
Characterization of Enlarged Tongues in Cloned Piglets
 , , ,
, , ,
Abstract
:
1. Introduction
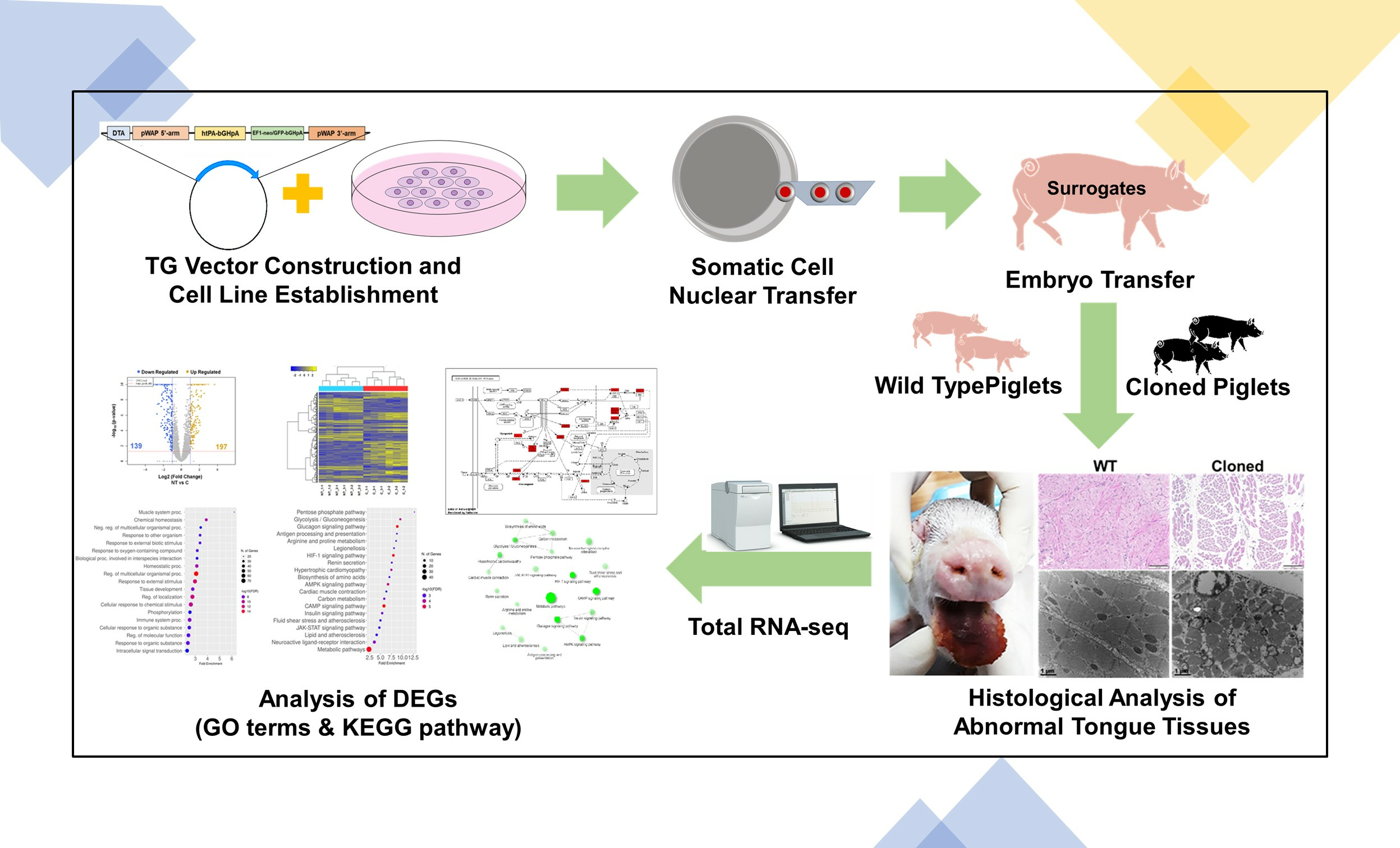
2. Materials and Methods
2.1. General Information and Ethycal Statement
2.2. Experimental Design
2.3. In Vitro Maturation
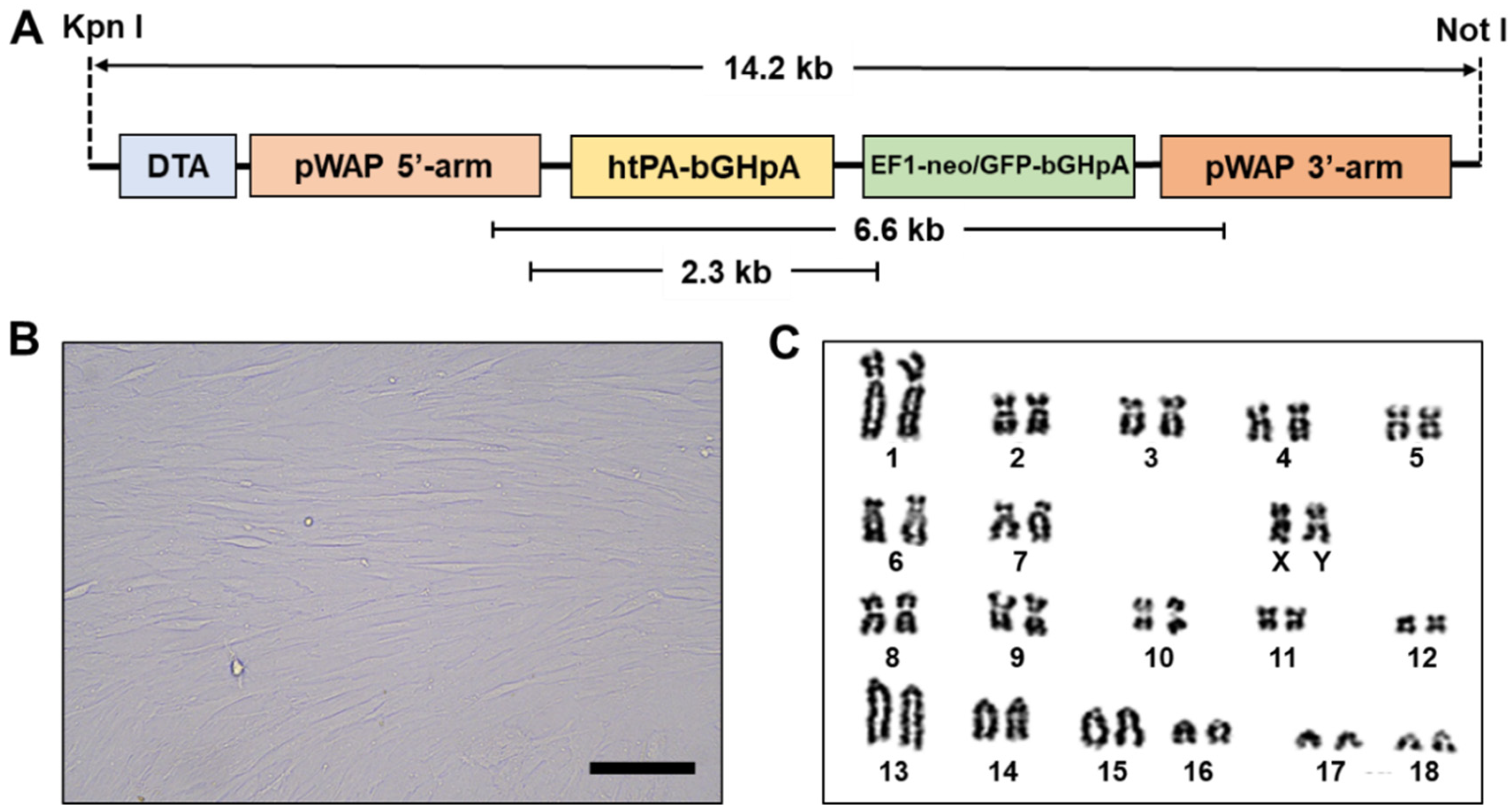
2.4. Production of Transgenic Cell Lines
2.5. Somatic Cell Nuclear Transfer
2.6. In Vitro Culture and Embryo Transfer
2.7. Histology of Tongue Tissues
2.8. Genomic Analysis
2.9. Statistical Analysis
3. Results
3.1. Establishment of Transgenic Cell Lines
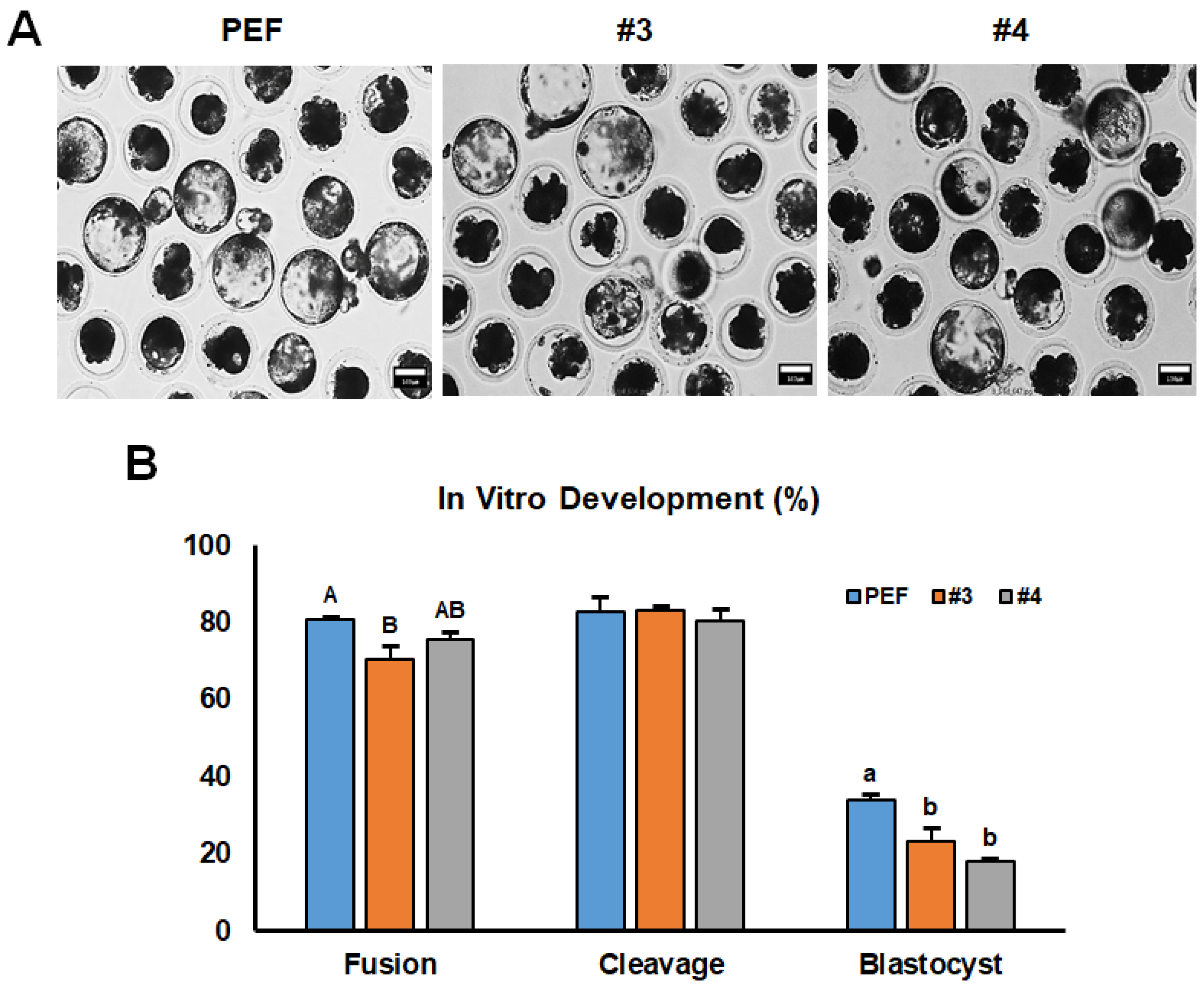
3.2. Developmental Competence of Cloned Embryos
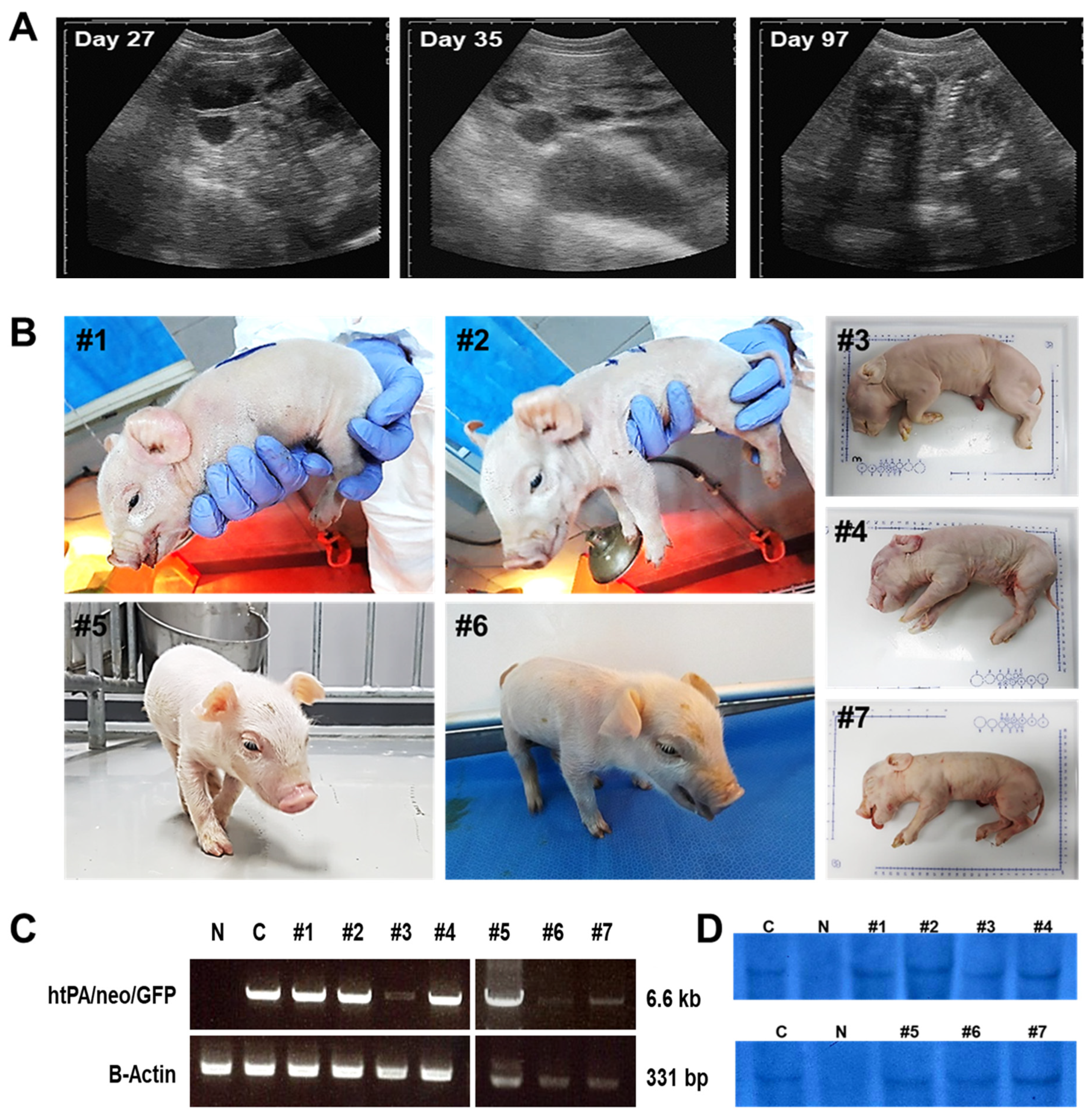
3.3. Production of Cloned htPA Transgenic Piglets
3.4. Histological Analysis of Abnormal Tongues in Cloned htPA Piglets
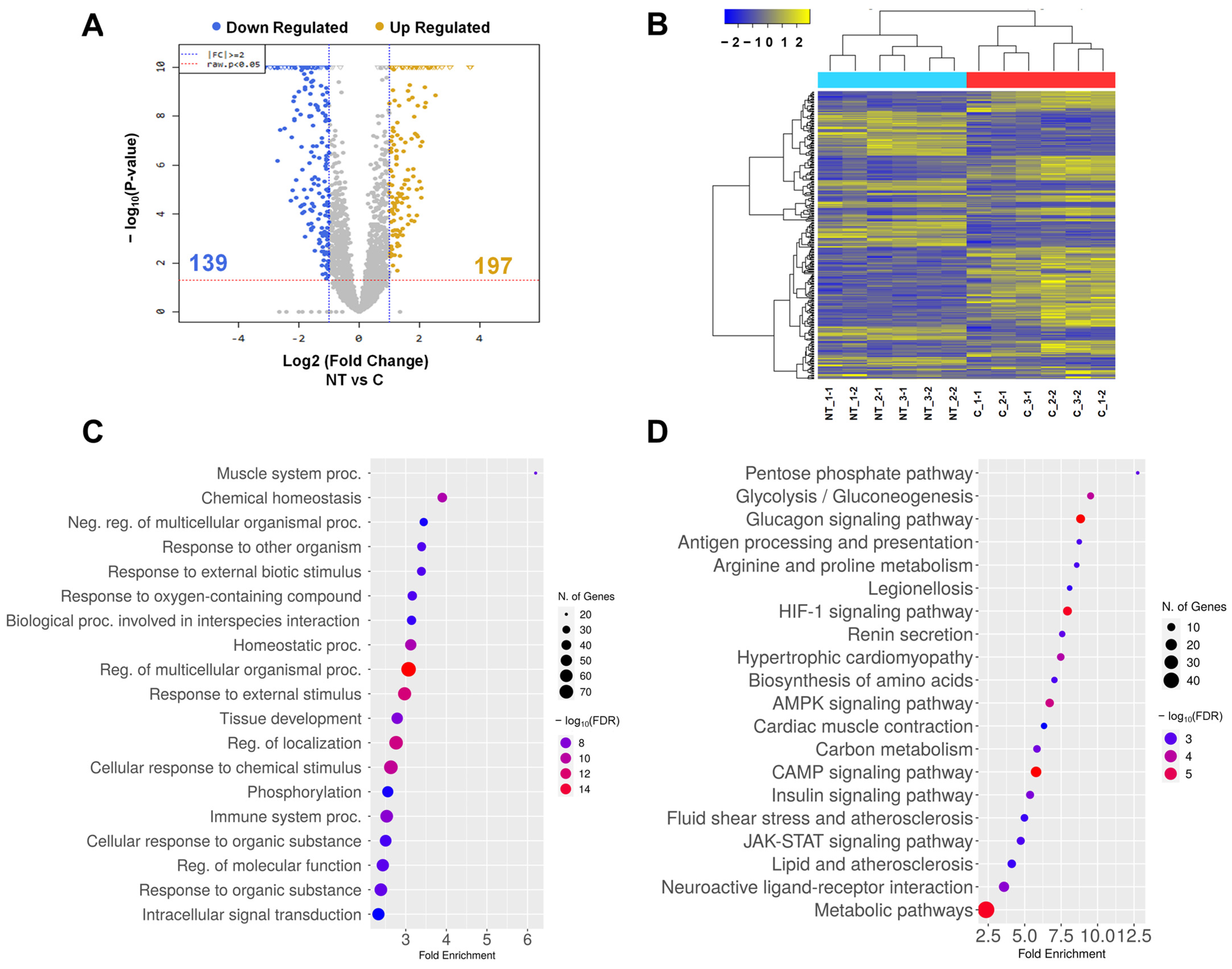
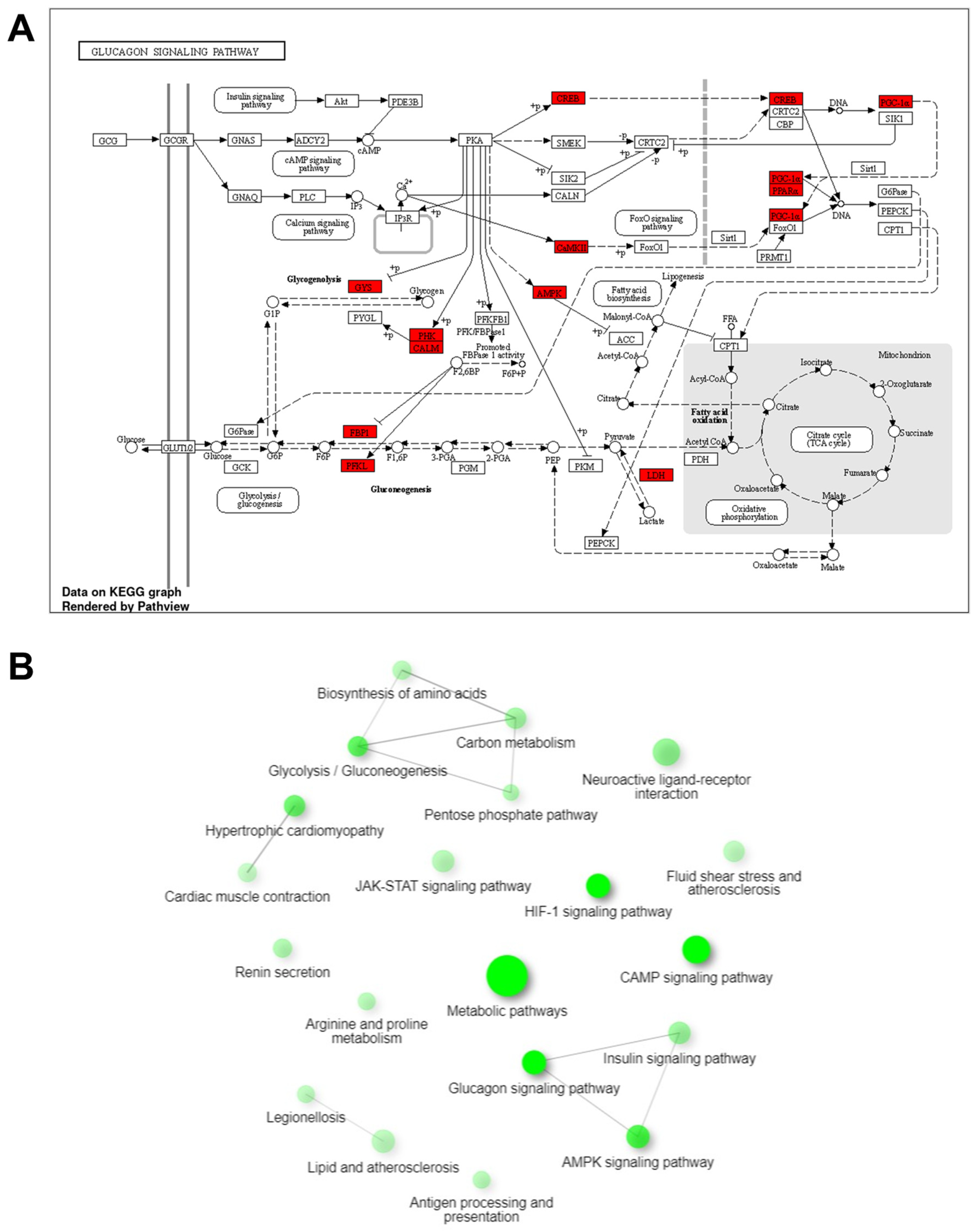
3.5. Analysis of Transcriptome of Abnormal Tongue in Cloned Piglets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schelstraete, W.; Devreese, M.; Croubels, S. Comparative toxicokinetics of Fusarium mycotoxins in pigs and humans. Food Chem. Toxicol. 2020, 137, 111140. [Google Scholar] [CrossRef] [PubMed]
- Camacho, P.; Fan, H.; Liu, Z.; He, J.Q. Large Mammalian Animal Models of Heart Disease. J. Cardiovasc. Dev. Dis. 2016, 3, 30. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Zheng, W.; Chen, M.; Zou, Q.; Tang, C.; Zhou, X. Genetically engineered pigs for xenotransplantation: Hopes and challenges. Front. Cell Dev. Biol. 2022, 10, 1093534. [Google Scholar] [CrossRef] [PubMed]
- Blutke, A.; Wanke, R. Sampling Strategies and Processing of Biobank Tissue Samples from Porcine Biomedical Models. J. Vis. Exp. 2018, 133, 57276. [Google Scholar] [CrossRef]
- Gordon, J.W.; Scangos, G.A.; Plotkin, D.J.; Barbosa, J.A.; Ruddle, F.H. Genetic transformation of mouse embryos by microinjection of purified DNA. Proc. Natl. Acad. Sci. USA 1980, 77, 7380–7384. [Google Scholar] [CrossRef]
- Campbell, K.H. A background to nuclear transfer and its applications in agriculture and human therapeutic medicine. J. Anat. 2002, 200, 267–275. [Google Scholar] [CrossRef]
- Hryhorowicz, M.; Lipinski, D.; Hryhorowicz, S.; Nowak-Terpilowska, A.; Ryczek, N.; Zeyland, J. Application of Genetically Engineered Pigs in Biomedical Research. Genes 2020, 11, 670. [Google Scholar] [CrossRef]
- Young, L.E.; Fairburn, H.R. Improving the safety of embryo technologies: Possible role of genomic imprinting. Theriogenology 2000, 53, 627–648. [Google Scholar] [CrossRef]
- Matoba, S.; Liu, Y.; Lu, F.; Iwabuchi, K.A.; Shen, L.; Inoue, A.; Zhang, Y. Embryonic development following somatic cell nuclear transfer impeded by persisting histone methylation. Cell 2014, 159, 884–895. [Google Scholar] [CrossRef]
- Matoba, S.; Zhang, Y. Somatic Cell Nuclear Transfer Reprogramming: Mechanisms and Applications. Cell Stem Cell 2018, 23, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhai, Y.; Ma, X.; Zhang, S.; An, X.; Yu, H.; Li, Z. Down-Regulation of H3K4me3 by MM-102 Facilitates Epigenetic Reprogramming of Porcine Somatic Cell Nuclear Transfer Embryos. Cell Physiol. Biochem. 2018, 45, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.J.; Cheng, W.T.; Wu, S.C.; Chen, H.L.; Tsai, T.C.; Yang, S.H.; Chen, C.M. Differential differences in methylation status of putative imprinted genes among cloned swine genomes. PLoS ONE 2012, 7, e32812. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jia, Q.; Zhao, J.; Li, X.; Yu, M.; Samuel, M.S.; Zhao, S.; Prather, R.S.; Li, C. Dysregulation of genome-wide gene expression and DNA methylation in abnormal cloned piglets. BMC Genom. 2014, 15, 811. [Google Scholar] [CrossRef] [PubMed]
- Park, K.W.; Lai, L.; Cheong, H.T.; Cabot, R.; Sun, Q.Y.; Wu, G.; Rucker, E.B.; Durtschi, D.; Bonk, A.; Samuel, M.; et al. Mosaic gene expression in nuclear transfer-derived embryos and the production of cloned transgenic pigs from ear-derived fibroblasts. Biol. Reprod. 2002, 66, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Qiao, C.M.; Liu, W.W.; Jiang, H.Y.; Jing, Q.Q.; Liao, Y.Y.; Xing, Y.Y. Genome-wide DNA methylation and transcription analysis in tongue and biceps femoris muscles of cloned pigs with macroglossia. Anim. Genet. 2021, 52, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Winther, K.D.; Secher, J.O.; Callesen, H. Postmortem findings in cloned and transgenic piglets dead before weaning. Theriogenology 2015, 84, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Riego, E.; Limonta, J.; Aguilar, A.; Pérez, A.; de Armas, R.; Solano, R.; Ramos, B.; Castro, F.O.; de la Fuente, J. Production of transgenic mice and rabbits that carry and express the human tissue plasminogen activator cDNA under the control of a bovine alpha S1 casein promoter. Theriogenology 1993, 39, 1173–1185. [Google Scholar] [CrossRef]
- Song, S.; Ge, X.; Cheng, Y.; Lu, R.; Zhang, T.; Yu, B.; Ji, X.; Qi, Z.; Rong, Y.; Yuan, Y.; et al. High-level expression of a novel recombinant human plasminogen activator (rhPA) in the milk of transgenic rabbits and its thrombolytic bioactivity in vitro. Mol. Biol. Rep. 2016, 43, 775–783. [Google Scholar] [CrossRef]
- Denman, J.; Hayes, M.; O’Day, C.; Edmunds, T.; Bartlett, C.; Hirani, S.; Ebert, K.M.; Gordon, K.; McPherson, J.M. Transgenic expression of a variant of human tissue-type plasminogen activator in goat milk: Purification and characterization of the recombinant enzyme. Biotechnology 1991, 9, 839–843. [Google Scholar] [CrossRef]
- Chung, H.J.; Park, H.J.; Baek, S.Y.; Park, J.K.; Lee, W.Y.; Kim, K.W.; Jo, Y.M.; Hochi, S.; Kim, Y.M.; Choi, T.J.; et al. Production of human tissue-type plasminogen activator (htPA) using in vitro cultured transgenic pig mammary gland cells. Anim. Biotechnol. 2019, 30, 317–322. [Google Scholar] [CrossRef]
- Gravanis, I.; Tsirka, S.E. Tissue-type plasminogen activator as a therapeutic target in stroke. Expert. Opin. Ther. Targets 2008, 12, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.S.; Shim, J.; Oh, K.B.; Lee, H.; Park, M.R. cd26 Knockdown Negatively Affects Porcine Parthenogenetic Preimplantation Embryo Development. Animals 2022, 12, 1662. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Lee, H.; Oh, K.B. DNA methylation change of pWAP promoter region in porcine transgenic somatic cells. Korea Acad.-Ind. Coop. Soc. 2021, 22, 317–323. [Google Scholar] [CrossRef]
- Hwang, I.S.; Kwon, D.J.; Oh, K.B.; Ock, S.A.; Chung, H.J.; Cho, I.C.; Lee, J.W.; Im, G.S.; Hwang, S. Production of Cloned Korean Native Pig by Somatic Cell Nuclear Transfer. Dev. Reprod. 2015, 19, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.J.; Kim, D.H.; Hwang, I.S.; Kim, D.E.; Kim, H.J.; Kim, J.S.; Lee, K.; Im, G.S.; Lee, J.W.; Hwang, S. Generation of alpha-1,3-galactosyltransferase knocked-out transgenic cloned pigs with knocked-in five human genes. Transgenic Res. 2017, 26, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, K.; Suzuki, C.; Tanaka, A.; Anas, I.M.; Iwamura, S. Birth of piglets derived from porcine zygotes cultured in a chemically defined medium. Biol. Reprod. 2002, 66, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.M.; Katayama, M.; Magnuson, S.R.; Falduto, M.T.; Torres, K.E. Transcript profiling of individual twin blastomeres derived by splitting two-cell stage murine embryos. Biol. Reprod. 2011, 84, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Kruithof, E.K.; Dunoyer-Geindre, S. Human tissue-type plasminogen activator. Thromb. Haemost. 2014, 112, 243–254. [Google Scholar] [CrossRef]
- Lijnen, H.R.; Collen, D. Tissue-type plasminogen activator. Ann. Biol. Clin. 1987, 45, 198–201. [Google Scholar]
- Loscalzo, J.; Braunwald, E. Tissue plasminogen activator. N. Engl. J. Med. 1988, 319, 925–931. [Google Scholar] [CrossRef]
- Gordon, K.; Lee, E.; Vitale, J.A.; Smith, A.E.; Westphal, H.; Hennighausen, L. Production of human tissue plasminogen activator in transgenic mouse milk. Nat. Biotechnol. 1987, 5, 1183–1187. [Google Scholar] [CrossRef]
- Ito, Y.; Noguchi, K.; Morishima, Y.; Yamaguchi, K. Generation and characterization of tissue-type plasminogen activator transgenic rats. J. Thromb. Thrombolysis 2018, 45, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Ebert, K.M.; Selgrath, J.P.; DiTullio, P.; Denman, J.; Smith, T.E.; Memon, M.A.; Schindler, J.E.; Monastersky, G.M.; Vitale, J.A.; Gordon, K. Transgenic production of a variant of human tissue-type plasminogen activator in goat milk: Generation of transgenic goats and analysis of expression. Biotechnology 1991, 9, 835–838. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Li, Z.L.; Huang, W.M.; Huang, W.J.; Li, J. Expression of human tissue-type plasminogen activator in cow mammary gland. Di Yi Jun Yi Da Xue Xue Bao 2004, 24, 546–548, 552. [Google Scholar] [PubMed]
- Nukumi, N.; Ikeda, K.; Osawa, M.; Iwamori, T.; Naito, K.; Tojo, H. Regulatory function of whey acidic protein in the proliferation of mouse mammary epithelial cells in vivo and in vitro. Dev. Biol. 2004, 274, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Triplett, A.A.; Sakamoto, K.; Matulka, L.A.; Shen, L.; Smith, G.H.; Wagner, K.U. Expression of the whey acidic protein (Wap) is necessary for adequate nourishment of the offspring but not functional differentiation of mammary epithelial cells. Genesis 2005, 43, 1–11. [Google Scholar] [CrossRef]
- Park, J.K.; Lee, Y.K.; Lee, P.; Chung, H.J.; Kim, S.; Lee, H.G.; Seo, M.K.; Han, J.H.; Park, C.G.; Kim, H.T.; et al. Recombinant human erythropoietin produced in milk of transgenic pigs. J. Biotechnol. 2006, 122, 362–371. [Google Scholar] [CrossRef]
- Lee, M.H.; Lin, Y.S.; Tu, C.F.; Yen, C.H. Recombinant human factor IX produced from transgenic porcine milk. Biomed. Res. Int. 2014, 2014, 315375. [Google Scholar] [CrossRef]
- Velander, W.H.; Johnson, J.L.; Page, R.L.; Russell, C.G.; Subramanian, A.; Wilkins, T.D.; Gwazdauskas, F.C.; Pittius, C.; Drohan, W.N. High-level expression of a heterologous protein in the milk of transgenic swine using the cDNA encoding human protein C. Proc. Natl. Acad. Sci. USA 1992, 89, 12003–12007. [Google Scholar] [CrossRef]
- Kim, G.; Roy, P.K.; Fang, X.; Hassan, B.M.; Cho, J. Improved preimplantation development of porcine somatic cell nuclear transfer embryos by caffeine treatment. J. Vet. Sci. 2019, 20, e31. [Google Scholar] [CrossRef] [PubMed]
- Boquest, A.C.; Grupen, C.G.; Harrison, S.J.; McIlfatrick, S.M.; Ashman, R.J.; d’Apice, A.J.; Nottle, M.B. Production of cloned pigs from cultured fetal fibroblast cells. Biol. Reprod. 2002, 66, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.K.; Somfai, T.; Salamone, D.; Thu Huong, V.T.; Le Thi Nguyen, H.; Huu, Q.X.; Hoang, A.T.; Phan, H.T.; Thi Pham, Y.K.; Pham, L.D. Optimization of donor cell cycle synchrony, maturation media and embryo culture system for somatic cell nuclear transfer in the critically endangered Vietnamese Ỉ pig. Theriogenology 2021, 166, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Kurome, M.; Geistlinger, L.; Kessler, B.; Zakhartchenko, V.; Klymiuk, N.; Wuensch, A.; Richter, A.; Baehr, A.; Kraehe, K.; Burkhardt, K.; et al. Factors influencing the efficiency of generating genetically engineered pigs by nuclear transfer: Multi-factorial analysis of a large data set. BMC Biotechnol. 2013, 13, 43. [Google Scholar] [CrossRef] [PubMed]
- Meijer, A.E. The pentose phosphate pathway in skeletal muscle under patho-physiological conditions. A combined histochemical and biochemical study. Prog. Histochem. Cytochem. 1991, 22, 1–118. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.; Yang, J.; Zhou, S.; Wang, Y.; Li, Y.; Tong, X. The Role of the Pentose Phosphate Pathway in Diabetes and Cancer. Front. Endocrinol. 2020, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Ozen, H. Glycogen storage diseases: New perspectives. World J. Gastroenterol. 2007, 13, 2541–2553. [Google Scholar] [CrossRef]
- Charron, M.J.; Vuguin, P.M. Lack of glucagon receptor signaling and its implications beyond glucose homeostasis. J. Endocrinol. 2015, 224, R123–R130. [Google Scholar] [CrossRef]
- DeBaun, M.R.; King, A.A.; White, N. Hypoglycemia in Beckwith-Wiedemann syndrome. Semin. Perinatol. 2000, 24, 164–171. [Google Scholar] [CrossRef]
- Facchinello, N.; Astone, M.; Audano, M.; Oberkersch, R.E.; Spizzotin, M.; Calura, E.; Marques, M.; Crisan, M.; Mitro, N.; Santoro, M.M. Oxidative pentose phosphate pathway controls vascular mural cell coverage by regulating extracellular matrix composition. Nat. Metab. 2022, 4, 123–140. [Google Scholar] [CrossRef]
- Kanungo, S.; Wells, K.; Tribett, T.; El-Gharbawy, A. Glycogen metabolism and glycogen storage disorders. Ann. Transl. Med. 2018, 6, 474. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor Cells | #3 | #4 | Total | |
|---|---|---|---|---|
| No. of embryos transferred | ||||
| Total | 3687 | 5458 | 9145 | |
| Mean ± SD | 335.2 ± 20.5 | 321.1 ± 12.9 | 326.6 ± 11.1 | |
| No. of surrogates | ||||
| Total | 11 | 17 | 28 | |
| Delivered (%) | 4 (36.4) | 3 (17.7) | 7 (25.0) | |
| No. of offspring (stillborn) | 11 (4) | 12 (3) | 23 (7) | |
| No. of offspring/embryos (%) | 11/3687 (0.3) | 12/5458 (0.2) | ND | |
| Mean weight of offspring (g) * | 964.1 ± 70.1 a | 826.7 ± 58.1 b | ND | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.-R.; Ahn, J.S.; Lee, M.G.; Lee, B.R.; Ock, S.A.; Byun, S.J.; Hwang, I.-S. Characterization of Enlarged Tongues in Cloned Piglets. Curr. Issues Mol. Biol. 2023, 45, 9103-9116. https://doi.org/10.3390/cimb45110571
Park M-R, Ahn JS, Lee MG, Lee BR, Ock SA, Byun SJ, Hwang I-S. Characterization of Enlarged Tongues in Cloned Piglets. Current Issues in Molecular Biology. 2023; 45(11):9103-9116. https://doi.org/10.3390/cimb45110571
Chicago/Turabian StylePark, Mi-Ryung, Jin Seop Ahn, Min Gook Lee, Bo Ram Lee, Sun A Ock, Sung June Byun, and In-Sul Hwang. 2023. "Characterization of Enlarged Tongues in Cloned Piglets" Current Issues in Molecular Biology 45, no. 11: 9103-9116. https://doi.org/10.3390/cimb45110571