
In Vitro Evidence of Differential Immunoregulatory Response between MDA-MB-231 and BT-474 Breast Cancer Cells Induced by Bone Marrow-Derived Mesenchymal Stromal Cells Conditioned Medium
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
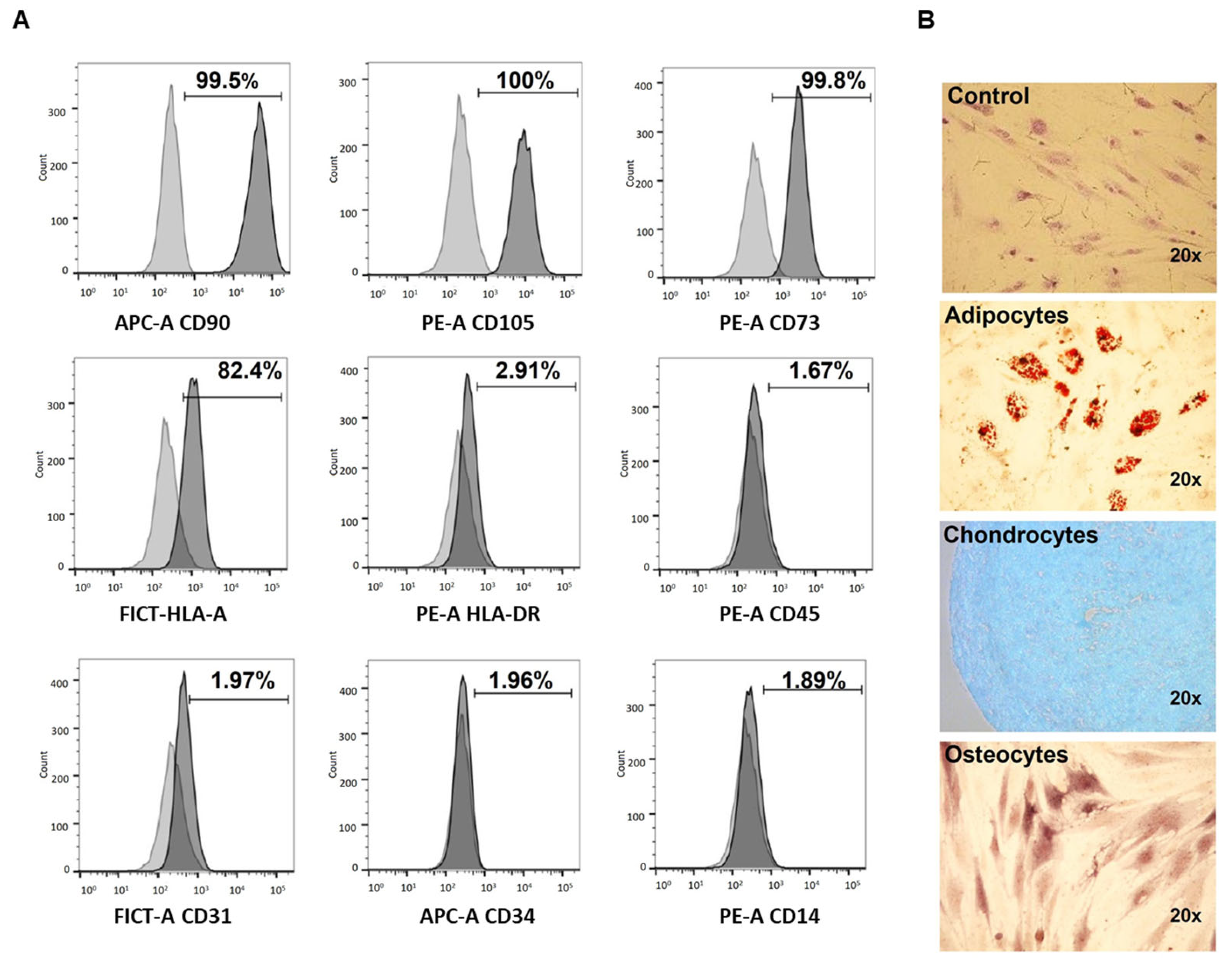
2.2. Characterization of hBM-MSC
2.2.1. Immunophenotype
2.2.2. Differentiation to Adipocytes
2.2.3. Differentiation to Chondrocytes
2.2.4. Differentiation to Osteocytes
2.3. Conditioned Media Preparation
2.4. Breast Cancer Cell Lines
2.5. qRT-PCR Assays
2.6. MNCs Proliferation Assays in Co-Cultures
2.7. Quantitative Analysis of Soluble Molecules
2.8. Detection of IDO Expression in Breast Cancer Cells
2.9. Detection of Treg Cells
2.10. Statistical Analyses
3. Results
3.1. Characterization of hBM-MSC
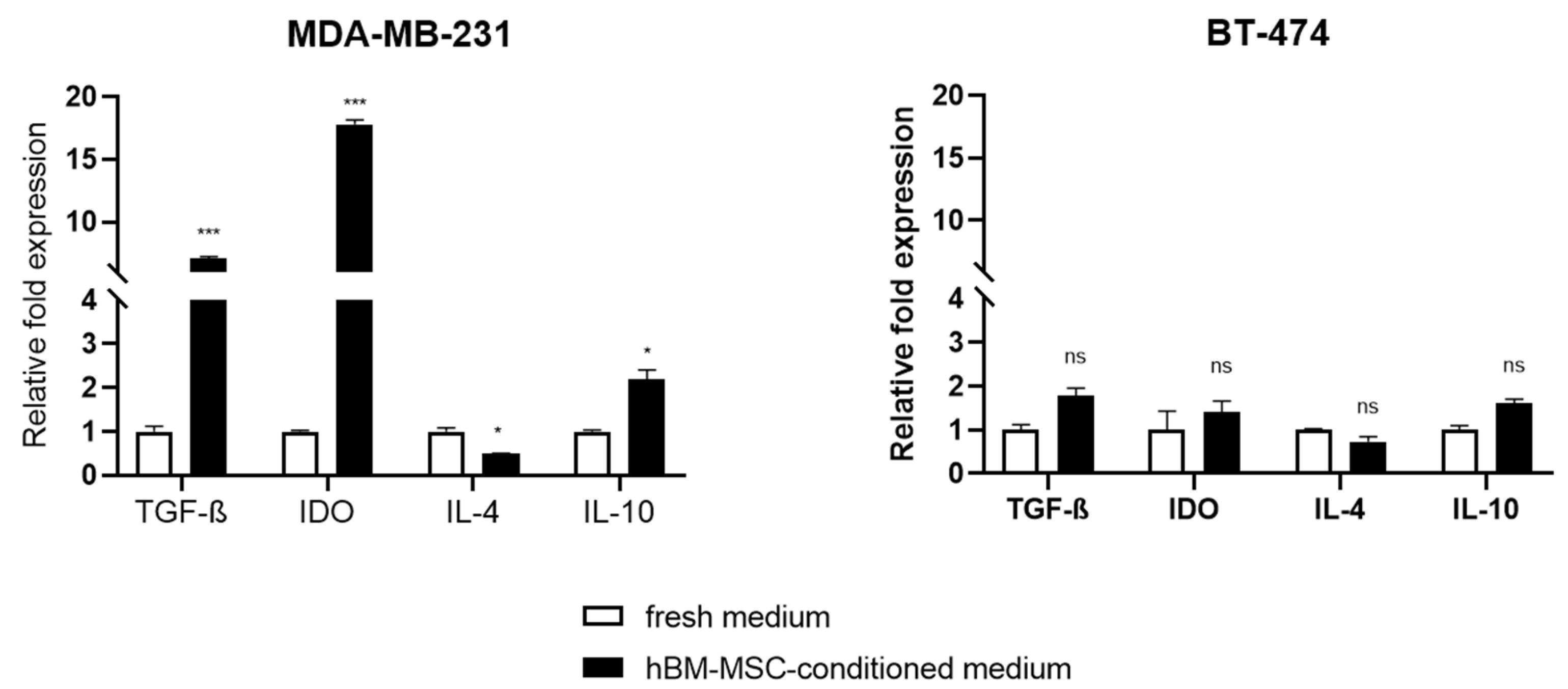
3.2. Effect of hBM-MSC-cm on the Expression of Immunomodulatory Genes in MDA-MB-231 and BT-474 Cells
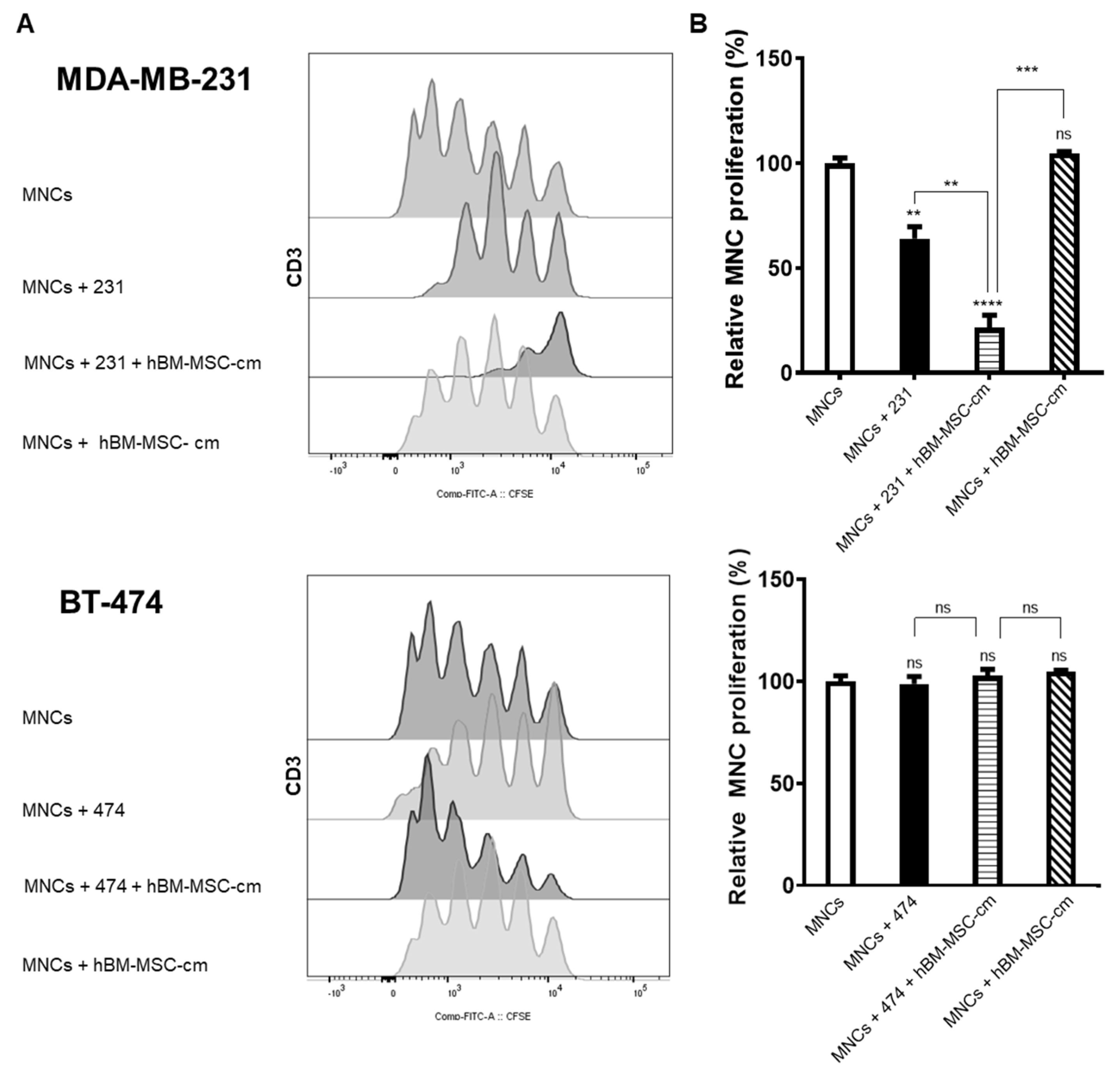
3.3. Proliferation Analysis of Mononuclear Cells Derived from Peripheral Blood as a Result in Co-Culture with Breast Cancer Cells Lines and hBM-MSC-cm
3.4. Evaluation of Candidate Molecules Involved in the Regulation of MNCs Proliferation
3.5. hBM-MSC-cm Differentially Induces IDO Expression on MDA-MB-231 Breast Cancer Cell Line
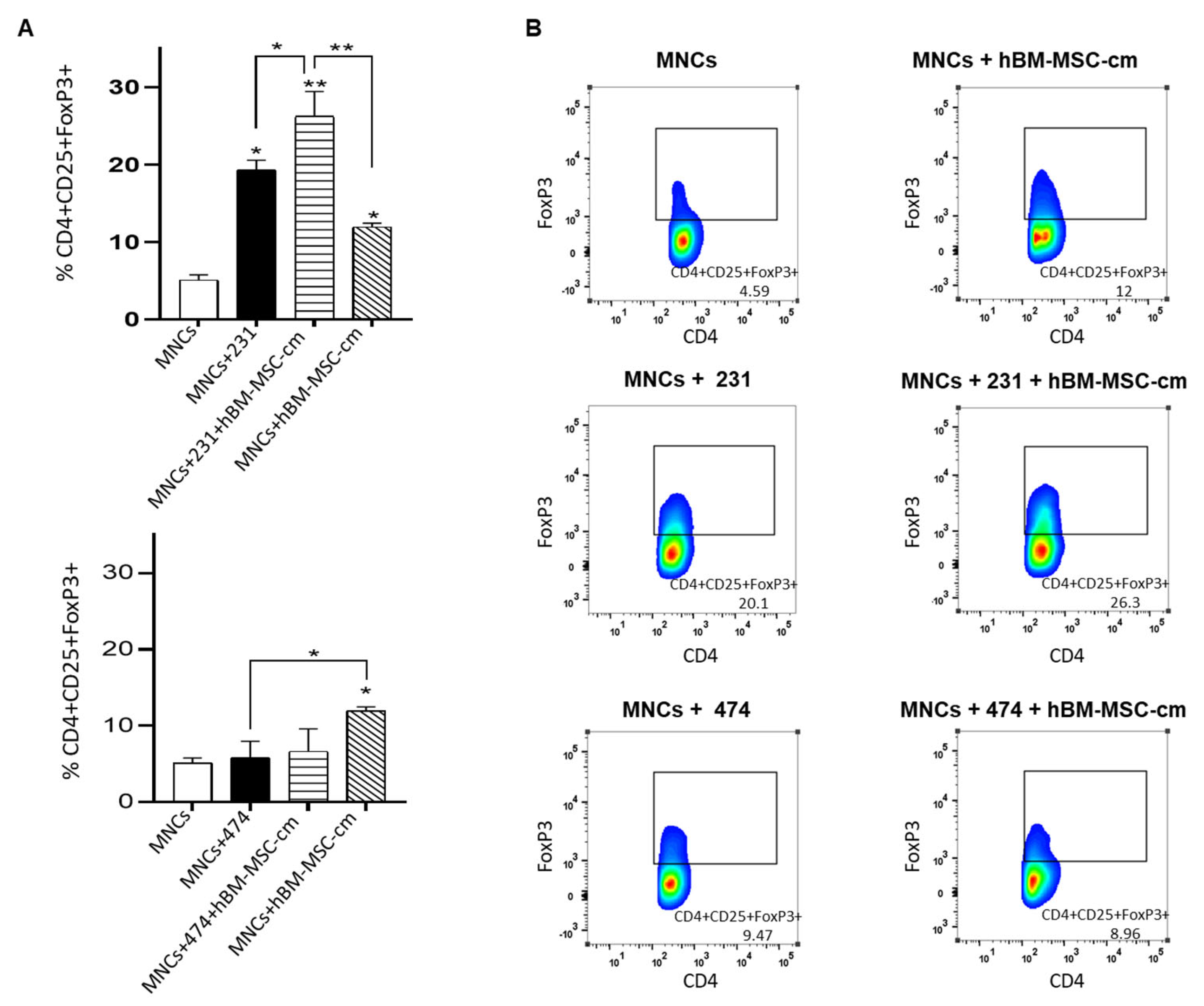
3.6. Evaluation of the Effect of hBM-MSC-cm on the Capability of MDA-MB-231 and BT-474 Breast Cancer Cells to Generate Regulatory T Lymphocytes (CD4+CD25+FoxP3+)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast Cancer. Nat. Rev. Dis. Prim. 2019, 5, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA J. Am. Med. Assoc. 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Wang, S.; Ali, N. Advanced Approaches to Breast Cancer Classification and Diagnosis. Front. Pharmacol. 2021, 11, 632079. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Li, T.; Bai, Z.; Yang, Y.; Liu, X.; Zhan, J.; Shi, B. Breast Cancer Intrinsic Subtype Classification, Clinical Use and Future Trends. Am. J. Cancer. Res. 2015, 5, 2929–2943. [Google Scholar] [PubMed]
- American Cancer Society. Breast Cancer Facts & Figures 2019–2020. Atlanta. 2019. Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/breast-cancer-facts-and-figures/breast-cancer-facts-and-figures-2019-2020.pdf (accessed on 8 December 2022).
- Noh, J.; Yu, J.; Kim, W.; Park, A.; Park, K.S. Bone Marrow-Derived Mesenchymal Stem Cells Migrate toward Hormone-Insensitive Prostate Tumor Cells Expressing TGF-β via N-Cadherin. Biomedicines 2021, 9, 1572. [Google Scholar] [CrossRef]
- Kidd, S.; Spaeth, E.; Dembinski, J.L.; Dietrich, M.; Watson, K.; Klopp, A.; Battula, V.L.; Weil, M.; Andreeff, M.; Marini, F.C. Direct Evidence of Mesenchymal Stem Cell Tropism for Tumor and Wounding Microenvironments Using in Vivo Bioluminescent Imaging. Stem Cells 2009, 27, 2614–2623. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Cheng, J.; Tang, Y.; Liu, Z.; Xu, C.; Liu, Y.; Sun, Y. Human Bone Marrow-Derived Mesenchymal Stem Cells Produced TGFbeta Contributes to Progression and Metastasis of Prostate Cancer. Cancer Investig. 2012, 30, 513–518. [Google Scholar] [CrossRef]
- Matsumoto, K.; Umitsu, M.; De Silva, D.M.; Roy, A.; Bottaro, D.P. Hepatocyte Growth Factor/MET in Cancer Progression and Biomarker Discovery. Cancer Sci. 2017, 108, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Farooqi, A.A.; Siddik, Z.H. Platelet-Derived Growth Factor (PDGF) Signalling in Cancer: Rapidly Emerging Signalling Landscape. Cell Biochem. Funct. 2015, 33, 257–265. [Google Scholar] [CrossRef]
- Spaeth, E.; Klopp, A.; Dembinski, J.; Andreeff, M.; Marini, F. Inflammation and Tumor Microenvironments: Defining the Migratory Itinerary of Mesenchymal Stem Cells. Gene Ther. 2008, 15, 730–738. [Google Scholar] [CrossRef]
- Wang, L.; Wang, X.; Wang, L.; Chiu, J.D.; Van De Ven, G.; Gaarde, W.A.; Deleve, L.D. Hepatic Vascular Endothelial Growth Factor Regulates Recruitment of Rat Liver Sinusoidal Endothelial Cell Progenitor Cells. Gastroenterology 2012, 143, 1555–1563. [Google Scholar] [CrossRef] [Green Version]
- Sugihara, H.; Ishimoto, T.; Yasuda, T.; Izumi, D.; Eto, K.; Sawayama, H.; Miyake, K.; Kurashige, J.; Imamura, Y.; Hiyoshi, Y.; et al. Cancer-Associated Fibroblast-Derived CXCL12 Causes Tumor Progression in Adenocarcinoma of the Esophagogastric Junction. Med. Oncol. 2015, 32, 618. [Google Scholar] [CrossRef]
- Shi, Y.; Du, L.; Lin, L.; Wang, Y. Tumour-Associated Mesenchymal Stem/Stromal Cells: Emerging Therapeutic Targets. Nat. Rev. Drug Discov. 2016, 16, 35–52. [Google Scholar] [CrossRef]
- Direkze, N.C.; Forbes, S.J.; Brittan, M.; Hunt, T.; Jeffery, R.; Preston, S.L.; Poulsom, R.; Hodivala-Dilke, K.; Alison, M.R.; Wright, N.A. Multiple Organ Engraftment by Bone-Marrow-Derived Myofibroblasts and Fibroblasts in Bone-Marrow-Transplanted Mice. Stem Cells 2003, 21, 514–520. [Google Scholar] [CrossRef]
- AHN, S.Y. The Role of MSCs in the Tumor Microenvironment and Tumor Progression. Anticancer Res. 2020, 40, 3039–3047. [Google Scholar] [CrossRef]
- Sánchez, N.; Miranda, A.; Funes, J.M.; Hevia, G.; Pérez, R.; De León, J. Oncogenic Transformation Tunes the Cross-Talk between Mesenchymal Stem Cells and T Lymphocytes. Cell. Immunol. 2014, 289, 174–184. [Google Scholar] [CrossRef]
- O’Malley, G.; Heijltjes, M.; Houston, A.M.; Rani, S.; Ritter, T.; Egan, L.J.; Ryan, A.E. Mesenchymal Stromal Cells (MSCs) and Colorectal Cancer—A Troublesome Twosome for the Anti-Tumour Immune Response? Oncotarget 2016, 7, 60752–60774. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by Mesenchymal Stem Cells (MSCs): Mechanisms of Action of Living, Apoptotic, and Dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.A.; Meyer, J.R.; Greco, S.J.; Corcoran, K.E.; Bryan, M.; Rameshwar, P. Mesenchymal Stem Cells Protect Breast Cancer Cells through Regulatory T Cells: Role of Mesenchymal Stem Cell-Derived TGF-β. J. Immunol. 2010, 184, 5885–5894. [Google Scholar] [CrossRef]
- Razmkhah, M.; Jaberipour, M.; Erfani, N.; Habibagahi, M.; Talei, A.-R.; Ghaderi, A. Adipose Derived Stem Cells (ASCs) Isolated from Breast Cancer Tissue Express IL-4, IL-10 and TGF-Β1 and Upregulate Expression of Regulatory Molecules on T Cells: Do They Protect Breast Cancer Cells from the Immune Response? Cell. Immunol. 2011, 266, 116–122. [Google Scholar] [CrossRef]
- Mansour, I.; Zayed, R.A.; Said, F.; Latif, L.A. Indoleamine 2,3-Dioxygenase and Regulatory T Cells in Acute Myeloid Leukemia. Hematology 2016, 21, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Kadle, R.L.; Abdou, S.A.; Villarreal-Ponce, A.P.; Soares, M.A.; Sultan, D.L.; David, J.A.; Massie, J.; Rifkin, W.J.; Rabbani, P.; Ceradini, D.J. Microenvironmental Cues Enhance Mesenchymal Stem Cell-Mediated Immunomodulation and Regulatory T-Cell Expansion. PLoS ONE 2018, 13, e0193178. [Google Scholar] [CrossRef]
- McGranahan, N.; Rosenthal, R.; Hiley, C.T.; Rowan, A.J.; Watkins, T.B.K.; Wilson, G.A.; Birkbak, N.J.; Veeriah, S.; Van Loo, P.; Herrero, J.; et al. Allele-Specific HLA Loss and Immune Escape in Lung Cancer Evolution. Cell 2017, 171, 1259–1271. [Google Scholar] [CrossRef] [Green Version]
- Mittal, D.; Gubin, M.M.; Schreiber, R.D.; Smyth, M.J. New Insights into Cancer Immunoediting and Its Three Component Phases—Elimination, Equilibrium and Escape. Curr. Opin. Immunol. 2014, 27, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Di, G. Role of Tumor Microenvironment in Triple-Negative Breast Cancer and Its Prognostic Significance. Chin. J. Cancer Res. 2017, 29, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Soysal, S.D.; Tzankov, A.; Muenst, S.E. Role of the Tumor Microenvironment in Breast Cancer. Pathobiology 2015, 82, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, W.; Valentin, J.E.; Marra, K.G.; Donnenberg, A.D.; Donnenberg, V.S.; Rubin, J.P. An Animal Model of Local Breast Cancer Recurrence in the Setting of Autologous Fat Grafting for Breast Reconstruction. Stem Cells Transl. Med. 2018, 7, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Sepehr, K.S.; Razavi, A.; Hassan, Z.M.; Fazel, A.; Abdollahpour-Alitappeh, M.; Mossahebi-Mohammadi, M.; Yekaninejad, M.S.; Farhadihosseinabadi, B.; Hashemi, S.M. Comparative Immunomodulatory Properties of Mesenchymal Stem Cells Derived from Human Breast Tumor and Normal Breast Adipose Tissue. Cancer Immunol. Immunother. 2020, 69, 1841–1854. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- De la Rosa-Ruiz, M.D.P.; Álvarez-Pérez, M.A.; Cortés-Morales, V.A.; Monroy-García, A.; Mayani, H.; Fragoso-González, G.; Caballero-Chacón, S.; Diaz, D.; Candanedo-González, F.; Montesinos, J.J. Mesenchymal Stem/Stromal Cells Derived from Dental Tissues: A Comparative In Vitro Evaluation of Their Immunoregulatory Properties Against T Cells. Cells 2019, 8, 1491. [Google Scholar] [CrossRef] [Green Version]
- Saphner, T.; Tormey, D.C.; Gray, R. Annual Hazard Rates of Recurrence for Breast Cancer after Primary Therapy. J. Clin. Oncol. 1996, 14, 2738–2746. [Google Scholar] [CrossRef]
- Wei, L.; Zhu, S.; Li, M.; Li, F.; Wei, F.; Liu, J.; Ren, X. High Indoleamine 2,3-Dioxygenase Is Correlated with Microvessel Density and Worse Prognosis in Breast Cancer. Front. Immunol. 2018, 9, 724. [Google Scholar] [CrossRef]
- Llanes-Fernández, L.; Álvarez-Goyanes, R.I.; Arango-Prado, M.d.C.; Alcocer-González, J.M.; Mojarrieta, J.C.; Pérez, X.E.; López, M.O.; Odio, S.F.; Camacho-Rodríguez, R.; Guerra-Yi, M.E.; et al. Relationship between IL-10 and Tumor Markers in Breast Cancer Patients. Breast 2006, 15, 482–489. [Google Scholar] [CrossRef]
- Zarzynska, J.M. Two Faces of TGF-Beta1 in Breast Cancer. Mediators Inflamm. 2014, 2014, 141774. [Google Scholar] [CrossRef]
- Lewis, A.M.; Varghese, S.; Xu, H.; Alexander, H.R. Interleukin-1 and Cancer Progression: The Emerging Role of Interleukin-1 Receptor Antagonist as a Novel Therapeutic Agent in Cancer Treatment. J. Transl. Med. 2006, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Massagué, J. TGFβ in Cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Batlle, E.; Massagué, J. Transforming Growth Factor-β Signaling in Immunity and Cancer. Immunity 2019, 50, 924–940. [Google Scholar] [CrossRef]
- Sheikhpour, E.; Noorbakhsh, P.; Foroughi, E.; Farahnak, S.; Nasiri, R.; Neamatzadeh, H. A Survey on the Role of Interleukin-10 in Breast Cancer: A Narrative. Rep. Biochem. Mol. Biol. 2017, 7, 30–37. [Google Scholar]
- Levings, M.K.; Bacchetta, R.; Schulz, U.; Roncarolo, M.G. The Role of IL-10 and TGF-Beta in the Differentiation and Effector Function of T Regulatory Cells. Int. Arch. Allergy Immunol. 2002, 129, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Dewi, D.L.; Mohapatra, S.R.; Cabañes, S.B.; Adam, I.; Patterson, L.F.S.; Berdel, B.; Kahloon, M.; Thürmann, L.; Loth, S.; Heilmann, K.; et al. Suppression of Indoleamine-2,3-Dioxygenase 1 Expression by Promoter Hypermethylation in ER-Positive Breast Cancer. Oncoimmunology 2017, 6, e1274477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, A.; Zhang, Y.; Xu, J.; Yin, T.; Lu, X.-J. T Cell Dysfunction in Cancer Immunity and Immunotherapy. Front. Immunol. 2019, 10, 1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özgül Özdemir, R.B.; Özdemir, A.T.; Kırmaz, C.; Tuğlu, M.İ.; Şenol, Ö.; Özverel, C.S.; Berdeli, A. The Effects of Mesenchymal Stem Cells on the IDO, HLA-G and PD-L1 Expression of Breast Tumor Cells MDA-MB-231 and MCF-7. Arch. Clin. Exp. Med. 2019, 4, 132–137. [Google Scholar] [CrossRef]
- Zheng, Y.; Fang, Y.C.; Li, J. Expression Levels on Tumor Cells Affect Their Immunosuppressive Activity. Oncol. Lett. 2019, 18, 5399–5407. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Tang, Y.; Hua, S. Immunological Approaches Towards Cancer and Inflammation: A Cross Talk. Front. Immunol. 2018, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Mirlekar, B. Tumor Promoting Roles of IL-10, TGF-β, IL-4, and IL-35: Its Implications in Cancer Immunotherapy. SAGE Open Med. 2022, 10, 205031212110690. [Google Scholar] [CrossRef]
- Dennis, K.L.; Blatner, N.R.; Gounari, F.; Khazaie, K. Current Status of Interleukin-10 and Regulatory T-Cells in Cancer. Curr. Opin. Oncol. 2013, 25, 637–645. [Google Scholar] [CrossRef]
- Fujii, S.-I.; Shimizu, K.; Shimizu, T.; Lotze, M.T. Interleukin-10 Promotes the Maintenance of Antitumor CD8(+) T-Cell Effector Function in Situ. Blood 2001, 98, 2143–2151. [Google Scholar] [CrossRef] [Green Version]
- Zhai, L.; Bell, A.; Ladomersky, E.; Lauing, K.L.; Bollu, L.; Sosman, J.A.; Zhang, B.; Wu, J.D.; Miller, S.D.; Meeks, J.J.; et al. Immunosuppressive IDO in Cancer: Mechanisms of Action, Animal Models, and Targeting Strategies. Front. Immunol. 2020, 11, 1185. [Google Scholar] [CrossRef]
- Isla Larrain, M.T.; Rabassa, M.E.; Lacunza, E.; Barbera, A.; Cretón, A.; Segal-Eiras, A.; Croce, M.V. IDO Is Highly Expressed in Breast Cancer and Breast Cancer-Derived Circulating Microvesicles and Associated to Aggressive Types of Tumors by in Silico Analysis. Tumor Biol. 2014, 35, 6511–6519. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Abomaray, F.M.; Alshabibi, M.A.; AlAskar, A.S.; Kalionis, B. Immunomodulatory Properties of Human Placental Mesenchymal Stem/Stromal Cells. Placenta 2017, 59, 87–95. [Google Scholar] [CrossRef]
- Yuan, X.L.; Chen, L.; Zhang, T.T.; Ma, Y.H.; Zhou, Y.L.; Zhao, Y.; Wang, W.W.; Dong, P.; Yu, L.; Zhang, Y.Y.; et al. Gastric Cancer Cells Induce Human CD4+Foxp3+ Regulatory T Cells through the Production of TGF-Β1. World J. Gastroenterol. 2011, 17, 2019. [Google Scholar] [CrossRef]
- Prat, A.; Parker, J.S.; Karginova, O.; Fan, C.; Livasy, C.; Herschkowitz, J.I.; He, X.; Perou, C.M. Phenotypic and Molecular Characterization of the Claudin-Low Intrinsic Subtype of Breast Cancer. Breast Cancer Res. 2010, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Casbas-Hernandez, P.; Sun, X.; Roman-Perez, E.; D’Arcy, M.; Sandhu, R.; Hishida, A.; McNaughton, K.K.; Yang, X.R.; Makowski, L.; Sherman, M.E.; et al. Tumor Intrinsic Subtype Is Reflected in Cancer-Adjacent Tissue. Cancer Epidemiol. Biomark. Prev. 2015, 24, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Coleman, R.E. Clinical Features of Metastatic Bone Disease and Risk of Skeletal Morbidity. Clin. Cancer Res. 2006, 12 Pt 2, 6243–6249. [Google Scholar] [CrossRef] [Green Version]
- Hess, K.R.; Varadhachary, G.R.; Taylor, S.H.; Wei, W.; Raber, M.N.; Lenzi, R.; Abbruzzese, J.L. Metastatic Patterns in Adenocarcinoma. Cancer 2006, 106, 1624–1633. [Google Scholar] [CrossRef]
- Czapla, J.; Matuszczak, S.; Kulik, K.; Wiśniewska, E.; Pilny, E.; Jarosz-Biej, M.; Smolarczyk, R.; Sirek, T.; Zembala, M.O.; Zembala, M.; et al. The Effect of Culture Media on Large-Scale Expansion and Characteristic of Adipose Tissue-Derived Mesenchymal Stromal Cells. Stem Cell Res. Ther. 2019, 10, 235. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Company | Catalog Number |
|---|---|---|
| CD90 PE-Cy5 | BD Biosciences, San Diego, CA, USA | 555597 |
| CD105 eFlour450 | Ebioscience, San Diego, CA, USA | 48-1057-42 |
| CD73 PE-Cy7 | BD Biosciences, San Diego, CA, USA | 561258 |
| HLA-A | Biolegend, San Diego, CA, USA | 555552 |
| HLA-DR PE-Cy-7 | Biolegend, San Diego, CA, USA | 307616 |
| CD45 PE | BD Biosciences, San Diego, CA, USA | 555483 |
| CD31 FITC | BD Biosciences, San Diego, CA, USA | 555445 |
| CD34 APC | BD Biosciences, San Diego, CA, USA | 555824 |
| CD 14 PE | BD Biosciences, San Diego, CA, USA | 555398 |
| CD3 PE | BD Biosciences, San Diego, CA, USA | 555333 |
| 7AAD | BD Biosciences, San Diego, CA, USA | 559925 |
| CD45 APC | BD Biosciences, San Diego, CA, USA | 559865 |
| IDO PE | RyD systems, Minneapolis, MN, USA | IC6030P |
| CD4 FITC | BD Biosciences San Diego, CA, USA | 555346 |
| CD25 PE | BD Biosciences San Diego, CA, USA | 555432 |
| FoxP3 PECy/7 | Ebioscience, San Diego, CA, USA | 25-4777-42 |
| Probe | Sequence | Amplicon Length (bp) |
|---|---|---|
| GAPDH | F FGGTGTGAACCATGAGAAGTATGA R GAGTCCTTCCACGA TACCAAAG | 123 |
| IDO | F AGGATTCTTCCTGGTCTCTCT R GTGTCCCGTTCTTGCATTTG | 102 |
| TGF-β | F CGTGGAGCTGTACCAGAAATAC R CACAACTCCGGTGACATCAA | 112 |
| IL-10 | F GCTGGAGGACTTTAAGGGTTAC R GATGTCTGGGTCTTGGTTCTC | 106 |
| IL-4 | F GTTCTACAGCCACCATGAGAA R CCGTTTCAGGAATCGGATCA | 94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arenas-Luna, V.M.; Montesinos, J.J.; Cortés-Morales, V.A.; Navarro-Betancourt, J.R.; Peralta-Ildefonso, J.; Cisneros, B.; Hernández-Gutiérrez, S. In Vitro Evidence of Differential Immunoregulatory Response between MDA-MB-231 and BT-474 Breast Cancer Cells Induced by Bone Marrow-Derived Mesenchymal Stromal Cells Conditioned Medium. Curr. Issues Mol. Biol. 2023, 45, 268-285. https://doi.org/10.3390/cimb45010020
Arenas-Luna VM, Montesinos JJ, Cortés-Morales VA, Navarro-Betancourt JR, Peralta-Ildefonso J, Cisneros B, Hernández-Gutiérrez S. In Vitro Evidence of Differential Immunoregulatory Response between MDA-MB-231 and BT-474 Breast Cancer Cells Induced by Bone Marrow-Derived Mesenchymal Stromal Cells Conditioned Medium. Current Issues in Molecular Biology. 2023; 45(1):268-285. https://doi.org/10.3390/cimb45010020
Chicago/Turabian StyleArenas-Luna, Víctor M., Juan J. Montesinos, Víctor A. Cortés-Morales, José R. Navarro-Betancourt, Janneth Peralta-Ildefonso, Bulmaro Cisneros, and Salomón Hernández-Gutiérrez. 2023. "In Vitro Evidence of Differential Immunoregulatory Response between MDA-MB-231 and BT-474 Breast Cancer Cells Induced by Bone Marrow-Derived Mesenchymal Stromal Cells Conditioned Medium" Current Issues in Molecular Biology 45, no. 1: 268-285. https://doi.org/10.3390/cimb45010020