of MicroRNA Profiling. Ophthalmic Res. 2017, 57, 186–193. [Google Scholar] [CrossRef]
- Hong, N.; Jiang, B.; Gu, L.; Chen, S.-Y.; Tong, J.-P. MicroRNA-Expression Profiling in Myopia: A Meta-Analysis and Systematic Review. Ophthalmic Res. 2022, 65, 254–263. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. MiRWalk: An Online Resource for Prediction of MicroRNA Binding Sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]
- Li, D.; Knox, B.; Gong, B.; Chen, S.; Guo, L.; Liu, Z.; Tong, W.; Ning, B. Identification of Translational MicroRNA Biomarker Candidates for Ketoconazole-Induced Liver Injury Using Next-Generation Sequencing. Toxicol. Sci. 2021, 179, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; McMorran, R.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. The Comparative Toxicogenomics Database: Update 2019. Nucleic Acids Res. 2019, 47, D948–D954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A Comprehensive Platform Integrating Information on Human Disease-Associated Genes and Variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Assenov, Y.; Ramírez, F.; Schelhorn, S.-E.; Lengauer, T.; Albrecht, M. Computing Topological Parameters of Biological Networks. Bioinformatics 2008, 24, 282–284. [Google Scholar] [CrossRef] [Green Version]
- Alstott, J.; Bullmore, E.; Plenz, D. Powerlaw: A Python Package for Analysis of Heavy-Tailed Distributions. PLoS ONE 2014, 9, e85777. [Google Scholar] [CrossRef] [Green Version]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. CytoHubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape Plug-in to Decipher Functionally Grouped Gene Ontology and Pathway Annotation Networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.; Cui, Q.; Wang, J.; Zhou, Y. TransmiR v2.0: An Updated Transcription Factor-MicroRNA Regulation Database. Nucleic Acids Res. 2019, 47, D253–D258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbagallo, D.; Condorelli, A.; Ragusa, M.; Salito, L.; Sammito, M.; Banelli, B.; Caltabiano, R.; Barbagallo, G.; Zappalà, A.; Battaglia, R.; et al. Dysregulated MiR-671-5p/CDR1-AS / CDR1 / VSNL1 Axis Is Involved in Glioblastoma Multiforme. Oncotarget 2016, 7, 4746–4759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambelli, F.; Pesole, G.; Pavesi, G. Pscan: Finding over-Represented Transcription Factor Binding Site Motifs in Sequences from Co-Regulated or Co-Expressed Genes. Nucleic Acids Res. 2009, 37, W247–W252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.; Wan, C.; Mei, S.; Qin, Q.; Wu, Q.; Sun, H.; Chen, C.-H.; Brown, M.; Zhang, X.; Meyer, C.A.; et al. Cistrome Data Browser: Expanded Datasets and New Tools for Gene Regulatory Analysis. Nucleic Acids Res. 2019, 47, D729–D735. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Li, S.M.; An, W.; Du, J.; Liang, X.; Sun, Y.; Zhang, D.; Tian, J.; Wang, N. Safety and Efficacy of Low-Dose At-ropine Eyedrops for the Treatment of Myopia Progression in Chinese Children: A Randomized Clinical Trial. JAMA Ophthalmol. 2020, 11, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- McBrien, N.A.; Stell, W.K.; Carr, B. How Does Atropine Exert Its Anti-Myopia Effects? Ophthalmic Physiol. Opt. 2013, 33, 373–378. [Google Scholar] [CrossRef]
- Wang, L.Z.; Syn, N.; Li, S.; Barathi, V.A.; Tong, L.; Neo, J.; Beuerman, R.W.; Zhou, L. The Penetration and Distribution of Topical Atropine in Animal Ocular Tissues. Acta Ophthalmol. 2019, 97, e238–e247. [Google Scholar] [CrossRef]
- Yang, L.; Yue, W.; Zhang, H.; Zhang, Z.; Xue, R.; Dong, C.; Liu, F.; Chang, N.; Yang, L.; Li, L. Dual Targeting of Angipoietin-1 and von Willebrand Factor by MicroRNA-671-5p Attenuates Liver Angiogenesis and Fibrosis. Hepatol. Commun. 2022, 6, 1425–1442. [Google Scholar] [CrossRef]
- Dou, C.; Zhou, Z.; Xu, Q.; Liu, Z.; Zeng, Y.; Wang, Y.; Li, Q.; Wang, L.; Yang, W.; Liu, Q.; et al. Hypoxia-Induced TUFT1 Promotes the Growth and Metastasis of Hepatocellular Carcinoma by Activating the Ca2+/PI3K/AKT Pathway. Oncogene 2019, 38, 1239–1255. [Google Scholar] [CrossRef]
- Altuvia, Y.; Landgraf, P.; Lithwick, G.; Elefant, N.; Pfeffer, S.; Aravin, A.; Brownstein, M.J.; Tuschl, T.; Margalit, H. Clustering and Conservation Patterns of Human MicroRNAs. Nucleic Acids Res. 2005, 33, 2697–2706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Yu, X.; Wen, Y.; Jin, L.; Zhang, L.; Zhu, H.; Zhang, D.; Xie, C.; Guo, D.; Tong, J.; et al. Functions of Retinal Astrocytes and Müller Cells in Mammalian Myopia. BMC Ophthalmol. 2022, 22, 451. [Google Scholar] [CrossRef] [PubMed]
- Paylakhi, S.; Labelle-Dumais, C.; Tolman, N.G.; Sellarole, M.A.; Seymens, Y.; Saunders, J.; Lakosha, H.; deVries, W.N.; Orr, A.C.; Topilko, P.; et al. Müller Glia-Derived PRSS56 Is Required to Sustain Ocular Axial Growth and Prevent Refractive Error. PLoS Genet. 2018, 14, e1007244. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, W.; Zhao, F.; Zhou, Q.; Reinach, P.S.; Deng, L.; Ma, L.; Luo, S.; Srinivasalu, N.; Pan, M.; et al. Scleral Hypoxia Is a Target for Myopia Control. Proc. Natl. Acad. Sci. USA 2018, 115, E7091–E7100. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.-F.; Xie, H.-L.; Mao, X.-J.; Zhu, X.-B.; Xie, Z.-K.; Yang, H.-H.; Gao, Y.; Jin, X.-F.; Pan, Y.; Zhou, F. Relevant Factors of Estrogen Changes of Myopia in Adolescent Females. Chin. Med. J. 2015, 128, 659–663. [Google Scholar] [CrossRef]
- Lyu, I.J.; Kim, M.H.; Baek, S.-Y.; Kim, J.; Park, K.-A.; Oh, S.Y. The Association Between Menarche and Myopia: Findings From the Korean National Health and Nutrition Examination, 2008–2012. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4712–4718. [Google Scholar] [CrossRef] [Green Version]
- Vajaranant, T.S.; Pasquale, L.R. Estrogen Deficiency Accelerates Aging of the Optic Nerve. J. N. Am. Menopause Soc. 2012, 19, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Eisner, A.; Burke, S.N.; Toomey, M.D. Visual Sensitivity across the Menstrual Cycle. Vis. Neurosci. 2004, 21, 513–531. [Google Scholar] [CrossRef]
- Ogueta, S.B.; Schwartz, S.D.; Yamashita, C.K.; Farber, D.B. Estrogen Receptor in the Human Eye: Influence of Gender and Age on Gene Expression. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1906–1911. [Google Scholar]
- Bunone, G.; Briand, P.A.; Miksicek, R.J.; Picard, D. Activation of the Unliganded Estrogen Receptor by EGF Involves the MAP Kinase Pathway and Direct Phosphorylation. EMBO J. 1996, 15, 2174–2183. [Google Scholar] [CrossRef]
- Berto, M.; Jean, V.; Zwart, W.; Picard, D. ERα Activity Depends on Interaction and Target Site Corecruitment with Phosphorylated CREB1. Life Sci. Alliance 2018, 1, e201800055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazennec, G.; Thomas, J.A.; Katzenellenbogen, B.S. Involvement of Cyclic AMP Response Element Binding Protein (CREB) and Estrogen Receptor Phosphorylation in the Synergistic Activation of the Estrogen Receptor by Estradiol and Protein Kinase Activators. J. Steroid Biochem. Mol. Biol. 2001, 77, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, X.; Jiang, Y.; Liu, R.; Cao, D.; Pan, J.; Luo, Y. Identification of Key MiRNAs and Genes for Mouse Retinal Development Using a Linear Model. Mol. Med. Rep. 2020, 22, 494–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Yang, X.; Luo, Z.; Wu, J.; Lin, H. HIF-1α Aggravates Pathologic Myopia through the MiR-150-5p/LAMA4/P38 MAPK Signaling Axis. Mol. Cell. Biochem. 2022, 477, 1065–1074. [Google Scholar] [CrossRef]
- Huang, J.; Chen, P.; Yu, J.; Xu, L.; Yang, Y.; Wu, N.; Tan, M.; Ge, J.; Yu, K. Distribution of Phosphorylated Cyclic AMP Response Element Binding Protein (p-CREB-1) in Rat Retina. Int. J. Clin. Exp. Pathol. 2017, 10, 1253–1262. [Google Scholar]
- Shaywitz, A.J.; Greenberg, M.E. CREB: A Stimulus-Induced Transcription Factor Activated by a Diverse Array of Extracellular Signals. Annu. Rev. Biochem. 1999, 68, 821–861. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, X.; Zhuang, J.; Yu, K. The Role of Retinal Dysfunction in Myopia Development. Cell. Mol. Neurobiol. 2022. [Google Scholar] [CrossRef]
- Enzo, E.; Santinon, G.; Pocaterra, A.; Aragona, M.; Bresolin, S.; Forcato, M.; Grifoni, D.; Pession, A.; Zanconato, F.; Guzzo, G.; et al. Aerobic Glycolysis Tunes YAP/TAZ Transcriptional Activity. EMBO J. 2015, 34, 1349–1370. [Google Scholar] [CrossRef]
- Liu-Chittenden, Y.; Huang, B.; Shim, J.S.; Chen, Q.; Lee, S.-J.; Anders, R.A.; Liu, J.O.; Pan, D. Genetic and Pharmacological Disruption of the TEAD-YAP Complex Suppresses the Oncogenic Activity of YAP. Genes Dev. 2012, 26, 1300–1305. [Google Scholar] [CrossRef] [Green Version]
- Halder, G.; Dupont, S.; Piccolo, S. Transduction of Mechanical and Cytoskeletal Cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 2012, 13, 591–600. [Google Scholar] [CrossRef]
- Yu, F.-X.; Guan, K.-L. The Hippo Pathway: Regulators and Regulations. Genes Dev. 2013, 27, 355–371. [Google Scholar] [CrossRef] [Green Version]
- Sawada, A.; Kiyonari, H.; Ukita, K.; Nishioka, N.; Imuta, Y.; Sasaki, H. Redundant Roles of Tead1 and Tead2 in Notochord Development and the Regulation of Cell Proliferation and Survival. Mol. Cell. Biol. 2008, 28, 3177–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrauwen, I.; Szelinger, S.; Siniard, A.L.; Corneveaux, J.J.; Kurdoglu, A.; Richholt, R.; De Both, M.; Malenica, I.; Swaminathan, S.; Rangasamy, S.; et al. A De Novo Mutation in TEAD1 Causes Non-X-Linked Aicardi Syndrome. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3896–3904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fossdal, R.; Jonasson, F.; Kristjansdottir, G.T.; Kong, A.; Stefansson, H.; Gosh, S.; Gulcher, J.R.; Stefansson, K. A Novel TEAD1 Mutation Is the Causative Allele in Sveinsson’s Chorioretinal Atrophy (Helicoid Peripapillary Chorioretinal Degeneration). Hum. Mol. Genet. 2004, 13, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Flitcroft, D.I. The Complex Interactions of Retinal, Optical and Environmental Factors in Myopia Aetiology. Prog. Retin. Eye Res. 2012, 31, 622–660. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Z.; Huang, Y.; Chen, X.; Chen, T.; Hou, X.; Yu, N.; Li, Y.; Qiu, J.; Chen, P.; Yu, K.; et al. Identification of miR-671-5p and Its Related Pathways as General Mechanisms of Both Form-Deprivation and Lens-Induced Myopia in Mice. Curr. Issues Mol. Biol. 2023, 45, 2060-2072. https://doi.org/10.3390/cimb45030132
Cui Z, Huang Y, Chen X, Chen T, Hou X, Yu N, Li Y, Qiu J, Chen P, Yu K, et al. Identification of miR-671-5p and Its Related Pathways as General Mechanisms of Both Form-Deprivation and Lens-Induced Myopia in Mice. Current Issues in Molecular Biology. 2023; 45(3):2060-2072. https://doi.org/10.3390/cimb45030132
Chicago/Turabian StyleCui, Zedu, Yuke Huang, Xi Chen, Taiwei Chen, Xiangtao Hou, Na Yu, Yan Li, Jin Qiu, Pei Chen, Keming Yu, and et al. 2023. "Identification of miR-671-5p and Its Related Pathways as General Mechanisms of Both Form-Deprivation and Lens-Induced Myopia in Mice" Current Issues in Molecular Biology 45, no. 3: 2060-2072. https://doi.org/10.3390/cimb45030132