
API-2-Induced Cell Migration Is Overcome by Small Molecular Approaches Inhibiting β-Catenin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Drug Treatment
2.2. Gene-Expression Data Analysis
2.3. Immunocytochemistry and Phalloidin Staining
2.4. Wound-Healing Assay
2.5. Flow Cytometry Analysis
2.6. Statistical Analyses
3. Results
3.1. High Expression of CTNNB1 and FOXO3a Is Associated with Poor Survival in CRC Patients
3.2. KY1022 Effectively Destabilizes β-Catenin in the Presence of API-2
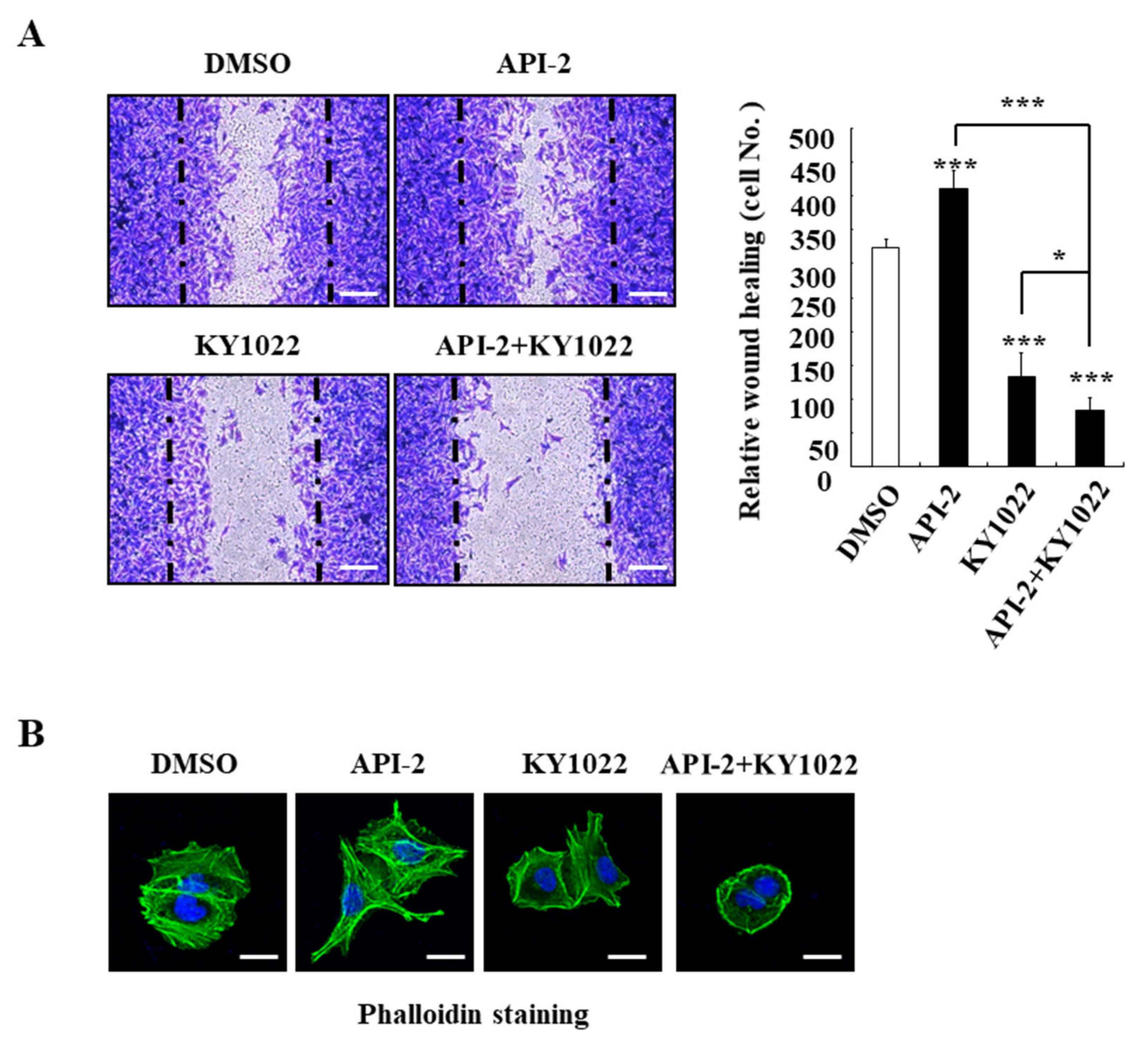
3.3. KY1022 Reverts the AKT-Inhibitor-Induced Cell Motility and Suppresses the Actin Rearrangement of CRC Cells
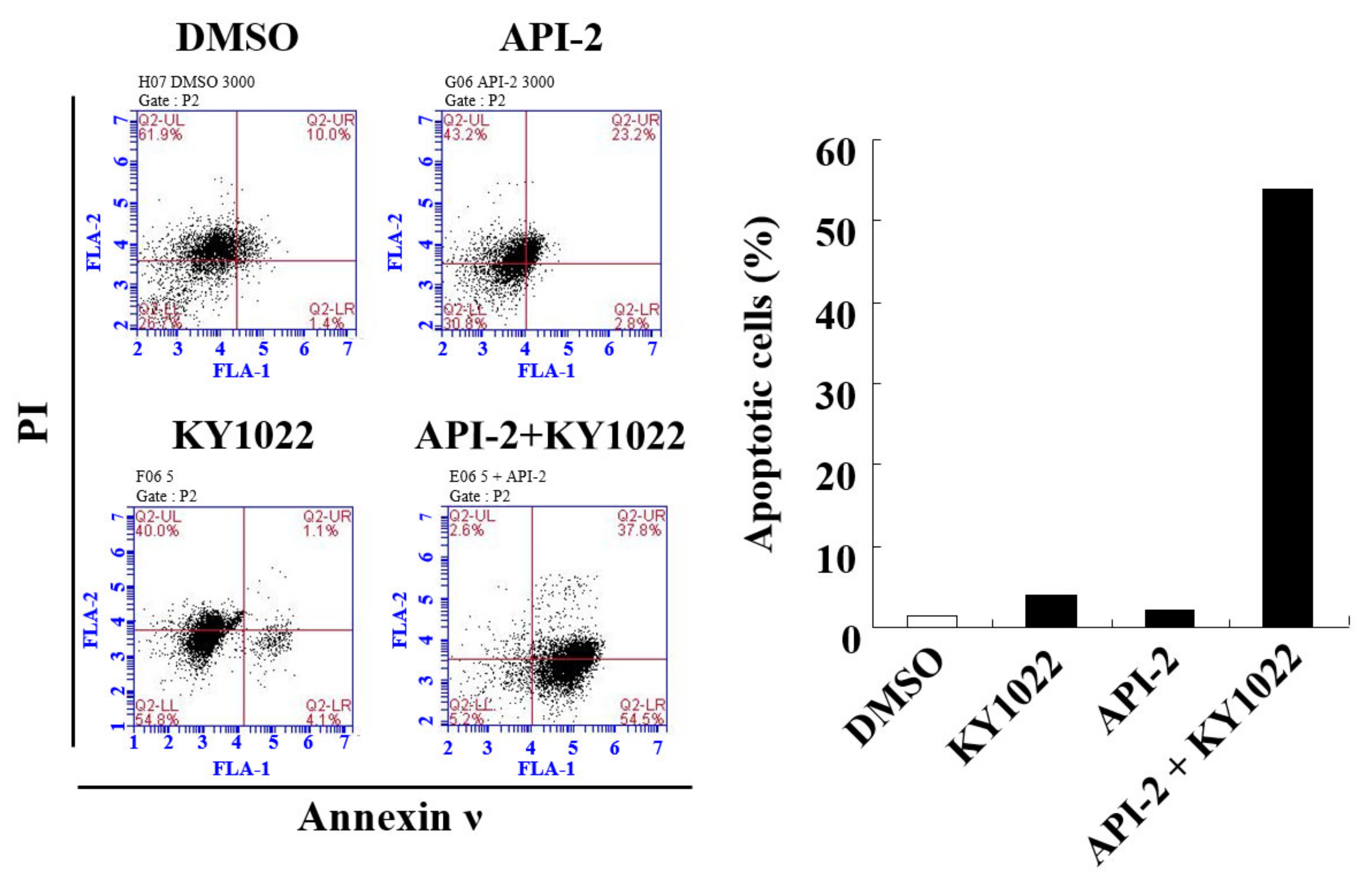
3.4. KY1022 Overcomes the AKT-Inhibitor-Induced Resistance of Apoptosis in CRC Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Vatandoust, S.; Price, T.J.; Karapetis, C.S. Colorectal cancer: Metastases to a single organ. World J. Gastroenterol. 2015, 21, 11767–11776. [Google Scholar] [CrossRef]
- Ye, Q.; Cai, W.; Zheng, Y.; Evers, B.M.; She, Q.B. ERK and AKT signaling cooperate to translationally regulate survivin expression for metastatic progression of colorectal cancer. Oncogene 2014, 33, 1828–1839. [Google Scholar] [CrossRef] [Green Version]
- Fevr, T.; Robine, S.; Louvard, D.; Huelsken, J. Wnt/beta-catenin is essential for intestinal homeostasis and maintenance of intestinal stem cells. Mol. Cell Biol. 2007, 27, 7551–7559. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Downward, J. Targeting RAS signalling pathways in cancer therapy. Nat. Rev. Cancer 2003, 3, 11–22. [Google Scholar] [CrossRef]
- Kang, D.W.; Lee, B.H.; Suh, Y.A.; Choi, Y.S.; Jang, S.J.; Kim, Y.M.; Choi, K.Y.; Min, D.S. Phospholipase D1 Inhibition Linked to Upregulation of ICAT Blocks Colorectal Cancer Growth Hyperactivated by Wnt/beta-Catenin and PI3K/Akt Signaling. Clin. Cancer Res. 2017, 23, 7340–7350. [Google Scholar] [CrossRef] [Green Version]
- Koveitypour, Z.; Panahi, F.; Vakilian, M.; Peymani, M.; Seyed Forootan, F.; Nasr Esfahani, M.H.; Ghaedi, K. Signaling pathways involved in colorectal cancer progression. Cell Biosci. 2019, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- Qiao, M.; Sheng, S.; Pardee, A.B. Metastasis and AKT activation. Cell Cycle 2008, 7, 2991–2996. [Google Scholar] [CrossRef]
- Cao, Z.; Liao, Q.; Su, M.; Huang, K.; Jin, J.; Cao, D. AKT and ERK dual inhibitors: The way forward? Cancer Lett. 2019, 459, 30–40. [Google Scholar] [CrossRef]
- Dienstmann, R.; Rodon, J.; Markman, B.; Tabernero, J. Recent developments in anti-cancer agents targeting PI3K, Akt and mTORC1/2. Recent Pat. Anti Cancer Drug Discov. 2011, 6, 210–236. [Google Scholar] [CrossRef]
- Hua, H.; Zhang, H.; Chen, J.; Wang, J.; Liu, J.; Jiang, Y. Targeting Akt in cancer for precision therapy. J. Hematol. Oncol. 2021, 14, 128. [Google Scholar] [CrossRef]
- Yang, J.-Y.; Hung, M.-C. Deciphering the role of forkhead transcription factors in cancer therapy. Curr. Drug Targets 2011, 12, 1284–1290. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Qi, P.; Hu, X. Downregulated FOXO3a Associates With Poor Prognosis and Promotes Cell Invasion and Migration via WNT/β-catenin Signaling in Cervical Carcinoma. Front. Oncol. 2020, 10, 903. [Google Scholar] [CrossRef]
- Laissue, P. The forkhead-box family of transcription factors: Key molecular players in colorectal cancer pathogenesis. Mol. Cancer 2019, 18, 5. [Google Scholar] [CrossRef] [Green Version]
- Tenbaum, S.P.; Ordóñez-Morán, P.; Puig, I.; Chicote, I.; Arqués, O.; Landolfi, S.; Fernández, Y.; Herance, J.R.; Gispert, J.D.; Mendizabal, L.; et al. β-catenin confers resistance to PI3K and AKT inhibitors and subverts FOXO3a to promote metastasis in colon cancer. Nat. Med. 2012, 18, 892–901. [Google Scholar] [CrossRef] [Green Version]
- Ordóñez-Morán, P.; Irmisch, A.; Barbáchano, A.; Chicote, I.; Tenbaum, S.; Landolfi, S.; Tabernero, J.; Huelsken, J.; Muñoz, A.; Pálmer, H.G. SPROUTY2 is a β-catenin and FOXO3a target gene indicative of poor prognosis in colon cancer. Oncogene 2014, 33, 1975–1985. [Google Scholar] [CrossRef] [Green Version]
- Arques, O.; Chicote, I.; Puig, I.; Tenbaum, S.P.; Argiles, G.; Dienstmann, R.; Fernandez, N.; Caratu, G.; Matito, J.; Silberschmidt, D.; et al. Tankyrase Inhibition Blocks Wnt/beta-Catenin Pathway and Reverts Resistance to PI3K and AKT Inhibitors in the Treatment of Colorectal Cancer. Clin. Cancer Res. 2016, 22, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Cha, P.H.; Cho, Y.H.; Lee, S.K.; Lee, J.; Jeong, W.J.; Moon, B.S.; Yun, J.H.; Yang, J.S.; Choi, S.; Yoon, J.; et al. Small-molecule binding of the axin RGS domain promotes beta-catenin and Ras degradation. Nat. Chem. Biol. 2016, 12, 593–600. [Google Scholar] [CrossRef]
- Cho, Y.H.; Cha, P.H.; Kaduwal, S.; Park, J.C.; Lee, S.K.; Yoon, J.S.; Shin, W.; Kim, H.; Ro, E.J.; Koo, K.H.; et al. KY1022, a small molecule destabilizing Ras via targeting the Wnt/β-catenin pathway, inhibits development of metastatic colorectal cancer. Oncotarget 2016, 7, 81727–81740. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.H.; Choi, H.; Oshima, M.; Cheong, J.H.; Kim, S.; Lee, J.H.; Park, Y.S.; Choi, H.S.; Kweon, M.N.; Pack, C.G.; et al. Estrogen-related receptor gamma functions as a tumor suppressor in gastric cancer. Nat. Commun. 2018, 9, 1920. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.H.; Ro, E.J.; Yoon, J.S.; Kwak, D.K.; Cho, J.; Kang, D.W.; Lee, H.Y.; Choi, K.Y. Small molecule-induced simultaneous destabilization of beta-catenin and RAS is an effective molecular strategy to suppress stemness of colorectal cancer cells. Cell Commun. Signal. 2020, 18, 38. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, S.; Shui, X.; Craig, K.P.; Koser, M.L.; Chopda, G.R.; Cyr, W.A.; Lai, C.; Dudek, H.; Wang, W.; Brown, B.D.; et al. β-Catenin mRNA Silencing and MEK Inhibition Display Synergistic Efficacy in Preclinical Tumor Models. Mol. Cancer Ther. 2018, 17, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Jeong, W.J.; Ro, E.J.; Choi, K.Y. Interaction between Wnt/beta-catenin and RAS-ERK pathways and an anti-cancer strategy via degradations of beta-catenin and RAS by targeting the Wnt/beta-catenin pathway. NPJ Precis. Oncol. 2018, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin. Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Yang, J.; Nie, J.; Ma, X.; Wei, Y.; Peng, Y.; Wei, X. Targeting PI3K in cancer: Mechanisms and advances in clinical trials. Mol. Cancer 2019, 18, 26. [Google Scholar] [CrossRef] [Green Version]
- Fodde, R. The APC gene in colorectal cancer. Eur. J. Cancer 2002, 38, 867–871. [Google Scholar] [CrossRef]
- Polakis, P. The oncogenic activation of beta-catenin. Curr. Opin. Genet. Dev. 1999, 9, 15–21. [Google Scholar] [CrossRef]
- Reymond, N.; d’Agua, B.B.; Ridley, A.J. Crossing the endothelial barrier during metastasis. Nat. Rev. Cancer 2013, 13, 858–870. [Google Scholar] [CrossRef]
- Hapach, L.A.; Mosier, J.A.; Wang, W.; Reinhart-King, C.A. Engineered models to parse apart the metastatic cascade. NPJ Precis Oncol. 2019, 3, 20. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Kang, M.-H.; Cho, Y.-H. API-2-Induced Cell Migration Is Overcome by Small Molecular Approaches Inhibiting β-Catenin. Curr. Issues Mol. Biol. 2022, 44, 6006-6014. https://doi.org/10.3390/cimb44120409
Kim Y, Kang M-H, Cho Y-H. API-2-Induced Cell Migration Is Overcome by Small Molecular Approaches Inhibiting β-Catenin. Current Issues in Molecular Biology. 2022; 44(12):6006-6014. https://doi.org/10.3390/cimb44120409
Chicago/Turabian StyleKim, Yonghyo, Myoung-Hee Kang, and Yong-Hee Cho. 2022. "API-2-Induced Cell Migration Is Overcome by Small Molecular Approaches Inhibiting β-Catenin" Current Issues in Molecular Biology 44, no. 12: 6006-6014. https://doi.org/10.3390/cimb44120409