
Severe Acute Respiratory Syndrome Coronavirus-2 Delta Variant Study In Vitro and Vivo

 and
and
Abstract
:1. Introduction
2. Methods
2.1. Virus
2.2. Cells
2.3. Quantitative RT-PCR
2.4. Nanopore Sequencing
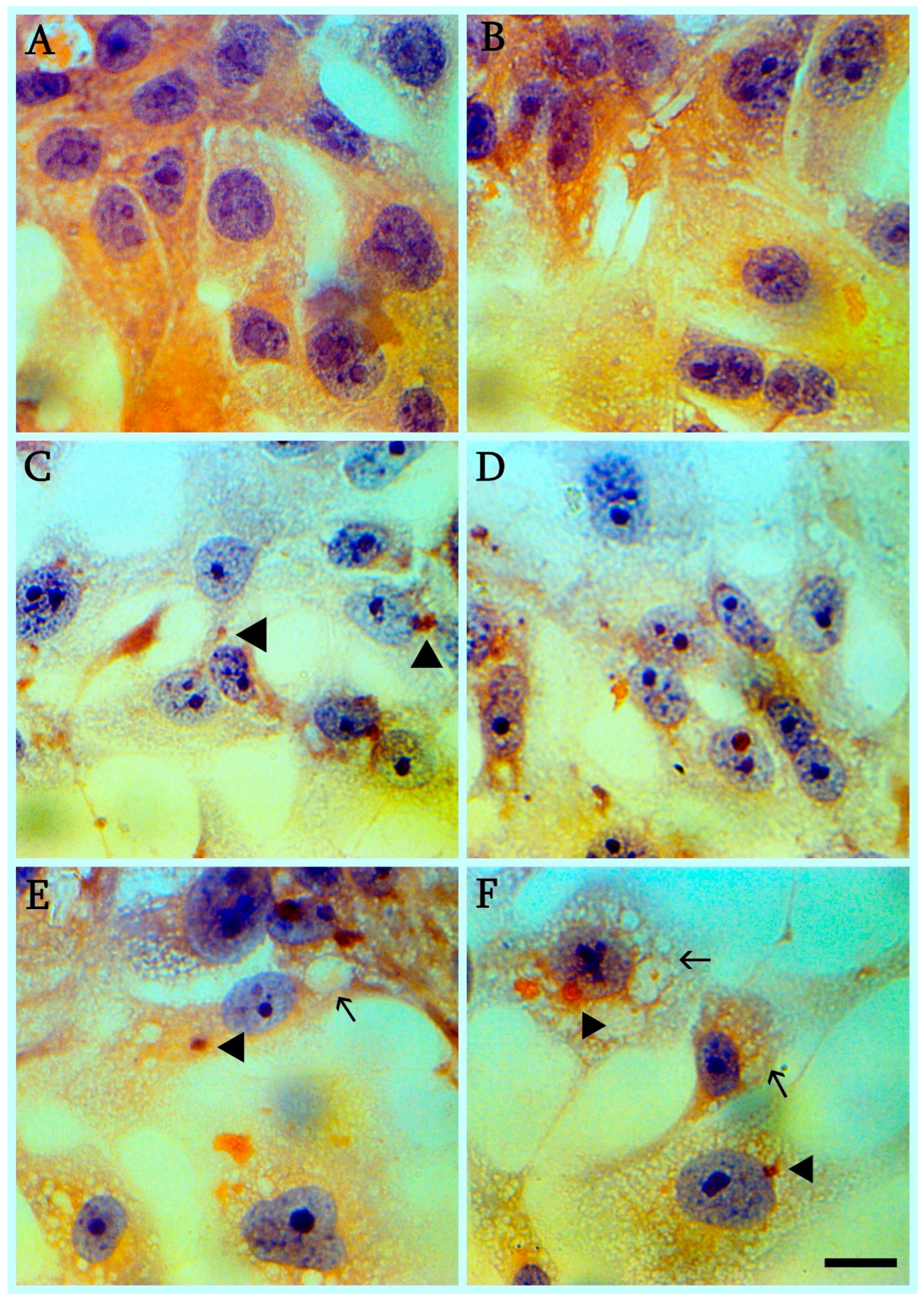
2.5. Staining
2.6. Experimental Infection of Syrian Hamsters
2.7. Bone Marrow Imprints and Cell Analysis
2.8. Tissue Samples
2.9. Micro-CT Imaging
2.10. Statistical Analysis
3. Results
3.1. Isolation and Adaptation of SARS-CoV-2Delta Variant
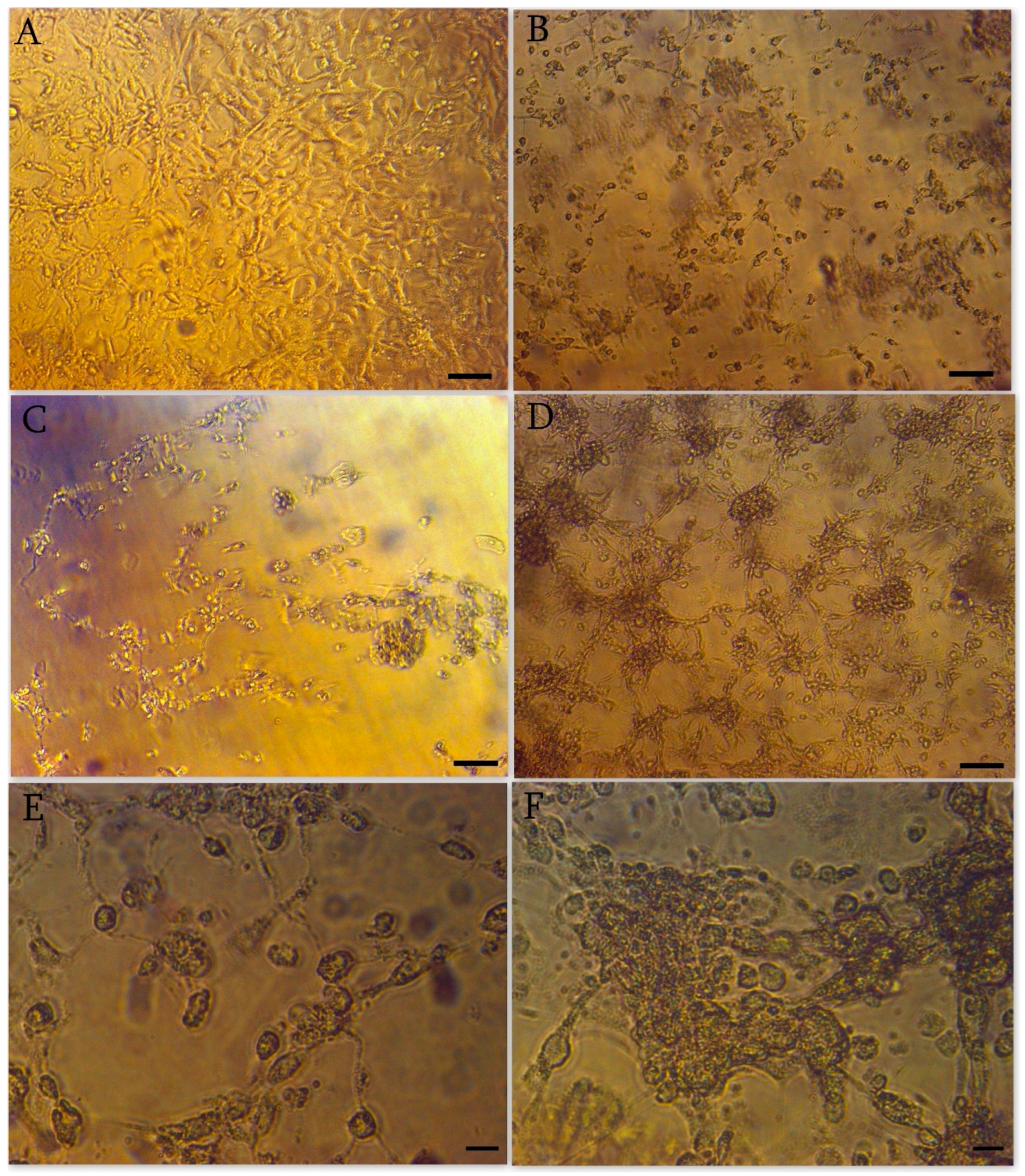
3.2. Cytopathic Effect
3.3. SARS-CoV-2 Quantification In Vitro
3.4. Changes in the SARS-CoV-2 Delta Variant Genome at Adaptation to Replication in VERO Cell
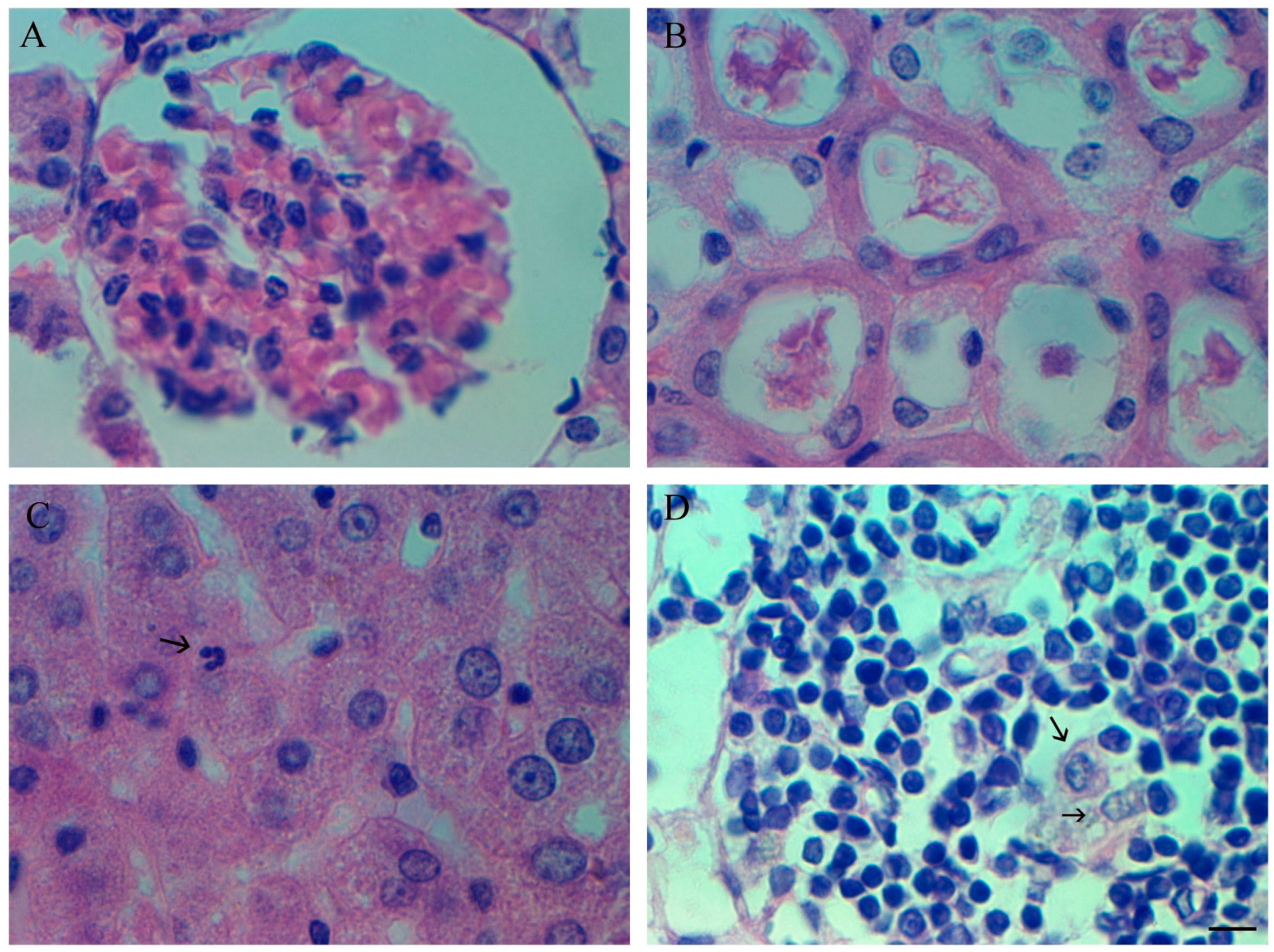
3.5. Changes in Blood Cell Populations in Syrian Golden Hamsters
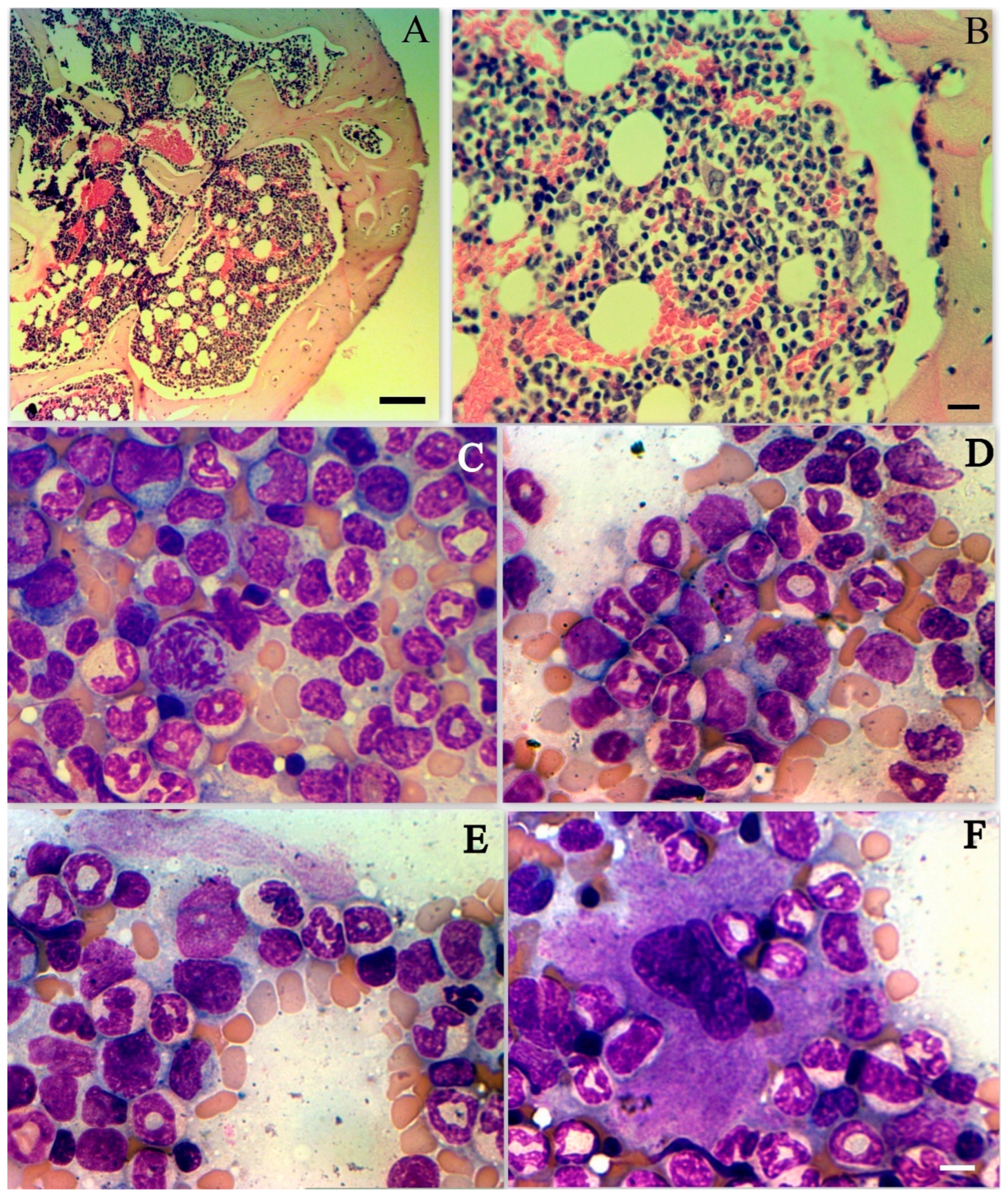
3.6. Changes in Bone Marrow Cell Populations
3.7. SARS-CoV-2 Infection in Syrian Golden Hamsters
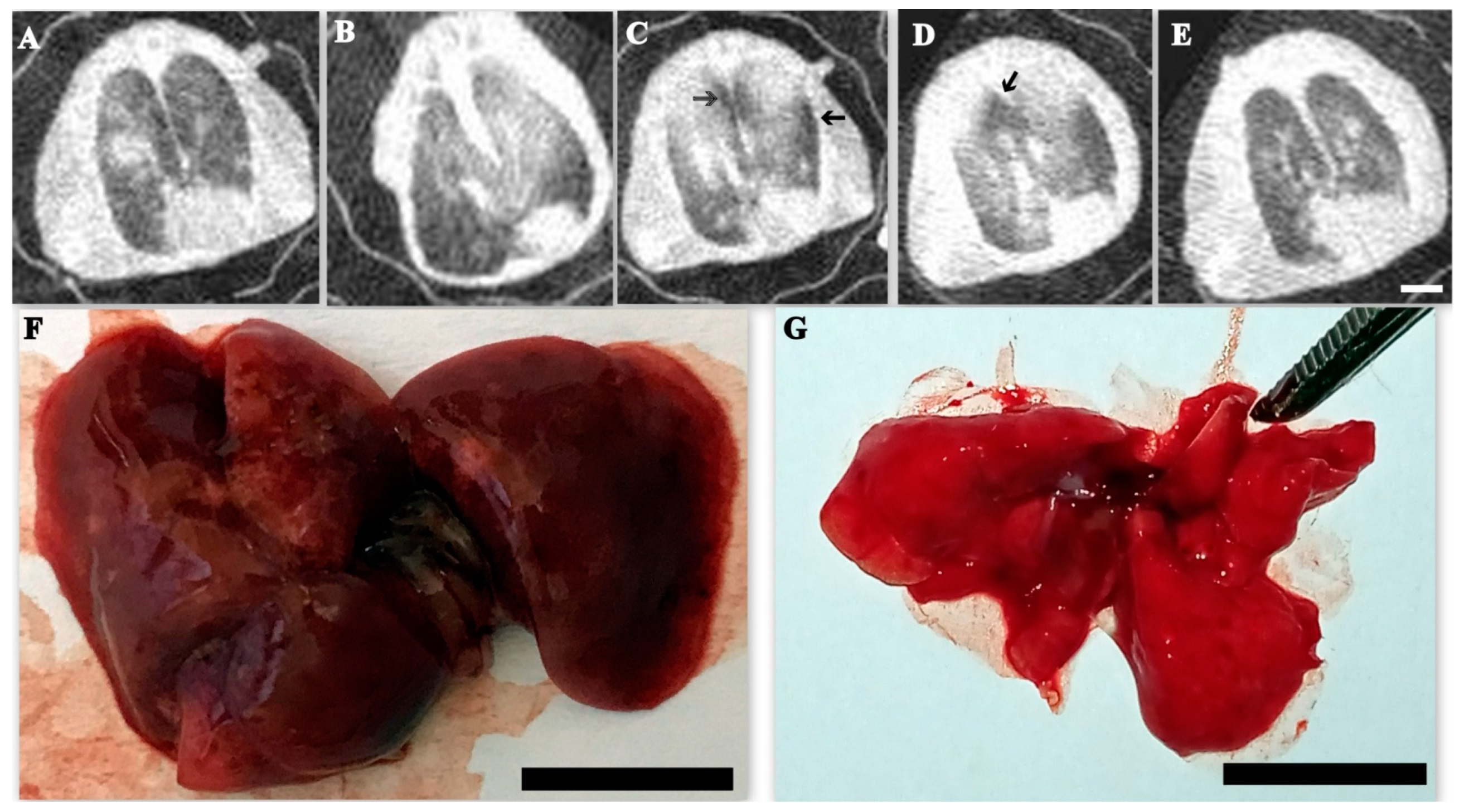
3.8. Lung Computed Tomography and Gross Pathology
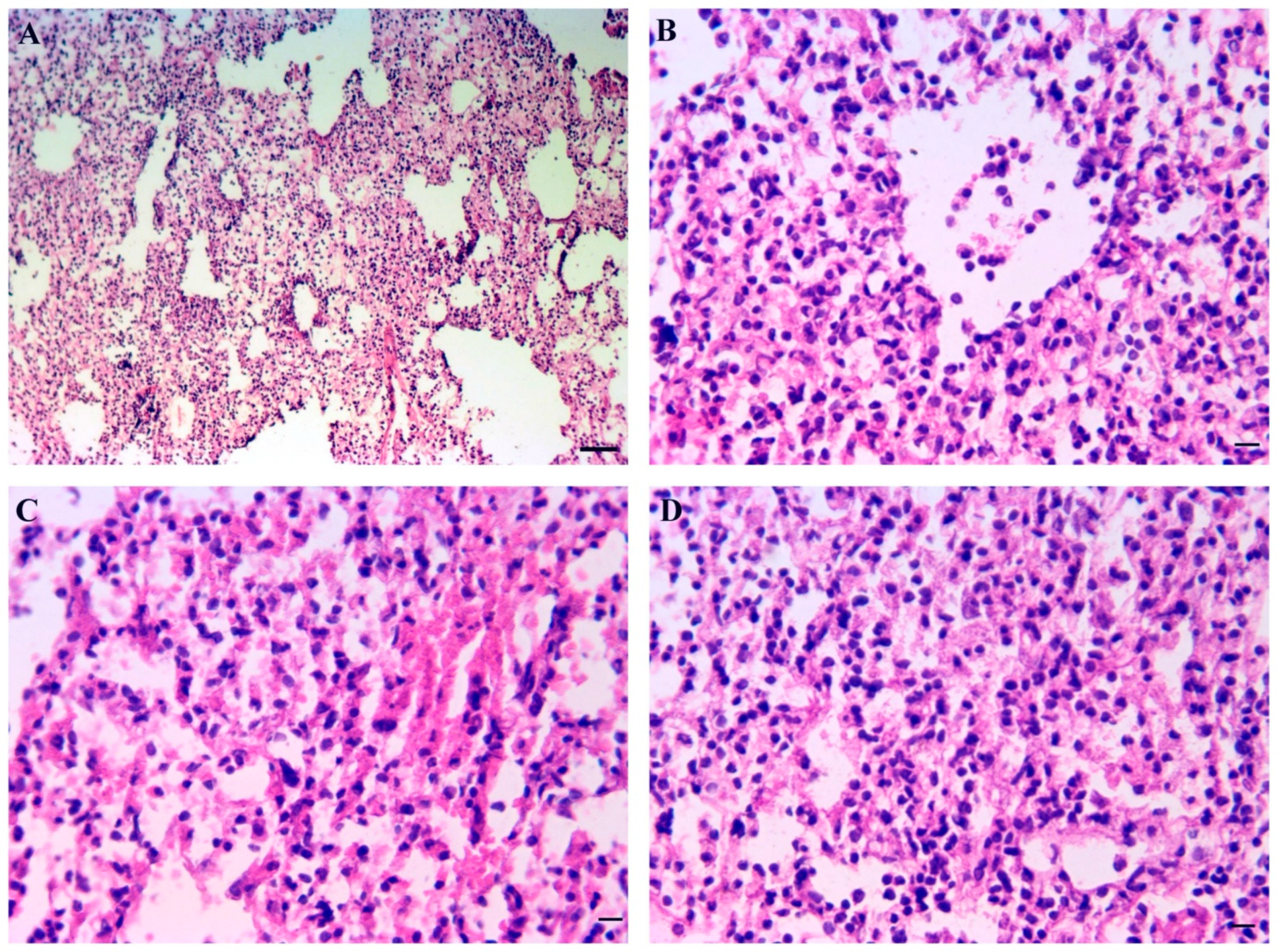
3.9. Inner Organs Histopathology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.T.; Leung, K.; Leung, G.M. Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: A modelling study. Lancet 2020, 395, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.C.; Shih, T.P.; Ko, W.C.; Tang, H.J.; Hsueh, P.R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, A.; Caccuri, F.; Bugatti, A.; Zani, A.; Vanoni, M.; Bonfanti, P.; Cazzaniga, M.E.; Perno, C.F.; Messa, C.; Alberghina, L. Methotrexate inhibits SARS-CoV-2 virus replication “in vitro”. J. Med. Virol. 2021, 93, 1780–1785. [Google Scholar] [CrossRef]
- Dalebout, T.J.; Zevenhoven-Dobbe, J.C.; Limpens, R.W.A.L.; van der Meer, Y.; Caly, L.; Druce, J.; de Vries, J.J.C.; Kikkert, M.; Bárcena, M.; Sidorov, I.; et al. SARS-coronavirus-2 replication in Vero E6 cells: Replication kinetics, rapid adaptation and cytopathology. J. Gen. Virol. 2020, 101, 925–940. [Google Scholar] [CrossRef]
- Kumar, S.; Thambiraja, T.S.; Karuppanan, K.; Subramaniam, G. Omicron and Delta variant of SARS-CoV-2: A comparative computational study of spike protein. J. Med. Virol. 2022, 94, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Zhang, A.J.; Yuan, S.; Poon, V.K.; Chan, C.C.; Lee, A.C.; Chan, W.M.; Fan, Z.; Tsoi, H.W.; Wen, L.; et al. Simulation of the Clinical and Pathological Manifestations of Coronavirus Disease 2019 (COVID-19) in a Golden Syrian Hamster Model: Implications for Disease Pathogenesis and Transmissibility. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, 2428–2446. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.; Zhang, A.J.; Chan, J.F.; Li, C.; Fan, Z.; Liu, F.; Chen, Y.; Liang, R.; Sridhar, S.; Cai, J.P.; et al. Oral SARS-CoV-2 Inoculation Establishes Subclinical Respiratory Infection with Virus Shedding in Golden Syrian Hamsters. Cell Rep. Med. 2020, 1, 100121. [Google Scholar] [CrossRef]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Dowling, W.E.; Funnell, S.; Gsell, P.S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.; Fung, K.; Choy, K.T.; Wong, A.; Kaewpreedee, P.; Perera, R.; Poon, L.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Bao, L.; Yu, P.; Qi, F.; Gong, S.; Wang, J.; Zhao, B.; Liu, M.; Han, Y.; Deng, W.; et al. SARS-CoV-2 Causes a Systemically Multiple Organs Damages and Dissemination in Hamsters. Front. Microbiol. 2021, 11, 618891. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, M.; Saito, J.; Zhao, H.; Sakamoto, A.; Hirota, K.; Ma, D. Inflammation Triggered by SARS-CoV-2 and ACE2 Augment Drives Multiple Organ Failure of Severe COVID-19: Molecular Mechanisms and Implications. Inflammation 2021, 44, 13–34. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wu, J.; Nie, J.; Zhang, L.; Hao, H.; Liu, S.; Zhao, C.; Zhang, Q.; Liu, H.; Nie, L.; et al. The Impact of Mutations in SARS-CoV-2 Spike on Viral Infectivity and Antigenicity. Cell 2020, 182, 1284–1294.e9. [Google Scholar] [CrossRef]
- Francis, M.E.; Goncin, U.; Kroeker, A.; Swan, C.; Ralph, R.; Lu, Y.; Etzioni, A.L.; Falzarano, D.; Gerdts, V.; Machtaler, S.; et al. SARS-CoV-2 infection in the Syrian hamster model causes inflammation as well as type I interferon dysregulation in both respiratory and non-respiratory tissues including the heart and kidney. PLoS Pathog. 2021, 17, e1009705. [Google Scholar] [CrossRef]
- Arakelyan, A.; Avetyan, D.; Karalyan, Z.; Akopyan, S.; Nikogosyan, M.; Khachatryan, G.; Sirunyan, T.; Gukasyan, L.; Zakharyan, R.; Muradyan, N.; et al. Molecular Genetics and Pathology of COVID-19. In Proceedings of the Compedium of the II International Scientific and Practical Conference on Counteraction of the Coronavirus Infection and Other Infectious Diseases, Saint-Petersburg, Russia, 9–10 December 2021; pp. 13–16. [Google Scholar]
- Avetyan, D.; Chavushyan, A.; Ghazaryan, H.; Melkonyan, A.; Stepanyan, A.; Zakharyan, R.; Hayrapetyan, V.; Atshemyan, S.; Khachatryan, G.; Sirunyan, T.; et al. SARS-CoV-2 detection by extraction-free qRT-PCR for massive and rapid COVID-19 diagnosis during a pandemic in Armenia. J. Virol. Methods 2021, 295, 114199. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Liu, Q.; Yu, L.; Wu, S.; Yin, X. Sheng Wu Gong Cheng Xue Bao. Chin. J. Biotechnol. 2020, 36, 732–739. [Google Scholar] [CrossRef]
- Tyson, J.R.; James, P.; Stoddart, D.; Sparks, N.; Wickenhagen, A.; Hall, G.; Choi, J.H.; Lapointe, H.; Kamelian, K.; Smith, A.D.; et al. Improvements to the ARTIC multiplex PCR method for SARS-CoV-2 genome sequencing using nanopore. bioRxiv 2020. [Google Scholar] [CrossRef]
- Loman, N.J.; Quick, J.; Simpson, J.T. A complete bacterial genome assembled de novo using only nanopore sequencing data. Nat. Methods 2015, 12, 733–735. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaryan, H.; Cholakyans, V.; Simonyan, L.; Misakyan, A.; Karalova, E.; Chavushyan, A.; Karalyan, Z. A study of lymphoid organs and serum proinflammatory cytokines in pigs infected with African swine fever virus genotype II. Arch. Virol. 2015, 160, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Munster, V.; Jarvis, M.A.; Feldmann, H. Defining the Syrian hamster as a highly susceptible preclinical model for SARS-CoV-2 infection. bioRxiv 2020, in press. [Google Scholar] [CrossRef]
- Braxton, A.M.; Creisher, P.S.; Ruiz-Bedoya, C.A.; Mulka, K.R.; Dhakal, S.; Ordonez, A.A.; Beck, S.E.; Jain, S.K.; Villano, J.S. Hamsters as a Model of Severe Acute Respiratory Syndrome Coronavirus-2. Comp. Med. 2021, 71, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Rosenke, K.; Meade-White, K.; Letko, M.; Clancy, C.; Hansen, F.; Liu, Y.; Okumura, A.; Tang-Huau, T.L.; Li, R.; Saturday, G.; et al. Defining the Syrian hamster as a highly susceptible preclinical model for SARS-CoV-2 infection. Emerg. Microbes Infect. 2020, 9, 2673–2684. [Google Scholar] [CrossRef]
- Hsu, C.J.; Lin, W.C.; Chou, Y.C.; Yang, C.M.; Wu, H.L.; Cheng, Y.H.; Liu, P.C.; Chang, J.Y.; Chen, H.Y.; Sun, J.R. Dynamic Changes of the Blood Chemistry in Syrian Hamsters Post-Acute COVID-19. Microbiol. Spectr. 2022, 10, e0236221. [Google Scholar] [CrossRef]
- Ohno, M.; Sasaki, M.; Orba, Y.; Sekiya, T.; Masum, M.A.; Ichii, O.; Sawamura, T.; Kakino, A.; Suzuki, Y.; Kida, H.; et al. Abnormal Blood Coagulation and Kidney Damage in Aged Hamsters Infected with Severe Acute Respiratory Syndrome Coronavirus 2. Viruses 2021, 13, 2137. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Positions | Virus | Gene, Product | Sequence |
|---|---|---|---|
| 809 | Original virus Virus after early stage of adaptation on Vero cells | gene = “ORF1ab product = “nsp2 | AGGGCATACACTC AGGGCACACACTC |
| 14,790 | Original virus Virus genome after adaptation on Vero cells | gene = “ORF1ab product = “RNA-dependent RNA polymerase | TGCTATCAGCGAT TGCTATTAGCGAT |
| 14,925 | Original virus Virus genome after adaptation on Vero cells | gene = “ORF1ab product = “RNA-dependent RNA polymerase | CATCGTTAACAAC CATCGTCAACAAC |
| 15,952 | Original virus Virus genome after adaptation on Vero cells | gene = “ORF1ab product = “RNA-dependent RNA polymerase | AGAATCATAGGGG AGAATCCTAGGGG |
| 18,117 | Original virus Virus after adaptation on Vero cells | gene = “ORF1ab” product = “3’-to-5’ exonuclease | TACACATCTCAGT TACACACCTCAGT |
| 18,744 | Original virus Virus genome after adaptation on Vero cells | gene = “ORF1ab” product = “3’-to-5’ exonuclease | TGATTATGTCTAT TGATTACGTCTAT |
| 21,137 | Original virus Virus genome after adaptation on Vero cells | gene = “ORF1ab product = “2’-O-ribose methyltransferase | AACAAAGGCTAGC AACAAAAGCTAGC |
| 23,803 | Original virus Virus after adaptation on Vero cells | Gene S spike glycoprotein product surface glycoprotein | TTCAACCGAATGC TTCAACTGAATGC |
| 24,410 | Original virus Virus after adaptation on Vero cells | Gene S spike glycoprotein product surface glycoprotein | GACCTCCTTTGTG GACCTCATTTGTG |
| 25,117 | Original virus Virus after adaptation on Vero cells | Gene S spike glycoprotein product surface glycoprotein | TGACCGTCTCAAT TGACCGCCTCAAT |
| 25,439 | Original virus Virus after adaptation on Vero cells | gene= ORF3a product ORF3a protein | CTTTGACGCAAGG CTTTGAAGCAAGG |
| 25,913 | original virus Virus after adaptation on Vero cells | gene = ORF3a product ORF3a protein | GTGATGACACAAC GTGATGGCACAAC |
| 26,885 | original virus Virus after adaptation on Vero cells | gene = M product membrane glycoprotein | TCTCAATGTGCCA TCTCAACGTGCCA |
| 28,299 | original virus Virus after adaptation on Vero cells | gene = N product = “nucleocapsid phosphoprotein | AAAATCTGCGAAA AAAATCAGCGAAA |
| 28,697 | original virus Virus after adaptation on Vero cells | gene = N product = nucleocapsid phosphoprotein | AATACATCAAAAG AATACACCAAAAG |
| 28,748 | original virus Virus after adaptation on Vero cells | gene = N product = nucleocapsid phosphoprotein | ATCGTGTTACAAC ATCGTGCTACGAC |
| Cells | Control | SARS-CoV-2-22 dpi | SARS-CoV-25 dpi | SARS-CoV-27 dpi | SARS-CoV-29 dpi |
|---|---|---|---|---|---|
| Erythroblast | - | 0.9 ± 0.1 | - | - | - |
| Lymphoblast | 0.3 ± 0.1 | 3.2 ± 0.5 * | 2.7 ± 0.5 * | 2.0 ± 0.3 | 3.0 ± 0.4 * |
| Lymphocyte | 61.8 ± 7.1 | 54.7 ± 6.3 | 44.0 ± 4.9 | 30.9 ± 4.2 * | 46.9 ± 5.1 |
| Limph.patol. | - | 1 ± 0.1 * | 1.0 ± 0.1 * | 1.0 ± 0.1 * | - |
| Monoblast | 0.4 ± 0.1 | 1 ± 0.1 | - | 1.0 ± 0.1 | 1.0 ± 0.1 |
| Monocyte | 3.9 ±0.4 | 2.8 ± 0.2 | 3.3 ± 0.6 | 4.0 ± 0.5 | 3.0 ± 0.2 |
| Mieloid | - | 3.0 ± 0.6 * | 0.01 ± 0.01 | 2.0 ± 0.4 * | 1.0 ± 0.3 |
| Metamielocyte | 1.4 ± 0.1 | 3.2 ± 0.2 | 0.1 ± 0.04 | 3.3 ± 0.5 | 3.0 ± 0.4 |
| Band | 17.5 ± 2.6 | 15.0 ± 2.7 | 30.3 ± 3.0 | 30.2 ± 2.8 * | 25.7 ± 2.2 |
| Segment | 13.3 ± 2.1 | 10.5 ± 1.8 | 15.3 ± 1.3 | 22.5 ± 1.9 * | 15.0 ± 1.5 |
| Pathol neutrophil | - | 2.0 ± 0.7 * | 0.01 ± 0.01 | 3.0 ± 0.9 * | 0.5 ± 0.7 |
| Eosinophil | 1.4 ± 0.2 | 1.5 ± 0.3 | 1.0 ± 0.1 | 0.1 ± 0.1 | 0.5 ± 0.2 |
| Basophil | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.3 ± 0.1 | 0.01 ± 0.01 | 0.01 ± 0.01 |
| Destroied | - | 1.0 ± 0.05 * | 2.0 ± 0.01 * | 0.01 ± 0.01 | - |
| Cellls | Control | SARS-CoV-2 9 dpi | SARS-CoV-2 18 dpi |
|---|---|---|---|
| Proerythroblast | 2.09 ± 0.5 | 2.27 ± 0.7 | 1.85 ± 0.4 |
| Basophil erythroblast | 9.97 ± 1.1 | 5.02 ± 0.8 * | 6.92 ± 1.0 |
| Polichromatophil erythroblast | 15.53 ± 2.4 | 6.01 ± 0.8 * | 10.25 ± 1.3 |
| Ortochrom erythroblast | 9.11 ± 0.9 | 6.36 ± 1.5 | 7.21 ± 1.1 |
| Lymphoblast | 1.07 ± 0.2 | 3.98 ± 0.7 * | 2.01 ± 0.5 |
| Lymphocyte | 3.31 ± 0.7 | 11.09 ± 1.1 * | 4.97 ± 0.7 |
| Limph.patol. | - | 0.45 ± 0.06 * | 0.29 ± 0.04 |
| Monoblast | 0.81 ± 0.1 | 3.09 ± 0.9 * | 0.88 ± 0.07 |
| Monocyte | 1.17 ± 0.3 | 4.57 ± 1.1 * | 2.36 ± 0.5 |
| Myeloid | 13.65 ± 2.9 | 4.18 ± 0.9 * | 5.76 ± 0.9 * |
| Metamielocyte | 25.62 ± 4.7 | 11.32 ± 2.5 * | 20.34 ± 4.4 |
| Band | 14.13 ± 2.1 | 32.64 ± 6.4 * | 27.55 ± 5.8 |
| Segment | 2.35 ± 0.6 | 4.18 ± 1.2 | 7.52 ± 2.0 * |
| Pathol neutrophil | - | 0.81 ± 0.1 * | 0.53 ± 0.06 * |
| Eosinophil | 0.91 ± 0.2 | 1.45 ± 0.09 | 1.02 ± 0.2 |
| Basophil | 0.08 ± 0.01 | 0.09 ± 0.01 | 0.02 ± 0.01 |
| Destroied | 0.01 ± 0.01 | 1.80 ± 0.3 * | 0.24 ± 0.02 |
| Macroph. Resident | 0.14 ± 0.05 | 0.24 ± 0.05 | 0.09 ± 0.01 |
| Macroph. Island | 0.02 ± 0.01 | 0.09 ± 0.01 * | 0.07 ± 0.01 |
| Megacarioblast | 0.01 ± 0.01 | 0.06 ± 0.01 | 0.07 ± 0.01 * |
| Megakaryocyte basophil | 0.01 ± 0.01 | 0.05 ± 0.01 | 0.02 ± 0.01 |
| Megakaryocyte azurophil | 0.01 ± 0.01 | 0.04 ± 0.01 | 0.02 ± 0.01 |
| Mitos | 0.01 ± 0.01 | 0.18 ± 0.01 * | 0.01 ± 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avagyan, H.; Hakobyan, S.; Poghosyan, A.; Hakobyan, L.; Abroyan, L.; Karalova, E.; Avetisyan, A.; Sargsyan, M.; Baghdasaryan, B.; Bayramyan, N.; et al. Severe Acute Respiratory Syndrome Coronavirus-2 Delta Variant Study In Vitro and Vivo. Curr. Issues Mol. Biol. 2023, 45, 249-267. https://doi.org/10.3390/cimb45010019
Avagyan H, Hakobyan S, Poghosyan A, Hakobyan L, Abroyan L, Karalova E, Avetisyan A, Sargsyan M, Baghdasaryan B, Bayramyan N, et al. Severe Acute Respiratory Syndrome Coronavirus-2 Delta Variant Study In Vitro and Vivo. Current Issues in Molecular Biology. 2023; 45(1):249-267. https://doi.org/10.3390/cimb45010019
Chicago/Turabian StyleAvagyan, Hranush, Sona Hakobyan, Arpine Poghosyan, Lina Hakobyan, Liana Abroyan, Elena Karalova, Aida Avetisyan, Mariam Sargsyan, Bagrat Baghdasaryan, Nane Bayramyan, and et al. 2023. "Severe Acute Respiratory Syndrome Coronavirus-2 Delta Variant Study In Vitro and Vivo" Current Issues in Molecular Biology 45, no. 1: 249-267. https://doi.org/10.3390/cimb45010019