
The Nocardia Rubra Cell Wall Skeleton Regulates Macrophages and Promotes Wound Healing

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. In Vivo Experiments
2.2.1. Experimental Healing of Full-Thickness Excisional Wounds in Mice
2.2.2. Histopathological Examination
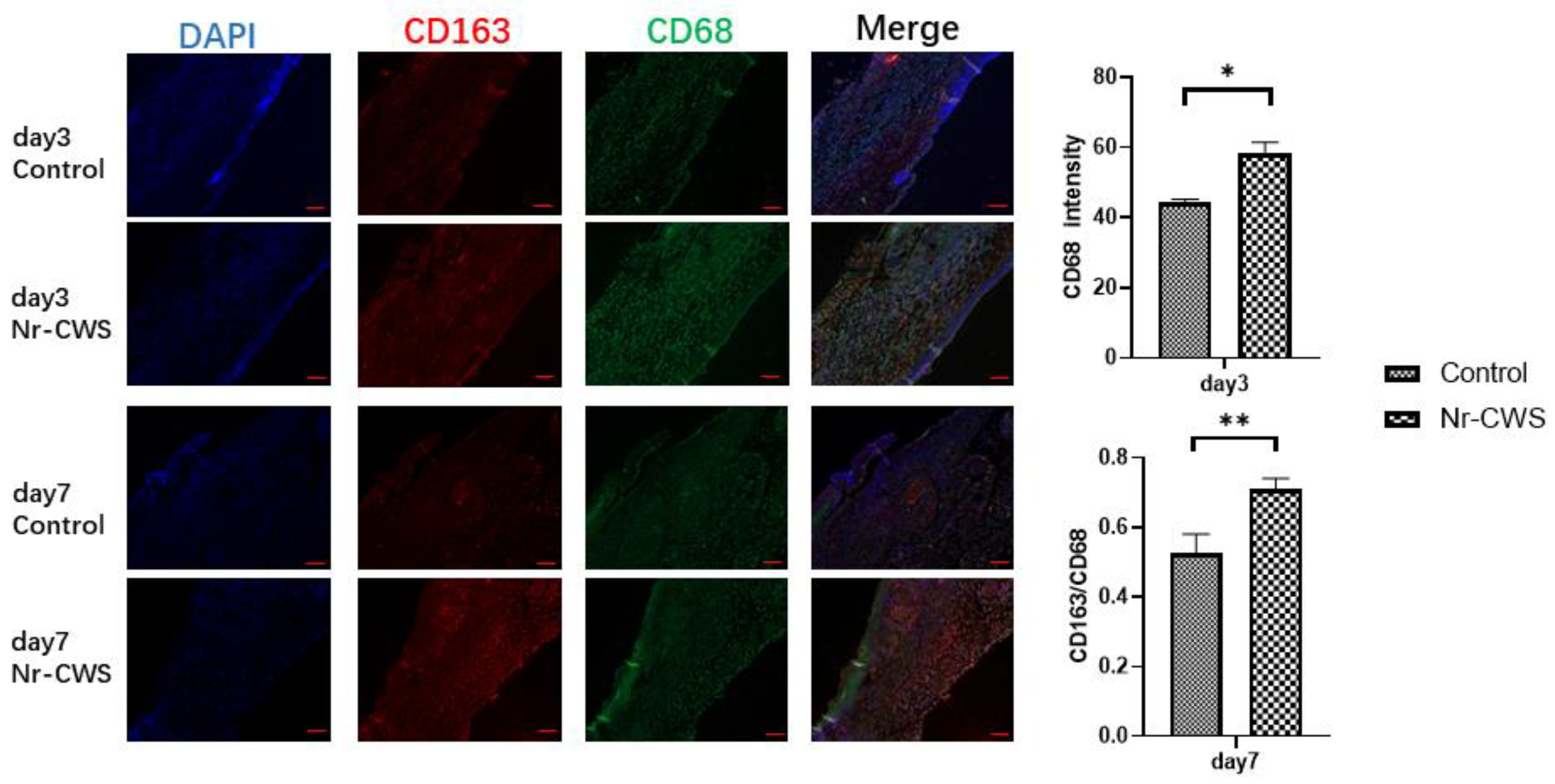
2.2.3. Immunofluorescence
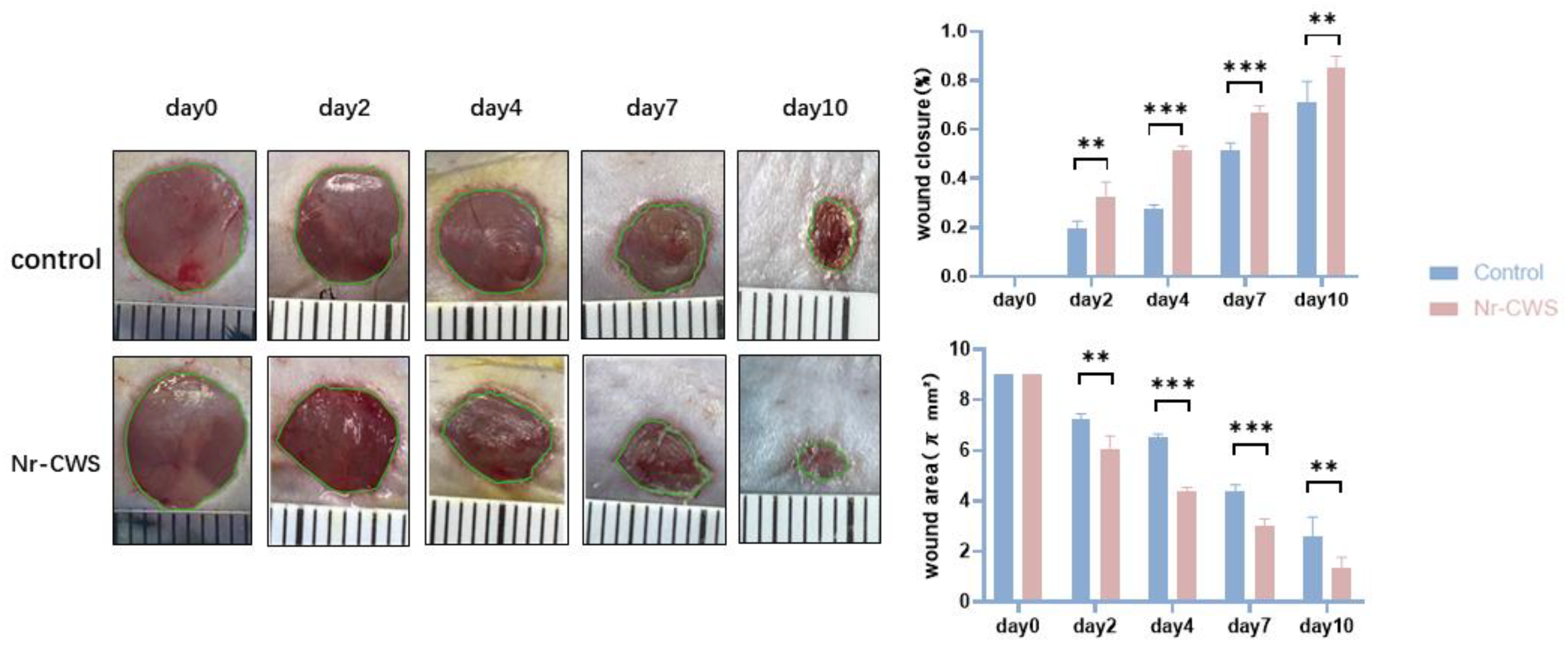
2.2.4. Wound Healing Rate
2.3. In Vitro Experiments
2.3.1. Cell Culture
2.3.2. Cell Viability Assay
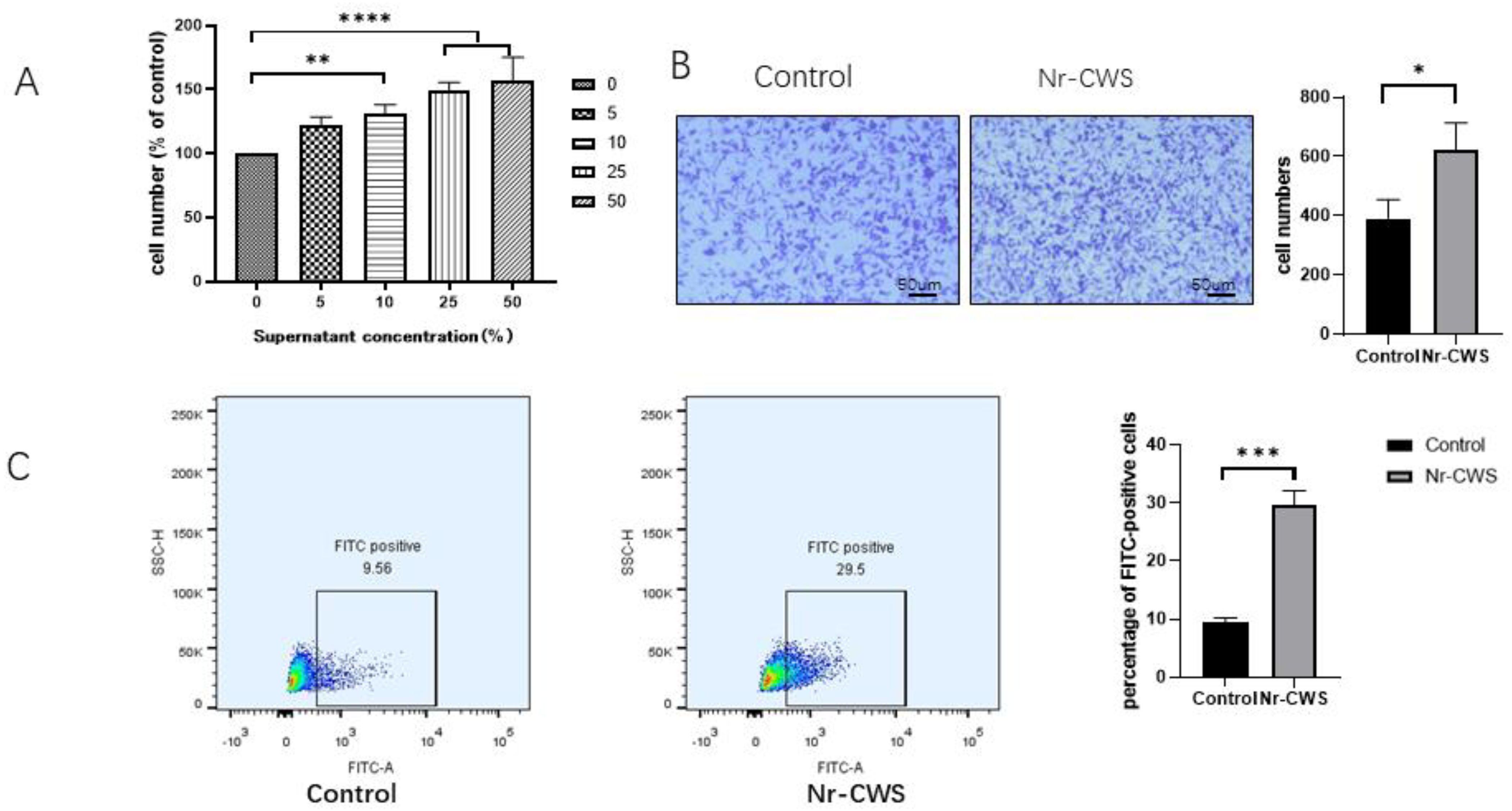
2.3.3. Cell Phagocytosis
2.3.4. Cell Migration
2.3.5. Cell Immunofluorescence
2.3.6. Flow Cytometry
2.3.7. RNA Extraction and Quantitative Real-Time PCR (qPCR)
2.3.8. ELISA
2.3.9. Western Blot Analysis
2.4. Statistical Analysis
3. Results
3.1. In Vitro Experiments
3.1.1. Nr-CWS Enhances the Proliferation, Migration, and Phagocytosis of RAW264.7 Macrophages
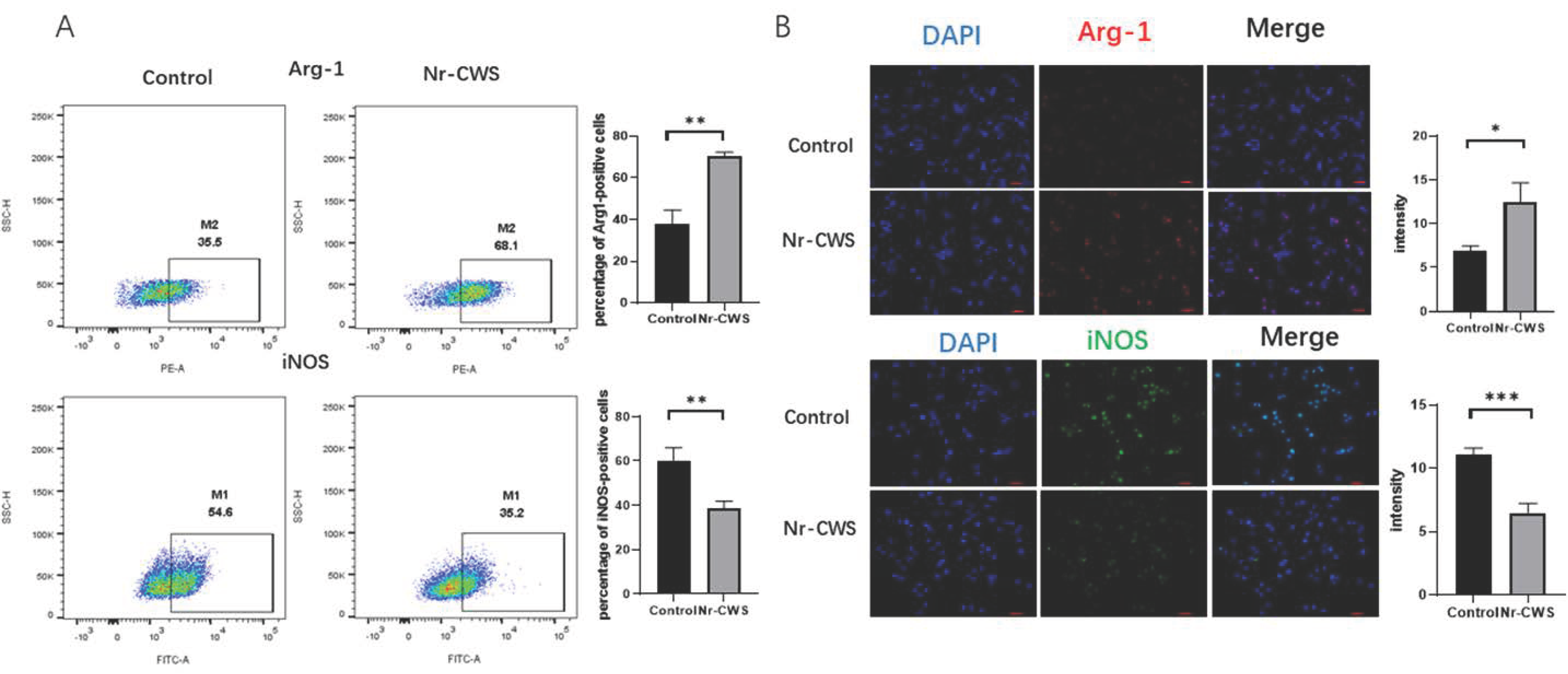
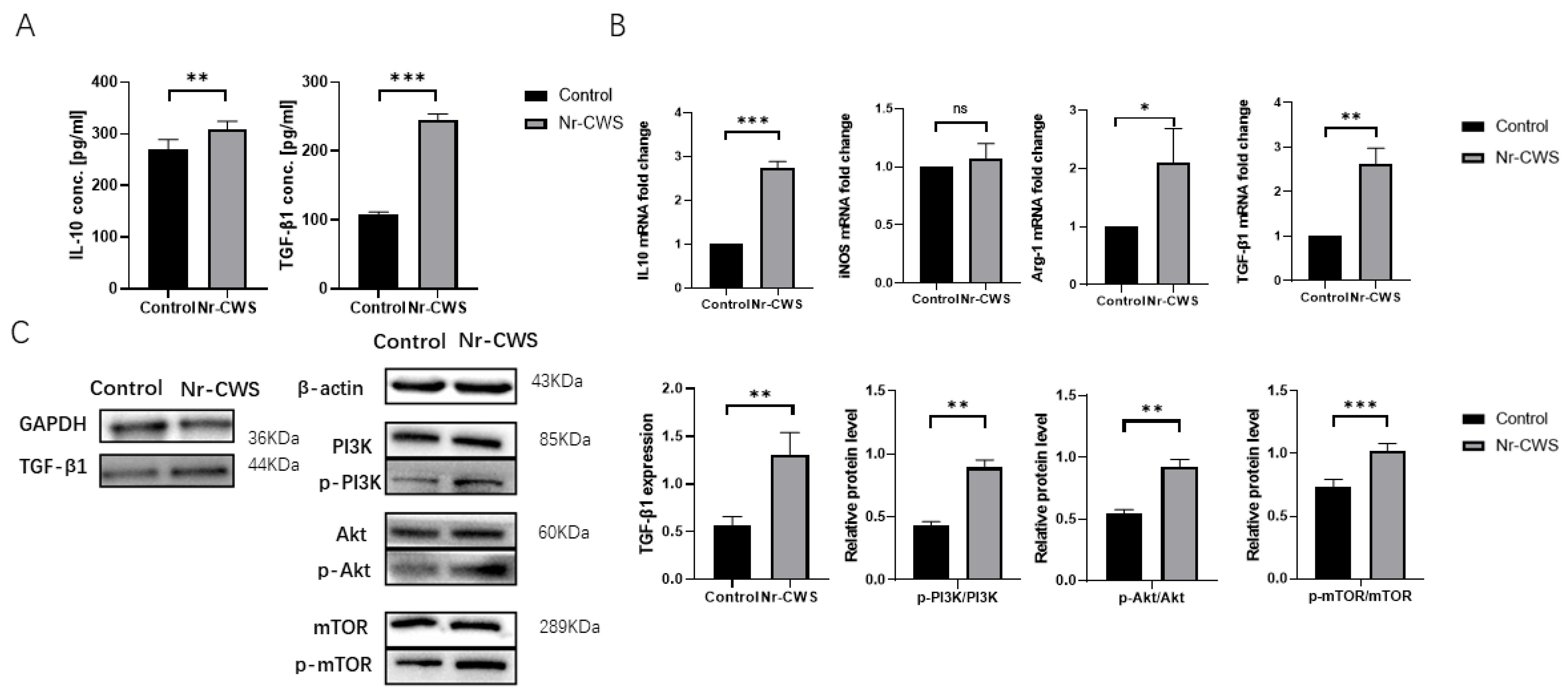
3.1.2. Nr-CWS Can Polarize Macrophages toward the M2 Type and Promote the Expression of Related Cytokines
3.2. In Vivo Experiments
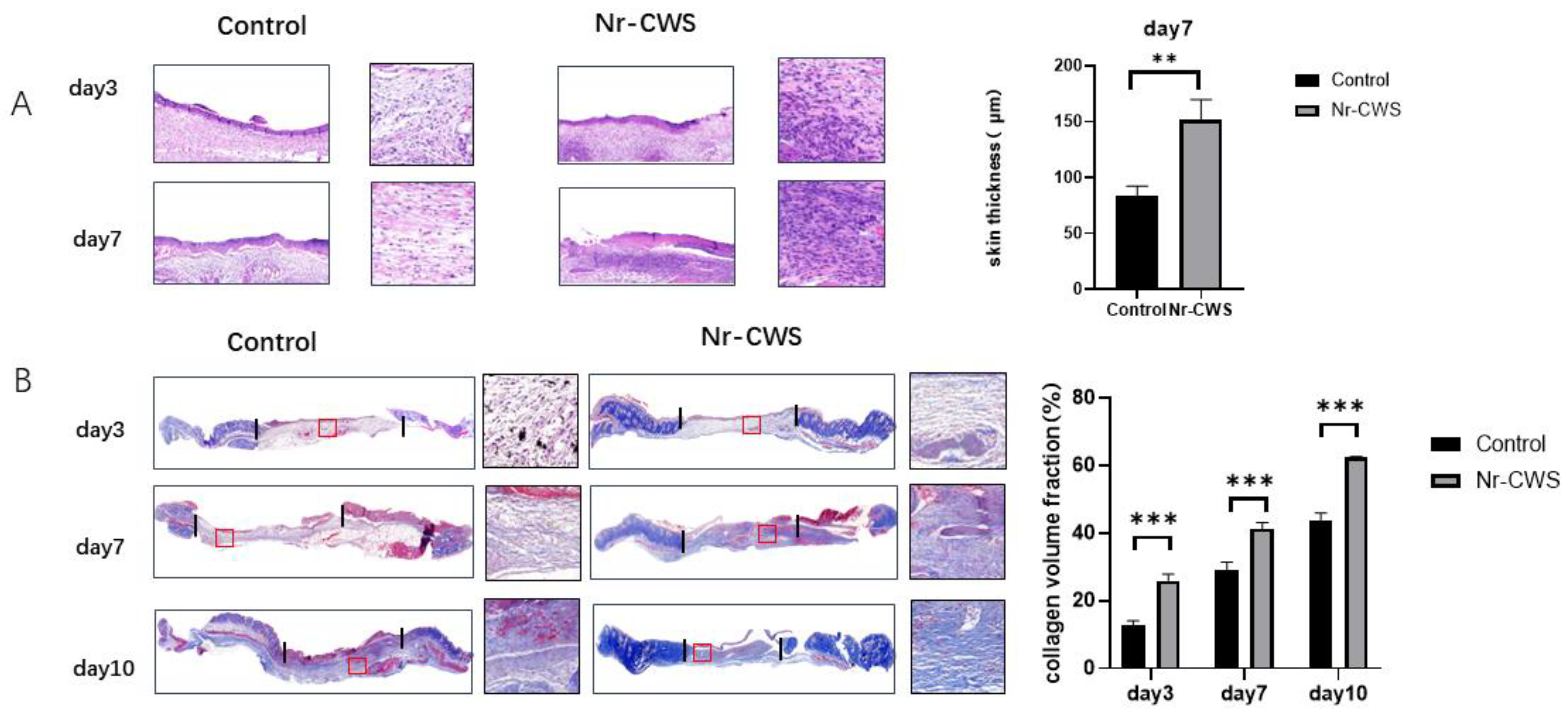
3.2.1. Nr-CWS Can Enhance the Recruitment of Macrophages in the Wound and Polarize the Macrophages
3.2.2. Nr-CWS Can Promote Wound Healing
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.; Wang, H.; Liu, Y.; Tao, T.; Zeng, Z.; Zhou, Y.; Wang, M. Nocardia rubra cell-wall skeleton influences the development of cervical carcinoma by promoting the antitumor effect of macrophages and dendritic cells. Cancer Med. 2022, 11, 1249–1268. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, Y.; Ma, B.; Wang, F.; Liu, S.; Xu, J.; Chen, X.-L.; Lü, X.-W. Nocardia rubra cell wall skeleton accelerates cutaneous wound healing by enhancing macro phage activation and angiogenesis. J. Int. Med. Res. 2018, 46, 2398–2409. [Google Scholar] [CrossRef]
- Heydari, P.; Kharaziha, M.; Varshosaz, J.; Javanmard, S.H. Current knowledge of immunomodulation strategies for chronic skin wound repair. J. Biomed. Mater. Res. Part B Appl. Biomater. 2022, 110, 265–288. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Huang, J.; Lin, M.; Xie, T.; You, T. Quercetin Promotes Diabetic Wound Healing via Switching Macrophages From M1 to M2 Polarization. J. Surg. Res. 2020, 246, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.; Jang, H.; Lee, Y.; Moon, Y.J.; Bae, E.J.; Yun, S.; Park, B. Myeloid cell-specific sirtuin 6 deficiency delays wound healing in mice by modulating inflammation and macrophage phenotypes. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- Chen, L.; Xu, Y.; Zhao, J.; Zhang, Z.; Yang, R.; Xie, J.; Liu, X.; Qi, S. Correction: Conditioned Medium from Hypoxic Bone Marrow-Derived Mesenchymal Stem Cells Enhances Wound Healing in Mice. PLoS ONE 2015, 10, e145565. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Huang, P.; Yuan, B.; Liu, T.; Lan, F.; Lu, X.; Dai, L.; Liu, Y.; Yin, H. Astragaloside IV enhances diabetic wound healing involving upregulation of alternatively activated macrophages. Int. Immunopharmacol. 2016, 35, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ma, Z.; Kong, L.; He, Y.; Chan, H.F.; Li, H. Modulation of macrophages by bioactive glass/sodium alginate hydrogel is crucial in skin regeneration enhancement. Biomaterials 2020, 256, 120216. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Nie, S.; Li, W.; Zheng, W.; Yin, P.; Gong, D.; Xie, M. Macrophage Immunomodulatory Activity of a Purified Polysaccharide Isolated fromGanoderma atrum. Phytother. Res. 2013, 27, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Hu, S.; Cui, Z.; Li, W. Probiotic Bacillus amyloliquefaciens mediate M1 macrophage polarization in mouse bone marrow-derived macrophages. Arch. Microbiol. 2013, 195, 349–356. [Google Scholar] [CrossRef]
- Zou, Y.; Zhang, J.; Xu, J.; Fu, L.; Xu, Y.; Wang, X.; Li, Z.; Zhu, L.; Sun, H.; Zheng, H.; et al. SIRT6 inhibition delays peripheral nerve recovery by suppressing migration, phagocytosis and M2-NE.Refpolarization of macrophages. Cell Biosci. 2021, 11, 210. [Google Scholar] [CrossRef]
- Rocher, C.; Singla, D.K. SMAD-PI3K-Akt-mTOR pathway mediates BMP-7 polarization of monocytes into M2 macrophages. PLoS ONE 2013, 8, e84009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, G.; Chen, S.; Ouyang, M.; Li, F.; Chen, L.; Yang, J. Colon Cancer Cell Secretes EGF to Promote M2 Polarization of TAM Through EGFR/PI3K/AKT/mTOR Pathway. Technol. Cancer Res. Treat. 2019, 18, 1078116716. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, A.; Fan, L. HSPA12B Secreted by Tumor-Associated Endothelial Cells Might Induce M2 Polarization of Macrophages via Activating PI3K/Akt/mTOR Signaling. Onco Targets Ther. 2020, 13, 9103–9111. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Sun, J.; Wang, X.; Ma, Y.; Kong, C.; Zhang, G.; Dou, H.; Nan, N.; Shi, M.; Yu, T.; et al. The biological macromolecule Nocardia rubra cell-wall skeleton as an avenue for cell-based immunotherapy. J. Cell. Physiol. 2019, 234, 15342–15356. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Snyder, R.J.; Lantis, J.; Kirsner, R.S.; Shah, V.; Molyneaux, M.; Carter, M.J. Macrophages: A review of their role in wound healing and their therapeutic use. Wound Repair Regen. 2016, 24, 613–629. [Google Scholar] [CrossRef]
- Vannella, K.M.; Wynn, T.A. Mechanisms of Organ Injury and Repair by Macrophages. Annu. Rev. Physiol. 2017, 79, 593–617. [Google Scholar] [CrossRef]
- Goren, I.; Allmann, N.; Yogev, N.; Schürmann, C.; Linke, A.; Holdener, M.; Waisman, A.; Pfeilschifter, J.; Frank, S. A Transgenic Mouse Model of Inducible Macrophage Depletion. Am. J. Pathol. 2009, 175, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Mirza, R.; DiPietro, L.A.; Koh, T.J. Selective and Specific Macrophage Ablation Is Detrimental to Wound Healing in Mice. Am. J. Pathol. 2009, 175, 2454–2462. [Google Scholar] [CrossRef]
- Nassar, D.; Letavernier, E.; Baud, L.; Aractingi, S.; Khosrotehrani, K. Calpain activity is essential in skin wound healing and contributes to scar formation. PLoS ONE 2012, 7, e37084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boniakowski, A.E.; Kimball, A.S.; Joshi, A.; Schaller, M.; Davis, F.M.; DenDekker, A.; Obi, A.T.; Moore, B.B.; Kunkel, S.L.; Gallagher, K.A. Murine macrophage chemokine receptor CCR2 plays a crucial role in macrophage recruitment and regulated inflammation in wound healing. Eur. J. Immunol. 2018, 48, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Chazaud, B. Inflammation and Skeletal Muscle Regeneration: Leave It to the Macrophages! Trends Immunol. 2020, 41, 481–492. [Google Scholar] [CrossRef]
- Lee, J.; Rodero, M.P.; Patel, J.; Moi, D.; Mazzieri, R.; Khosrotehrani, K. Interleukin-23 regulates interleukin-17 expression in wounds, and its inhibition accelerates diabetic wound healing through the alteration of macrophage polarization. FASEB J. 2018, 32, 2086–2094. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Chen, L.; Song, M.; He, J.; Zhang, L.; Li, C.; Wang, Q.; Yang, W.; Sun, W.; Leng, Y.; et al. Naoxintong accelerates diabetic wound healing by attenuating inflammatory response. Pharm. Biol. 2021, 59, 250–259. [Google Scholar] [CrossRef]
- Sindrilaru, A.; Peters, T.; Wieschalka, S.; Baican, C.; Baican, A.; Peter, H.; Hainzl, A.; Schatz, S.; Qi, Y.; Schlecht, A.; et al. An unrestrained proinflammatory M1 macrophage population induced by iron impairs wound healing in humans and mice. J. Clin. Investig. 2011, 121, 985–997. [Google Scholar] [CrossRef] [Green Version]
- Kane, A.E.; Sinclair, D.A. Epigenetic changes during aging and their reprogramming potential. Crit. Rev. Biochem. Mol. Biol. 2019, 54, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.; Jayaraman, V.; Huelsmann, E.J.; Bonish, B.; Burgad, D.; Sivaramakrishnan, G.; Qin, S.; DiPietro, L.A.; Zloza, A.; Zhang, C.; et al. Pro-inflammatory chemokine CCL2 (MCP-1) promotes healing in diabetic wounds by restoring the macrophage response. PLoS ONE 2014, 9, e91574. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences (5′-3′) | Product Size (bp) |
|---|---|---|
| iNOS | forward:ATCTTGGAGCGAGTTGTGGATTGTC | 146 |
| reverse:TAGGTGAGGGCTTGGCTGAGTG | ||
| Arg-1 | forward:AGACAGCAGAGGAGGTGAAGAGTAC | 118 |
| reverse:AAGGTAGTCAGTCCCTGGCTTATGG | ||
| IL-10 | forward:AGAGAAGCATGGCCCAGAAATCAAG | 136 |
| reverse:CTTCACCTGCTCCACTGCCTTG | ||
| TGF-β1 | forward:ACCGCAACAACGCCATCTATGAG | 91 |
| reverse:GGCACTGCTTCCCGAATGTCTG | ||
| GAPDH | forward:TGACATCAAGAAGGTGGTGAAGCAG | 224 |
| reverse:GTGTCGCTGTTGAAGTCAGAGGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, K.; Xu, Y.; Li, X.; Du, P.; Lu, Y.; Lyu, G. The Nocardia Rubra Cell Wall Skeleton Regulates Macrophages and Promotes Wound Healing. Curr. Issues Mol. Biol. 2022, 44, 5995-6005. https://doi.org/10.3390/cimb44120408
Hu K, Xu Y, Li X, Du P, Lu Y, Lyu G. The Nocardia Rubra Cell Wall Skeleton Regulates Macrophages and Promotes Wound Healing. Current Issues in Molecular Biology. 2022; 44(12):5995-6005. https://doi.org/10.3390/cimb44120408
Chicago/Turabian StyleHu, Kai, Yan Xu, Xiaoxiao Li, Pan Du, Yichi Lu, and Guozhong Lyu. 2022. "The Nocardia Rubra Cell Wall Skeleton Regulates Macrophages and Promotes Wound Healing" Current Issues in Molecular Biology 44, no. 12: 5995-6005. https://doi.org/10.3390/cimb44120408