
High-altitude Hypoxia Influences the Activities of the Drug-Metabolizing Enzyme CYP3A1 and the Pharmacokinetics of Four Cardiovascular System Drugs

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. UHPLC-MS Method Validation
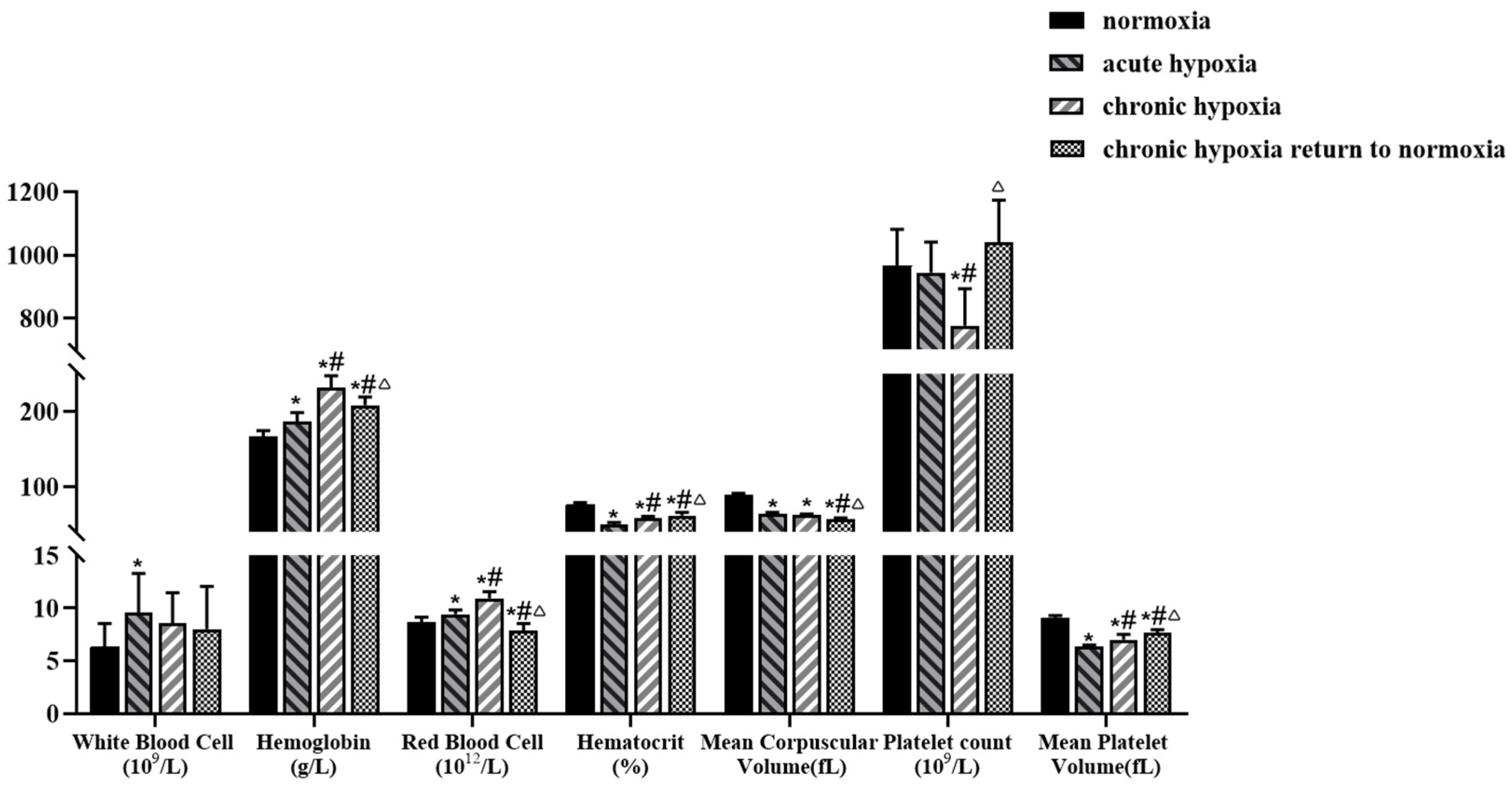
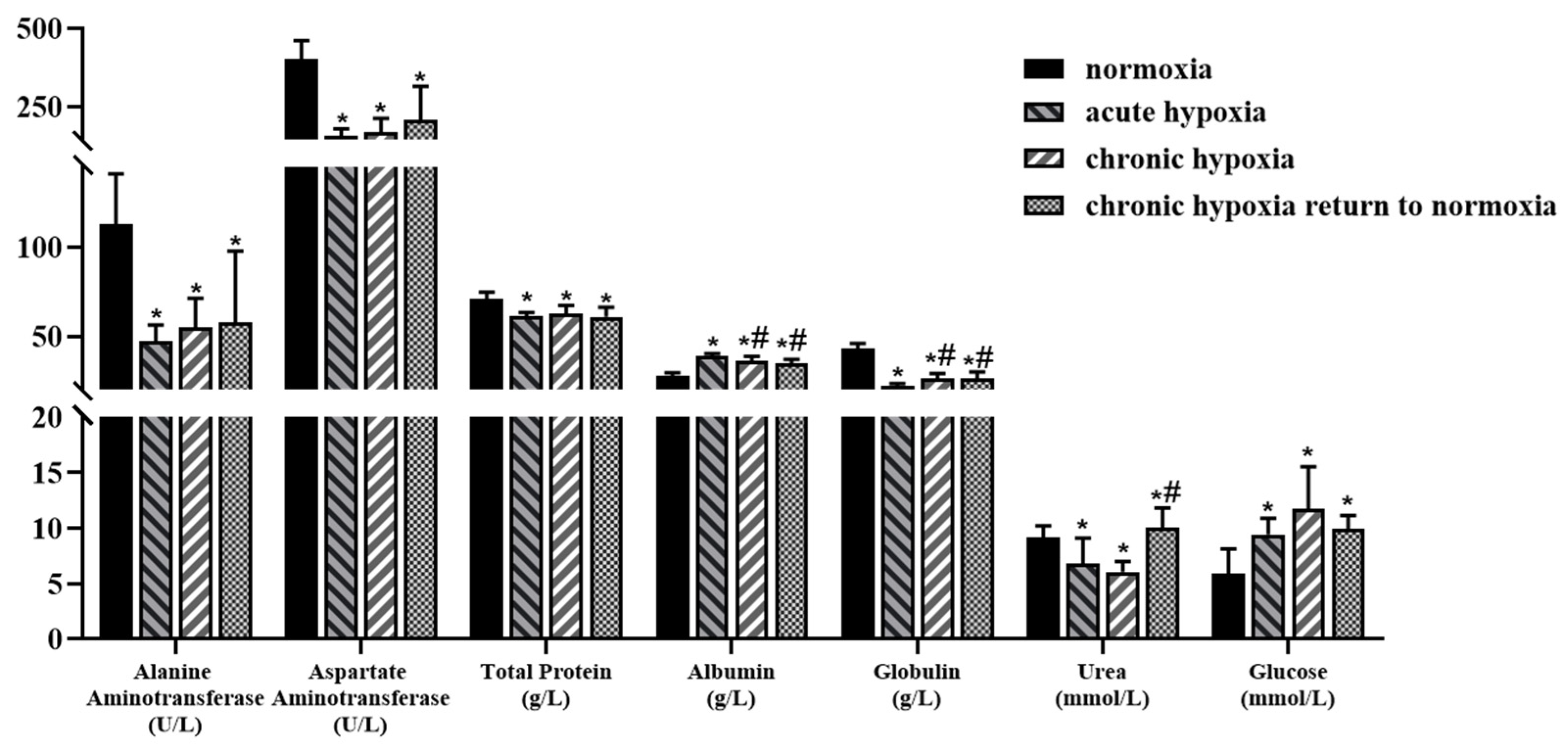
2.2. Physiologic and Biochemical Parameters
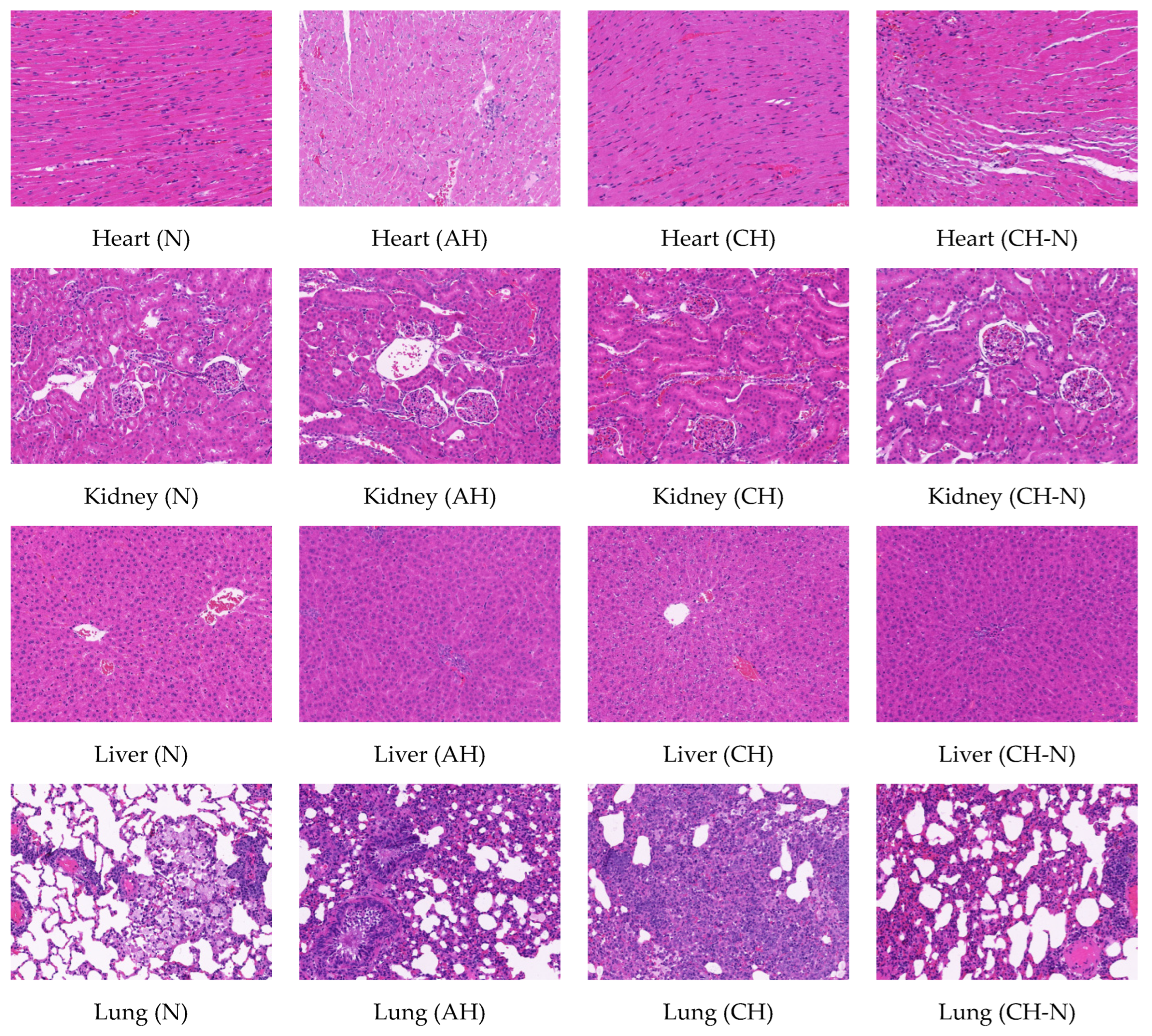
2.3. Hematoxylin-Eosin Staining of Heart, Kidney, Liver, and Lung Tissues in Rats
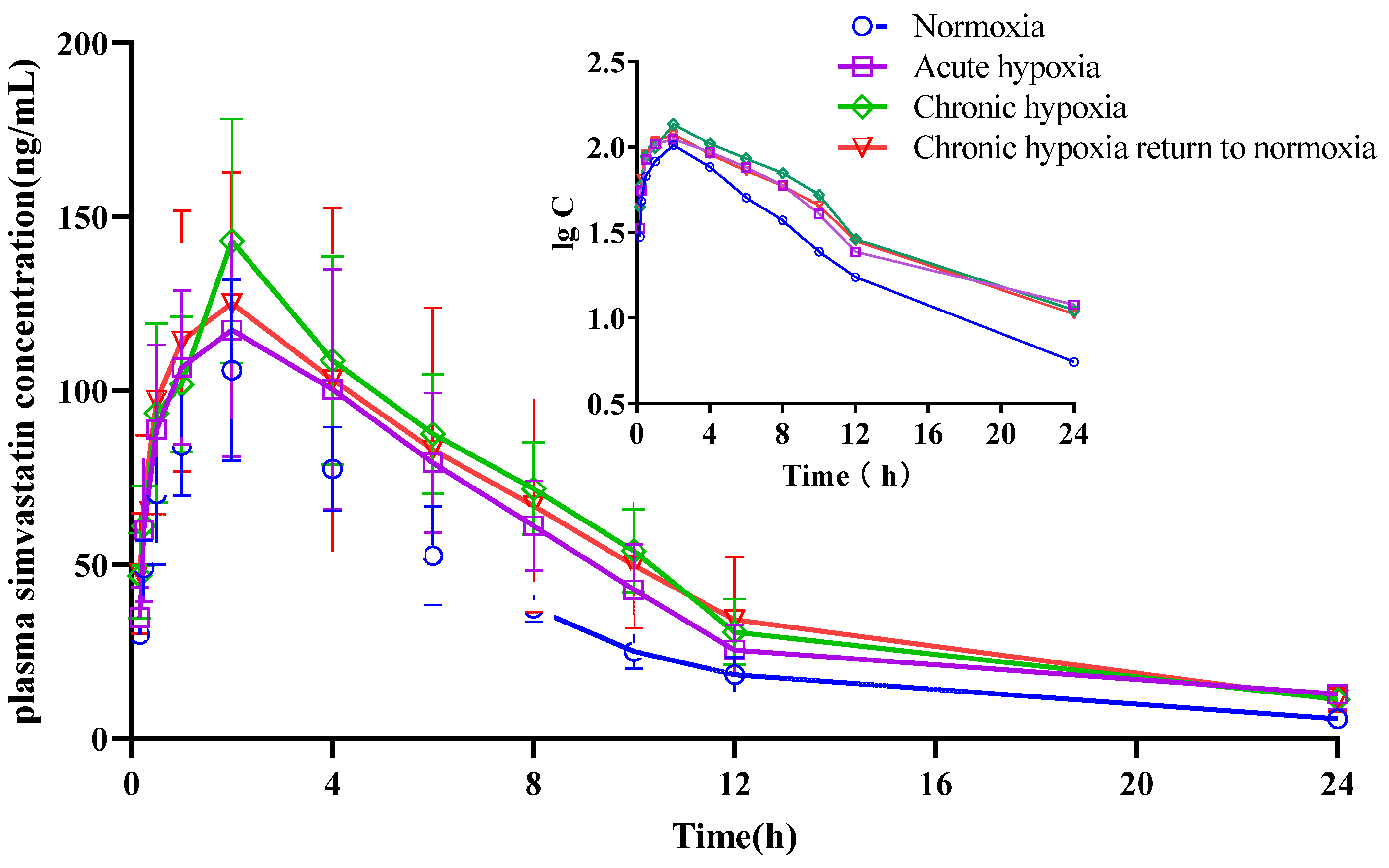
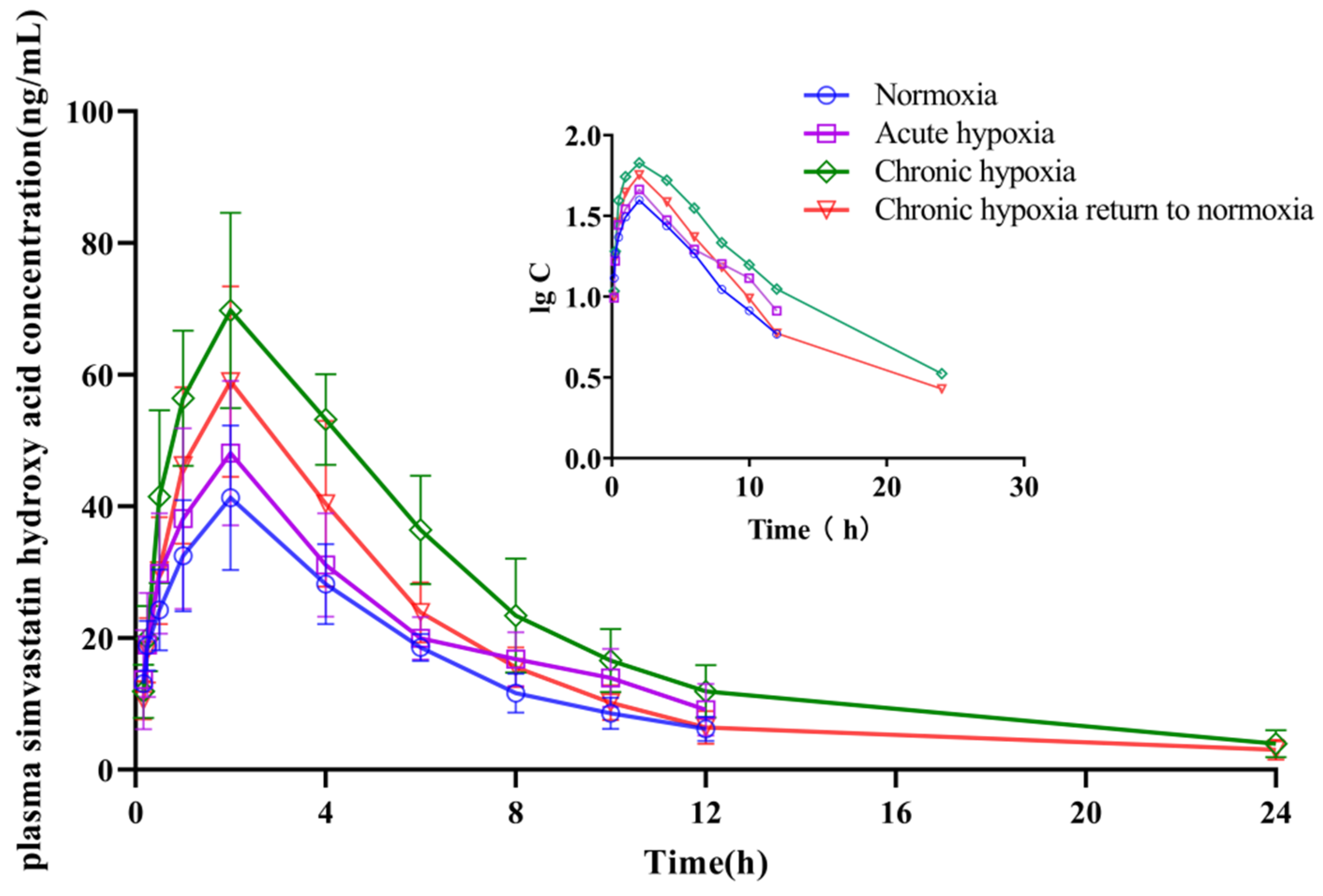
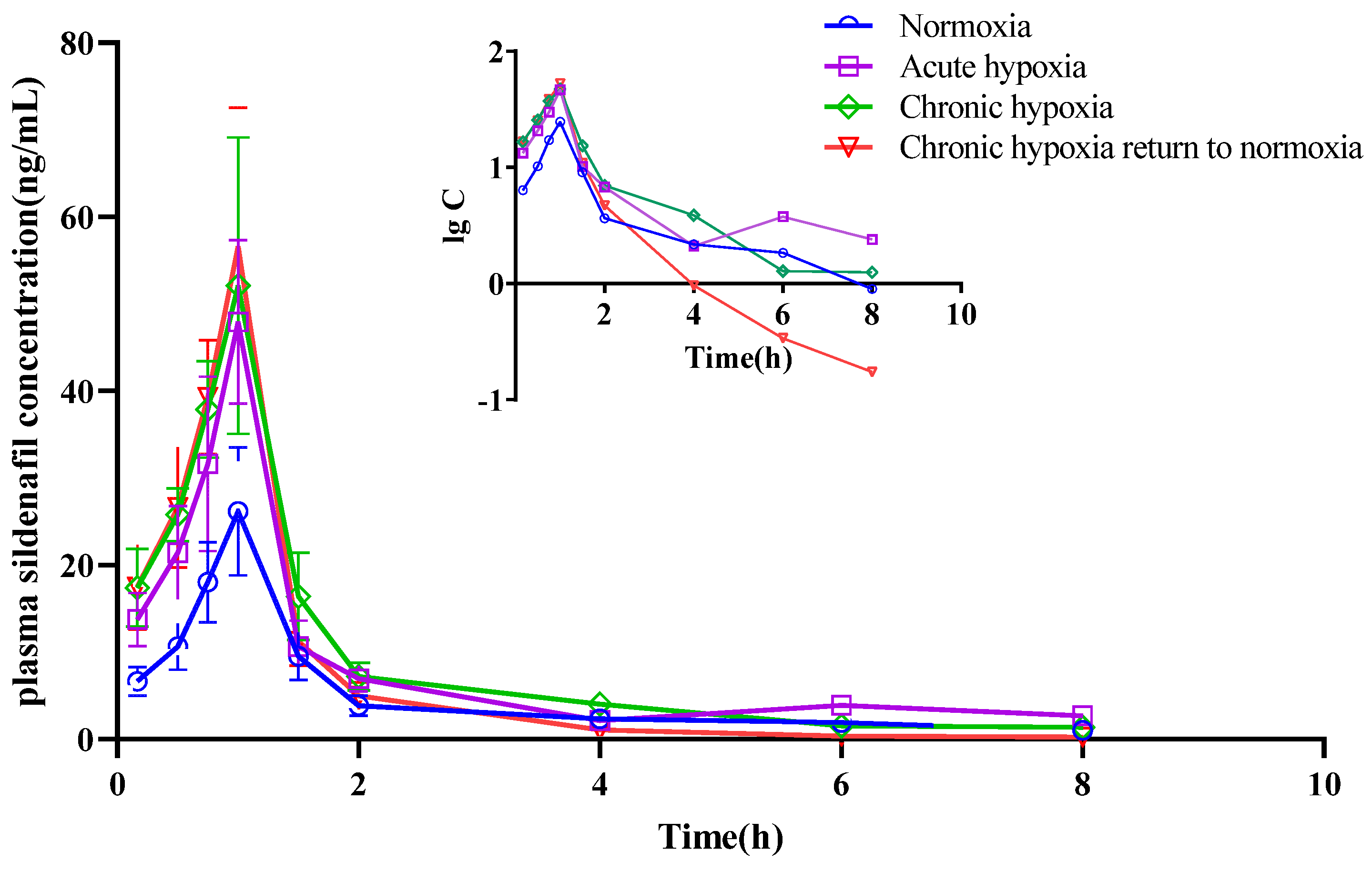
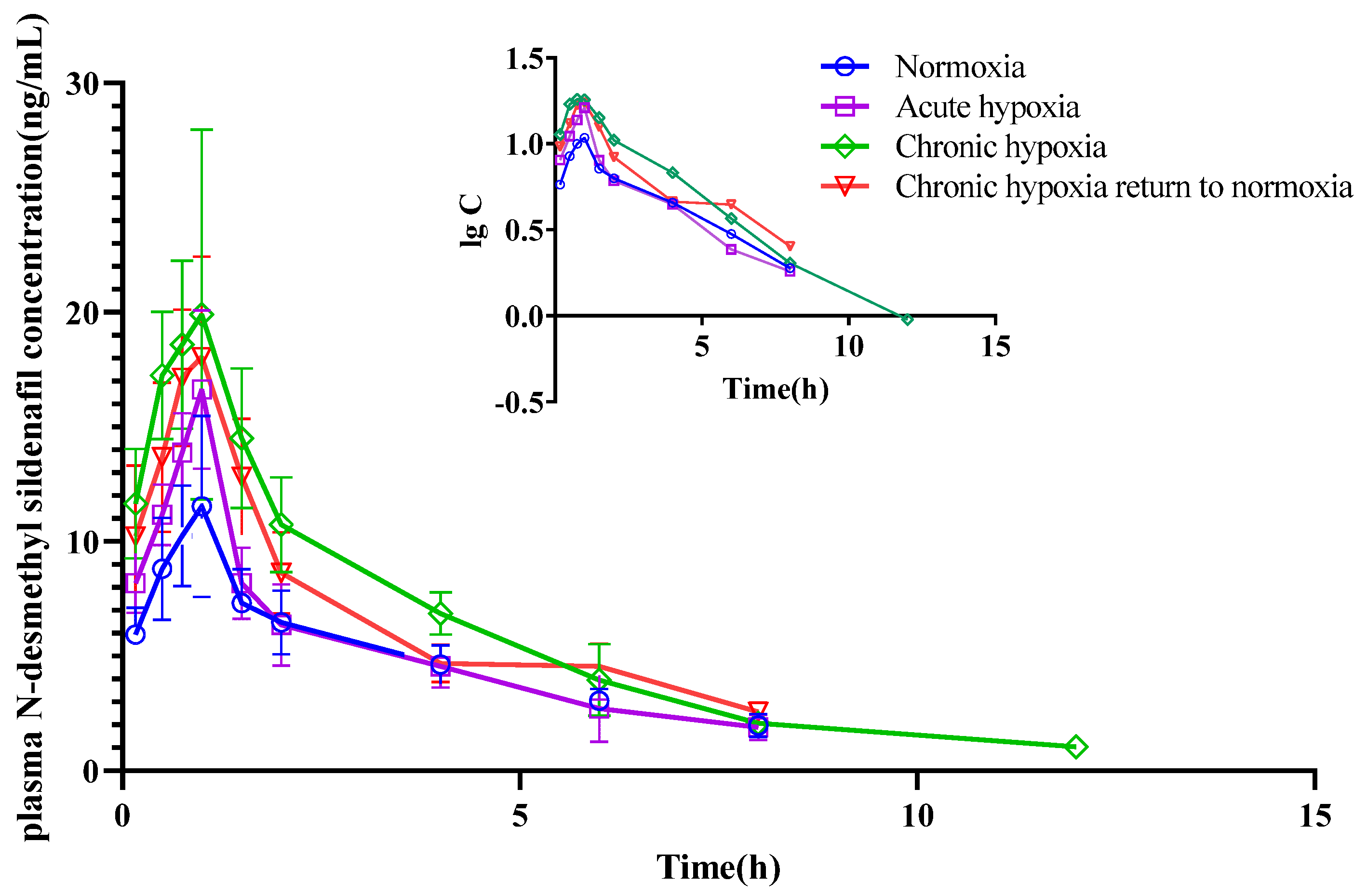
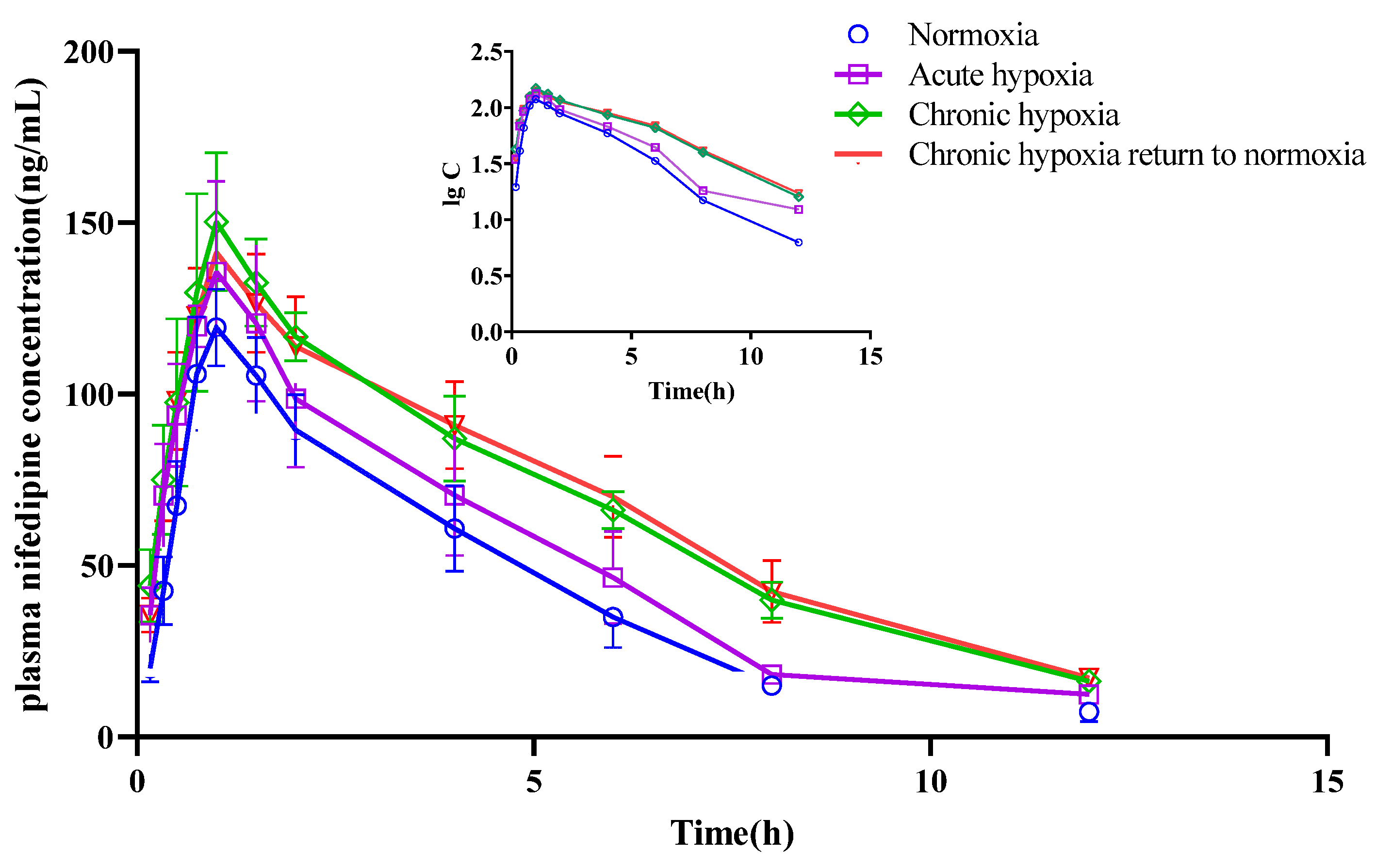
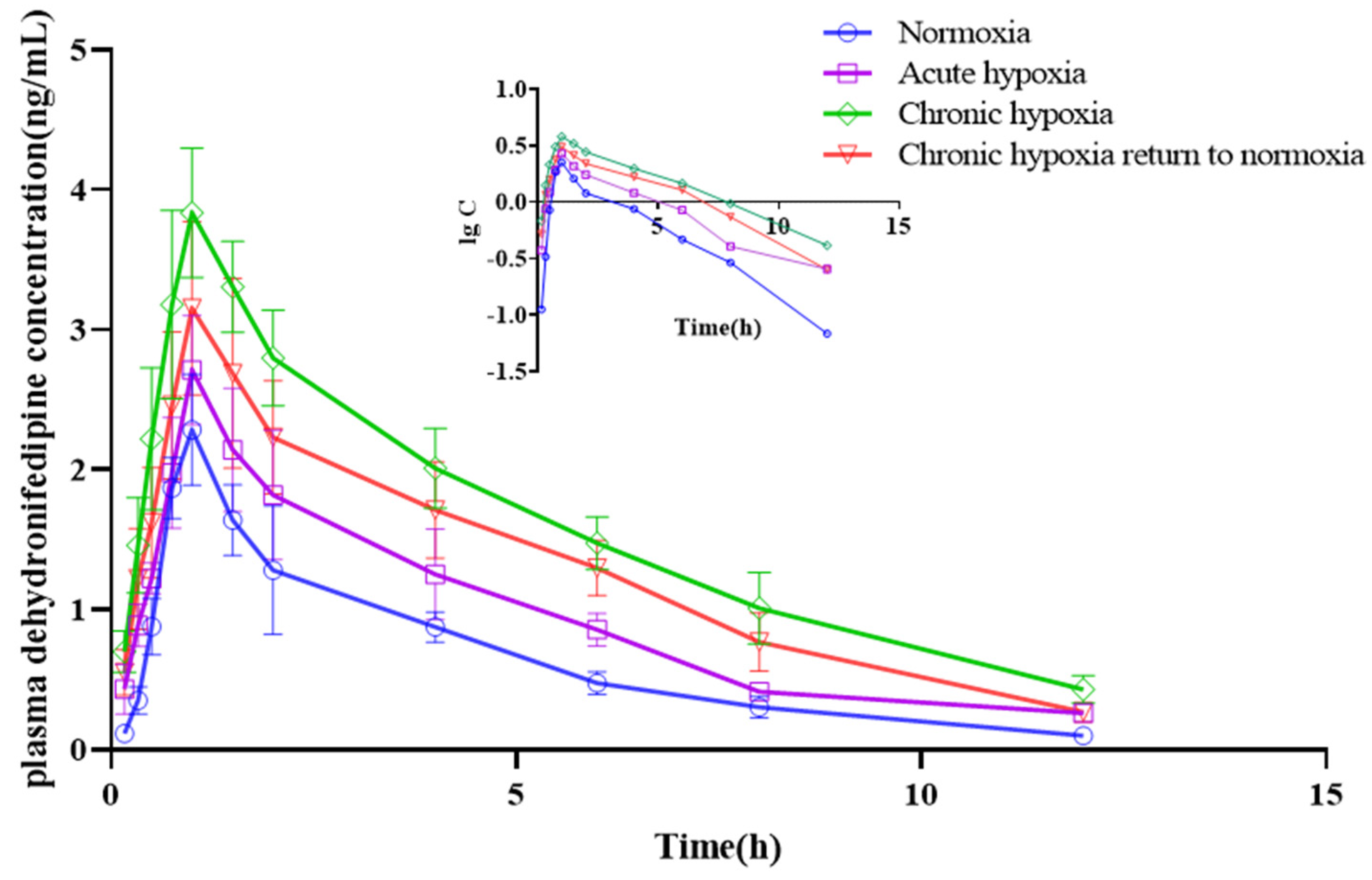
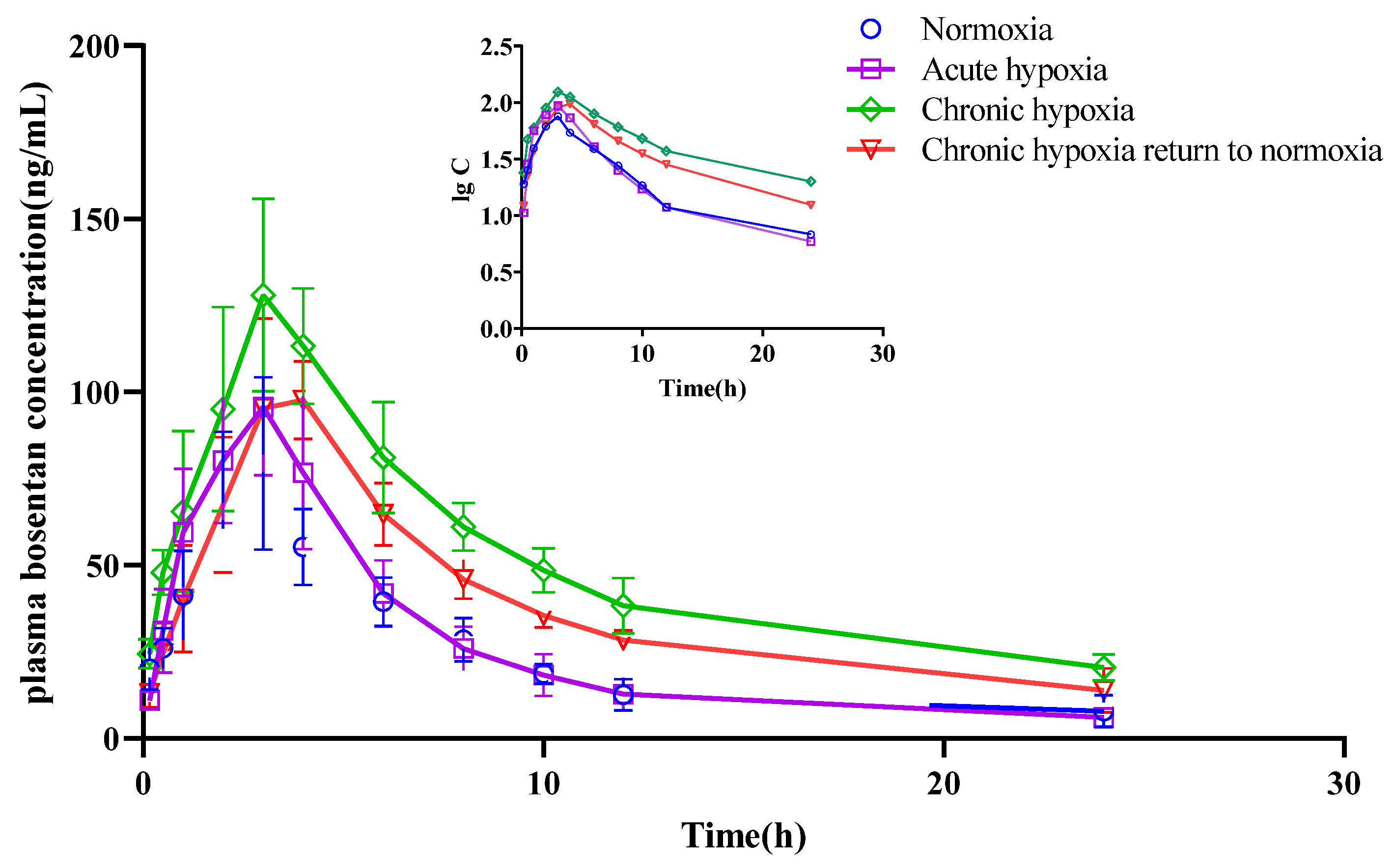
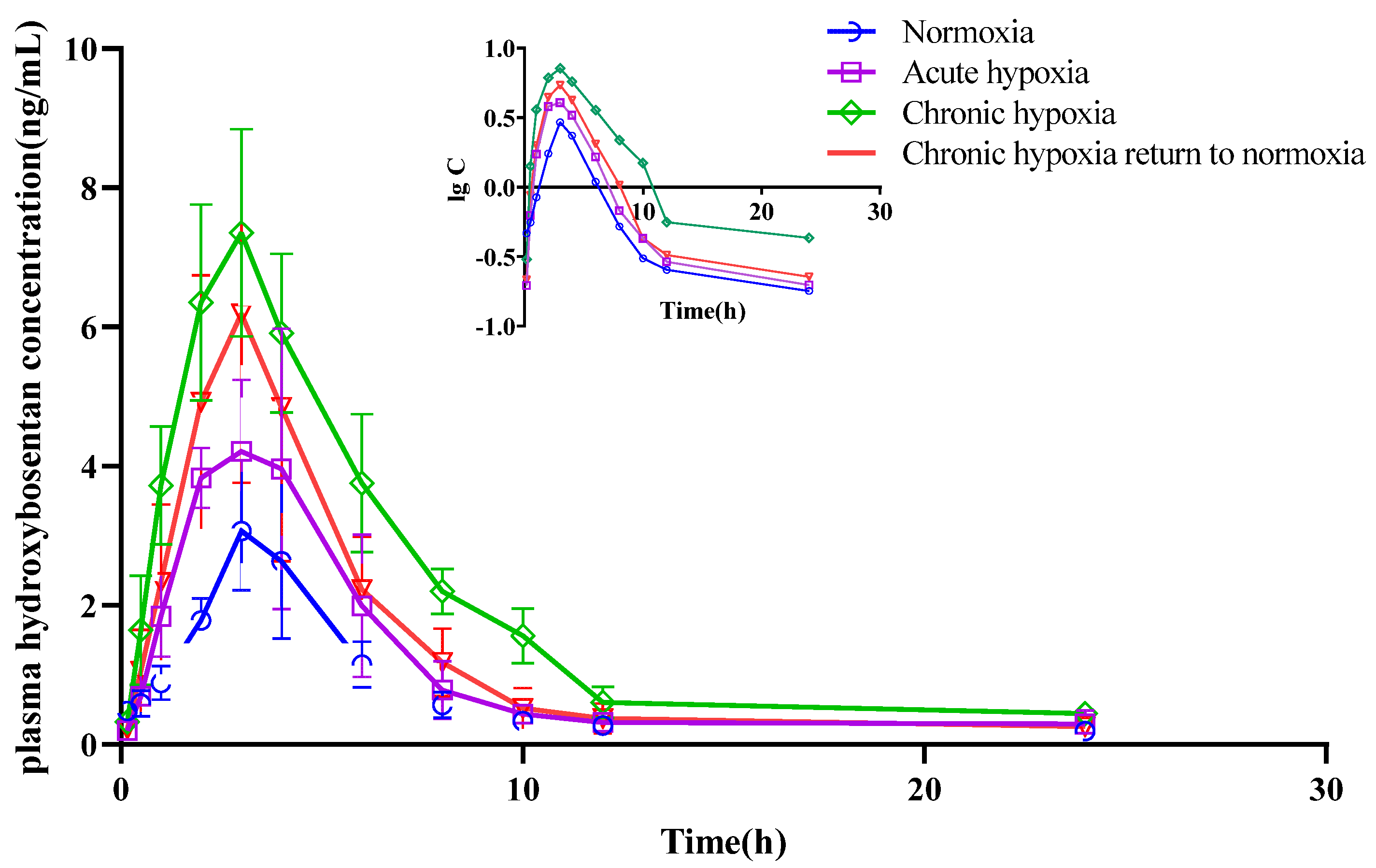
2.4. Pharmacokinetics
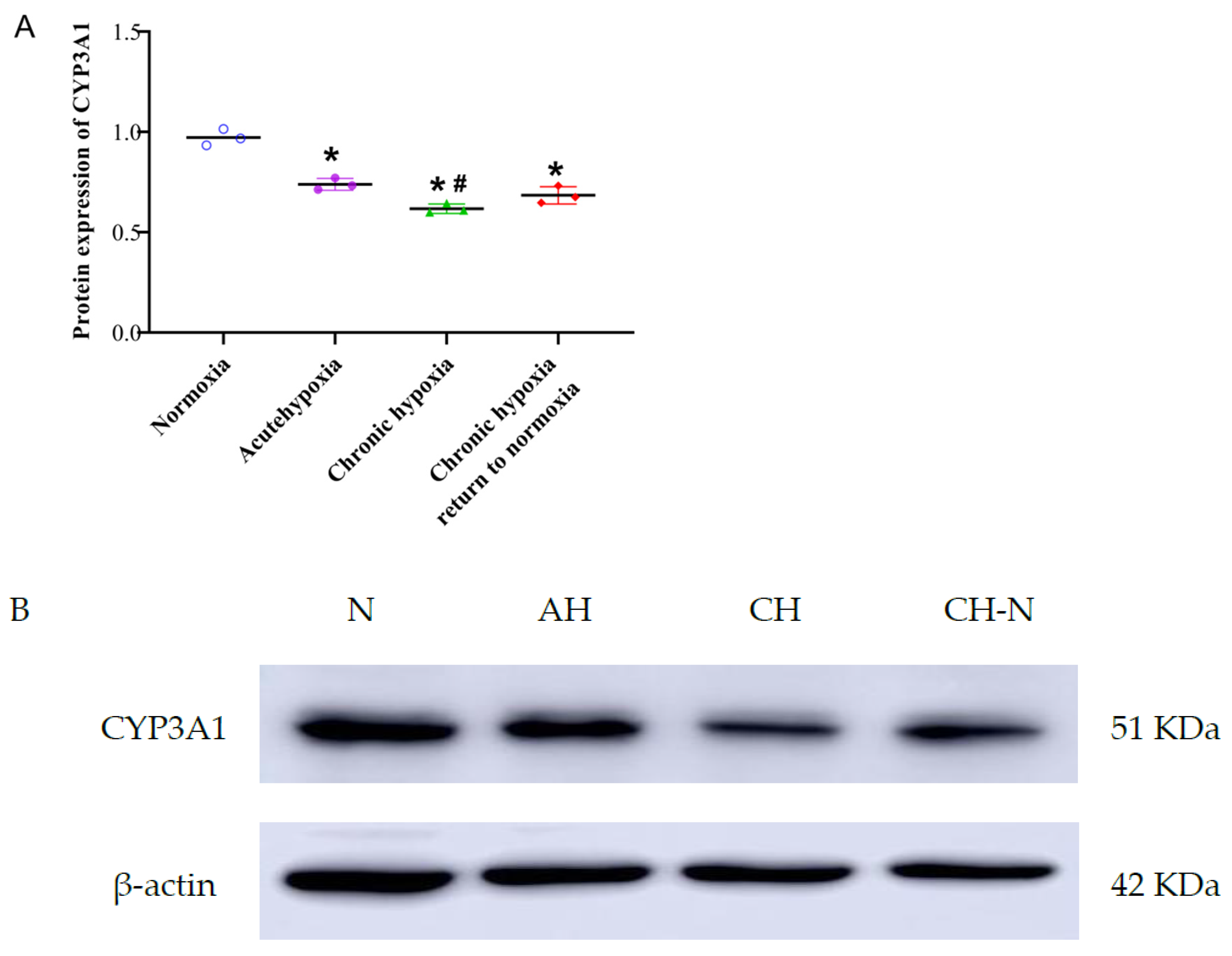
2.5. Protein Expression of CYP3A1
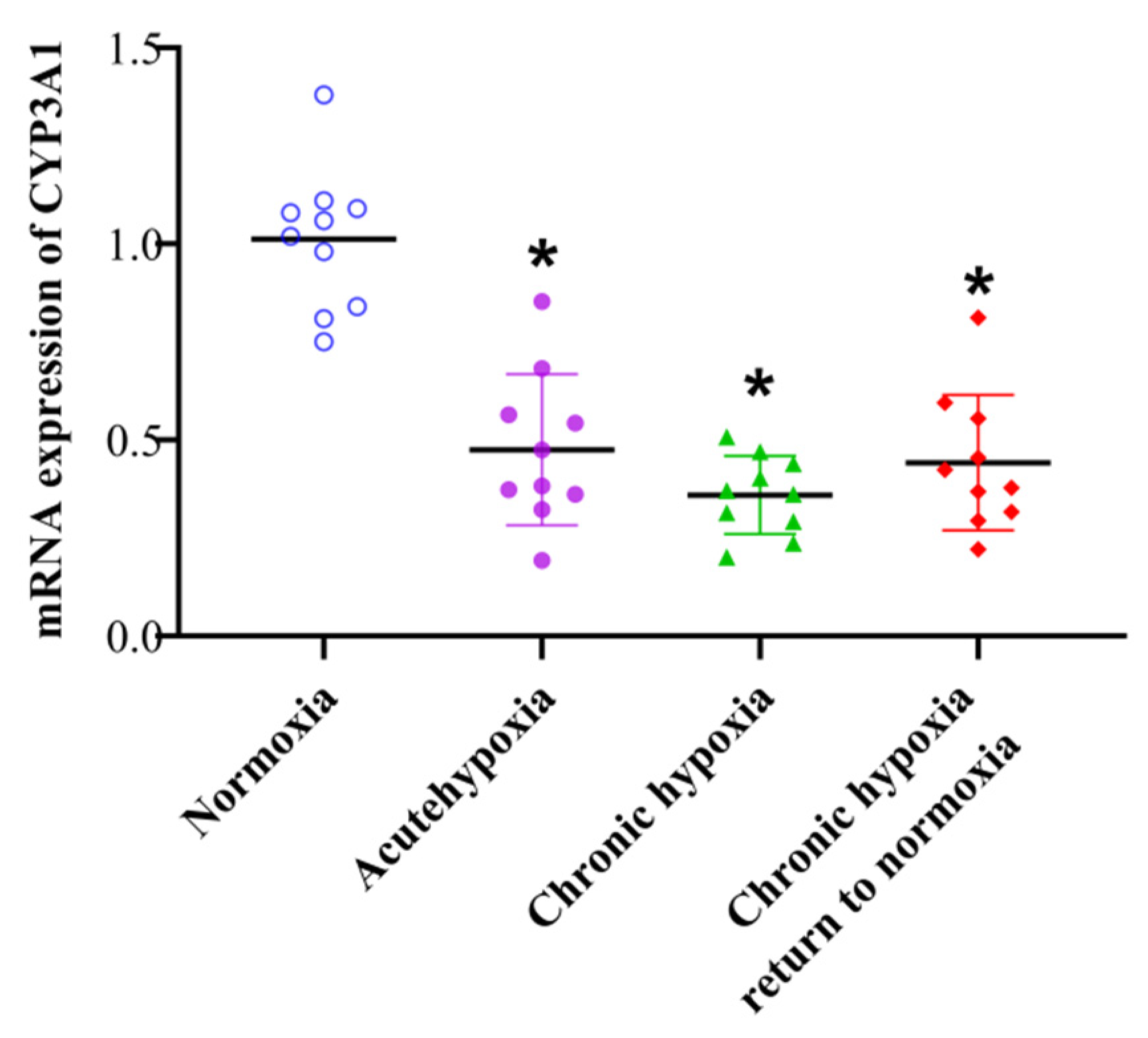
2.6. mRNA Expression of CYP3A1
3. Discussion
4. Materials and Methods
4.1. Reagents and Instruments
4.2. Animals and Experimental Treatments
4.3. Determination of Physiologic and Biochemical Parameters
4.4. Hematoxylin-Eosin Staining
4.5. Pharmacokinetic Study Design
4.6. Sample Processing
4.7. Chromatographic Conditions
4.8. Mass Spectrometry Conditions
4.9. UHPLC-MS Validation
4.10. Pharmacokinetic Analysis
4.11. Western Blot
4.12. Quantitative Real-Time Polymerase Chain Reaction
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, J.; He, N.; Wei, X.; Gao, Y.; Zuo, Y. Changes in Temperature Sensitivity and Activation Energy of Soil Organic Matter Decomposition in Different Qinghai-Tibet Plateau Grasslands. PLoS ONE 2015, 10, e0132795. [Google Scholar] [CrossRef] [PubMed]
- Eide 3rd, R.P.; Asplund, C.A. Altitude Illness: Update on Prevention and Treatment. Curr. Sport. Med. Rep. 2012, 11, 124–130. [Google Scholar] [CrossRef]
- Jiang, C.; Chen, J.; Liu, F.; Luo, Y.; Xu, G.; Shen, H.-Y.; Gao, W. Chronic mountain sickness in Chinese Han males who migrated to the Qinghai-Tibetan plateau: Application and evaluation of diagnostic criteria for chronic mountain sickness. BMC Public Health 2014, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manferdelli, G.; Marzorati, M.; Easton, C.; Porcelli, S. Changes in prefrontal cerebral oxygenation and microvascular blood volume in hypoxia and possible association with acute mountain sickness. Exp. Physiol. 2021, 106, 76–85. [Google Scholar] [CrossRef]
- Narvaez-Guerra, O.; Herrera-Enriquez, K.; Medina-Lezama, J.; Chirinos, J.A. Systemic Hypertension at High Altitude. Hypertension 2018, 72, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Chang, C.; Zhang, J.; Jiang, Y.; Ni, B.; Wang, Y. Prevalence and risk factors associated with hypertension and prehypertension in a working population at high altitude in China: A cross-sectional study. Environ. Health Prev. Med. 2017, 22. [Google Scholar] [CrossRef] [Green Version]
- Mingji, C.; Onakpoya, I.J.; Perera, R.; Ward, A.M.; Heneghan, C.J. Relationship between altitude and the prevalence of hypertension in Tibet: A systematic review. Heart 2015, 101, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Yue, L.; Fan, Z.; Sun, L.; Feng, W.; Li, J. Prevalence of Essential Hypertension and Its Complications Among Chinese Population at High Altitude. High Alt. Med. Biol. 2017, 18, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Chen, Q.; Wang, Y.; Wang, J.; Ren, Z.; Cao, Z.; Wang, B. Spatial pattern and variations in the prevalence of congenital heart disease in children aged 4–18 years in the Qinghai-Tibetan Plateau. Sci. Total Environ. 2018, 627, 158–165. [Google Scholar] [CrossRef]
- Sergi, C. EPAS1, Congenital heart disease, and high altitude: Disclosures by genetics, bioinformatics, and experimental embryology. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef]
- El-Sherif, N.; Turitto, G.; Boutjdir, M. Congenital Long QT syndrome and torsade de pointes. Ann. Noninvasive Electrocardiol. 2017, 22, e12481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, C.; Jihang, Z.; Yu, J.; Dai, L.; Ding, X.; Huang, L. PPARA genetic variants increase the risk for cardiac pumping function reductions following acute high-altitude exposure: A self-controlled study. Mol. Genet. Genom. Med. 2019, 7, e00919. [Google Scholar] [CrossRef] [Green Version]
- Bhattarai, P.; Paudel, B.H.; Thakur, D.; Bhattarai, B.; Subedi, B.; Khadka, R. Effect of long term high altitude exposure on cardiovascular autonomic adjustment during rest and post-exercise recovery. Ann. Occup. Environ. Med. 2018, 30. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Nian, Y.; Qiao, Y.; Yang, M.; Xin, Y.; Li, X. Hypoxia plays a key role in the pharmacokinetic changes of drugs at high altitude. Curr. Drug Metab. 2018, 19, 960–969. [Google Scholar] [CrossRef]
- Vij, A.G.; Kishore, K.; Dey, J. Effect of intermittent hypobaric hypoxia on efficacy & clearance of drugs. Indian J. Med. Res. 2012, 135, 211. [Google Scholar] [PubMed]
- Arancibia, A.; Gai, M.N.; Chávez, J.; Paulos, C.; Pinilla, E.; González, C.; Ritschel, W.A. Pharmacokinetics of prednisolone in man during acute and chronic exposure to high altitude. Int. J. Clin. Pharmacol. Ther. 2005, 43, 85–91. [Google Scholar] [CrossRef]
- Wei, L.; Li, L.; Zhang, B.; Ma, L. Propranolol Suppresses Cobalt Chloride-Induced Hypoxic Proliferation in Human Umbilical Vein Endothelial Cells in vitro. Pharmacology 2019, 103, 61–67. [Google Scholar] [CrossRef]
- Schroeder, T.; Piantadosi, C.A.; Natoli, M.J.; Autmizguine, J.; Cohen-Wolkowieczs, M.; Hamilton, K.L.; Noveck, R.J. Safety and ergogenic properties of combined aminophylline and ambrisentan in hypoxia. Clin. Pharmacol. 2018, 103, 888–898. [Google Scholar] [CrossRef] [Green Version]
- Tapia, L.; Irarrázaval, S. Acetazolamide for the treatment of acute mountain sickness. Medwave 2019, 19, e7737. [Google Scholar] [CrossRef]
- Li, X.; Gao, F.; Li, Z.; Guan, W.; Feng, W.; Ge, R. Comparison of the pharmacokinetics of sulfamethoxazole in male chinese volunteers at low altitude and acute exposure to high altitude versus subjects living chronically at high altitude: An open-label, controlled, prospective study. Clin. Ther. 2009, 31, 2744–2754. [Google Scholar] [CrossRef]
- Zhou, Y.; Wen, J.; Wang, G. Identification of cytochrome P450 isoenzymes involved in the metabolism of 23-hydroxybetulinic acid in human liver microsomes. Pharm. Biol. 2019, 58, 60–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikandan, P.; Nagini, S. Cytochrome P450 Structure, Function and Clinical Significance: A Review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P. Cytochrome P450 research and the journal of biological chemistry. J. Biol. Chem. 2019, 294, 1671–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tornio, A.; Backman, J.T. Cytochrome P450 in pharmacogenetics: An update. Adv. Pharmacol. 2018, 83, 3–32. [Google Scholar] [CrossRef] [Green Version]
- Guengerich, F.P.; Waterman, M.R.; Egli, M. Recent Structural Insights into Cytochrome P450 Function. Trends Pharmacol. Sci. 2016, 37, 625–640. [Google Scholar] [CrossRef] [Green Version]
- Kato, H. Computational prediction of cytochrome P450 inhibition and induction. Drug Metab. Pharmacokinet. 2019, 35, 30–44. [Google Scholar] [CrossRef]
- Park, J.W.; Choi, J.S. Role of kaempferol to increase bioavailability and pharmacokinetics of nifedipine in rats. Chin. J. Nat. Med. 2019, 17, 690–697. [Google Scholar] [CrossRef]
- Tubic-Grozdanis, M.; Hilfinger, J.M.; Amidon, G.L.; Kim, J.S.; Kijek, P.; Staubach, P.; Langguth, P. Pharmacokinetics of the CYP 3A Substrate Simvastatin following Administration of Delayed Versus Immediate Release Oral Dosage Forms. Pharm. Res. 2008, 25, 1591–1600. [Google Scholar] [CrossRef]
- Gillen, M.; Yang, C.; Wilson, D.; Valdez, S.; Lee, C.; Kerr, B.; Shen, Z. Evaluation of Pharmacokinetic Interactions Between Lesinurad, a New Selective Urate Reabsorption Inhibitor, and CYP Enzyme Substrates Sildenafil, Amlodipine, Tolbutamide, and Repaglinide. Clin. Pharmacol. Drug Dev. 2017, 6, 363–376. [Google Scholar] [CrossRef]
- Hyland, R.; Roe, E.G.H.; Jones, B.C.; Smith, D.A. Identification of the cytochrome P450 enzymes involved in the N-demethylation of sildenafil. Br. J. Clin. Pharmacol. 2008, 51, 239–248. [Google Scholar] [CrossRef]
- Chen, M.; Zhang, X.; Chen, Y.; Sun, W.; Wang, Z.; Huang, C.; Chen, R. Comparison of the Inhibitory Effect of Ketoconazole, Voriconazole, Fluconazole, and Itraconazole on the Pharmacokinetics of Bosentan and Its Corresponding Active Metabolite Hydroxy Bosentan in Rats. Xenobiotica 2019, 50, 280–287. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Li, Y.; Yuan, M.; Zhu, J.; Su, X.; Duan, Y. Effect of Exposure to Acute and Chronic High-Altitude Hypoxia on the Activity and Expression of CYP1A2, CYP2D6, CYP2C9, CYP2C19 and NAT2 in Rats. Pharmacology 2014, 93, 76–83. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Li, Y.; Zhu, J.; Su, X.; Yao, X.; Duan, Y. The Activity, Protein, and mRNA Expression of CYP2E1 and CYP3A1 in Rats after Exposure to Acute and Chronic High Altitude Hypoxia. High Alt. Med. Biol. 2014, 15, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Xia, L.; Jin, Z.; Jin, R.; Paudel, O.; Sham, J.S.K. Cytochrome P450 Epoxygenase-Dependent Activation of TRPV4 Channel Participates in Enhanced Serotonin-Induced Pulmonary Vasoconstriction in Chronic Hypoxic Pulmonary Hypertension. Anal. Cell. Pathol. 2020, 2020, 8927381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurdi, J.; Maurice, H.; El-Kadi, A.O.S.; Ong, H.; Dalkara, S.; Bélanger, P.M.; du Souich, P. Effect of hypoxia alone or combined with inflammation and 3-methylcholanthrene on hepatic cytochrome P450 in conscious rabbits. Br. J. Pharmacol. 1999, 128, 365–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zeng, Y.; Chen, X.; Zhang, Y.; Ding, J.; Xue, C. Decreased expression of hepatic cytochrome P450 1A2 (CYP1A2) in a chronic intermittent hypoxia mouse model. J. Thorac. Dis. 2018, 10, 825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Shao, H.; Wu, Q.; Sun, X.; Li, L.; Li, K.; Chen, H. Altered gene expression of hepatic cytochrome P450 in a rat model of intermittent hypoxia with emphysema. Mol. Med. Rep. 2017, 16, 881–886. [Google Scholar] [CrossRef] [Green Version]
- Kwapiszewska, G.; Johansen, A.K.Z.; Gomez-Arroyo, J.; Voelkel, N.F. The Role of the Aryl Hydrocarbon Receptor/ARNT/Cytochrome P450 System in Pulmonary Vascular Diseases. Circ. Res. 2019, 125, 356–366. [Google Scholar] [CrossRef]
- Zhou, Q.; Ruan, Z.R.; Jiang, B.; Yuan, H.; Zeng, S. Simvastatin pharmacokinetics in healthy Chinese subjects and its relations with CYP2C9, CYP3A5, ABCB1, ABCG2 and SLCO1B1 polymorphisms. Pharmazie 2013, 68, 124–128. [Google Scholar] [CrossRef]
- Humbert, M.; Simonneau, G. Sildenafil for Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2004, 169, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Abdelkawy, K.S.; Donia, A.M.; Turner, R.B.; Elbarbry, F. Effects of lemon and Seville orange juices on the pharmacokinetic properties of sildenafil in healthy subjects. Drugs RD 2016, 16, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivas, N.R. Clinical drug-drug interactions of bosentan, a potent endothelial receptor antagonist, with various drugs: Physiological role of enzymes and transporters. Gen. Physiol. Biophys. 2016, 35, 243–258. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, J.; Nian, Y.; Liu, G.; Duan, Y.; Bai, X.; Li, X. Pharmacokinetics of acetaminophen and metformin hydrochloride in rats after exposure to simulated high altitude hypoxia. Front. Pharmacol. 2021, 12, 692349. [Google Scholar] [CrossRef] [PubMed]
- Sarada, S.K.; Dipti, P.; Anju, B.; Pauline, T.; Kain, A.; Sairam, M.; Selvamurthy, W. Antioxidant effect of beta-carotene on hypoxia induced oxidative stress in male albino rats. J. Ethnopharmacol. 2002, 79, 149–153. [Google Scholar] [CrossRef]
- Schoutsen, B.; De Jong, J.W.; Harmsen, E.; De Tombe, P.P.; Achterberg, P.W. Myocardial xanthine oxidase/ dehydrogenase. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 1983, 762, 519–524. [Google Scholar] [CrossRef]
- Nepal, O.; Pokharel, B.; Khanal, K.; Mallik, S.; Kapoor, B.; Koju, R. Relationship Between Arterial Oxygen Saturation and Hematocrit, and Effect of Slow Deep Breathing on Oxygen Saturation in Himalayan High Altitude Populations. Kathmandu Univ. Med. J. 2013, 10, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Paterson, G.G.; Young, J.M.; Willson, J.A.; Graham, C.J.; Dru, R.C.; Lee, E.W.; Thompson, A.A.R. Hypoxia Modulates Platelet Purinergic Signalling Pathways. Thromb. Haemost. 2019, 120, 253–261. [Google Scholar] [CrossRef]
- Villafuerte, F.C. New genetic and physiological factors for excessive erythrocytosis and Chronic Mountain Sickness. J. Appl. Physiol. 2015, 119, 1481–1486. [Google Scholar] [CrossRef] [Green Version]
- Vij, A.G. Effect of prolonged stay at high altitude on platelet aggregation and fibrinogen levels. Platelets 2009, 20, 421–427. [Google Scholar] [CrossRef]
- Chen, S.; Du, C.; Shen, M.; Zhao, G.; Xu, Y.; Yang, K.; Wang, J. Sympathetic stimulation facilitates thrombopoiesis by promoting megakaryocyte adhesion, migration, and proplatelet formation. Blood 2015, 127, 1024–1035. [Google Scholar] [CrossRef]
- Zierk, J.; Arzideh, F.; Rechenauer, T.; Haeckel, R.; Rascher, W.; Metzler, M.; Rauh, M. Age and Sex-Specific Dynamics in 22 Hematologic and Biochemical Analytes from Birth to Adolescence. Clin. Chem. 2015, 61, 964–973. [Google Scholar] [CrossRef] [Green Version]
- Shin, M.K.; Yao, Q.; Jun, J.; Bevans-Fonti, S.; Yoo, D.Y.; Han, W.; Polotsky, V.Y. Carotid body denervation prevents fasting hyperglycemia during chronic intermittent hypoxia. J. Appl. Physiol. 2014, 117, 765–776. [Google Scholar] [CrossRef] [Green Version]
- Arestegui, A.H.; Fuquay, R.; Sirota, J.; Swenson, E.R.; Schoene, R.B.; Jefferson, J.A.; Escudero, E. High Altitude Renal Syndrome (HARS). J. Am. Soc. Nephrol. 2011, 22, 1963–1968. [Google Scholar] [CrossRef] [Green Version]
- Caravita, S.; Faini, A.; Bilo, G.; Revera, M.; Giuliano, A.; Gregorini, F.; Parati, G. Ischemic changes in exercise ECG in a hypertensive subject acutely exposed to high altitude. Possible role of a high-altitude induced imbalance in myocardial oxygen supply–demand. Int. J. Cardiol. 2014, 171, e100–e102. [Google Scholar] [CrossRef] [PubMed]
- Mix, J.; Elon, L.; Vi Thien Mac, V.; Flocks, J.; Economos, E.; Tovar-Aguilar, A.J.; McCauley, L.A. Hydration Status, Kidney Function, and Kidney Injury in Florida Agricultural Workers. J. Occup. Environ. Med. 2018, 60, e253–e260. [Google Scholar] [CrossRef] [PubMed]
- Plata, R.; Cornejo, A.; Arratia, C.; Anabaya, A.; Perna, A.; Dimitrov, B.D.; Remuzzi, G. Angiotensin-converting-enzyme inhibition therapy in altitude polycythaemia: A prospective randomised trial. Lancet 2002, 359, 663–666. [Google Scholar] [CrossRef]
- Yang, C.; Ling, H.; Zhang, M.; Yang, Z.; Wang, X.; Zeng, F.; Feng, J. Oxidative stress mediates chemical hypoxia-induced injury and inflammation by activating NF-κb-COX-2 pathway in HaCaT cells. Mol. Cells 2011, 31, 531–538. [Google Scholar] [CrossRef] [Green Version]
- Eichstaedt, C.A.; Mairbäurl, H.; Song, J.; Benjamin, N.; Fischer, C.; Dehnert, C.; Hinderhofer, K. Genetic Predisposition to High-Altitude Pulmonary Edema. High Alt. Med. Biol. 2020, 21, 28–36. [Google Scholar] [CrossRef]
- Seys, S.F.; Daenen, M.; Dilissen, E.; Van Thienen, R.; Bullens, D.M.A.; Hespel, P.; Dupont, L.J. Effects of high altitude and cold air exposure on airway inflammation in patients with asthma. Thorax 2013, 68, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Preissner, S.C.; Hoffmann, M.F.; Preissner, R.; Dunkel, M.; Gewiess, A.; Preissner, S. Polymorphic Cytochrome P450 Enzymes (CYPs) and Their Role in Personalized Therapy. PLoS ONE 2013, 8, e82562. [Google Scholar] [CrossRef]
- Sevrioukova, I.F.; Poulos, T.L. Understanding the mechanism of cytochrome P450 3A4: Recent advances and remaining problems. Dalton Trans. 2013, 42, 3116–3126. [Google Scholar] [CrossRef] [PubMed]
- Fradette, C.; Batonga, J.; Teng, S.; Piquette-Miller, M.; du Souich, P. Animal Models of Acute Moderate Hypoxia Are Associated with a Down-Regulation of CYP1A1, 1A2, 2B4, 2C5, and 2C16 and Up-Regulation of CYP3A6 and P-glycoprotein in Liver. Drug Metab. Dispos. 2007, 35, 765–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradette, C.; Souich, P.D. Hypoxia-inducible factor-1 and activator protein-1 modulate the upregulation of CYP3A6 induced by hypoxia. Br. J. Pharmacol. 2003, 140, 1146–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradette, C.; Yamaguchi, N.; du Souich, P. 5-Hydroxytryptamine is biotransformed by CYP2C9, 2C19 and 2B6 to hydroxylamine, which is converted into nitric oxide. Br. J. Pharmacol. 2004, 141, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Y.; Zhu, J.; Yang, J.; Gu, W.; Bai, X.; Liu, G.; Xiangyang, L. A decade’s review of miRNA: A center of transcriptional regulation of drugmetabolizing enzymes and transporters under hypoxia. Curr. Drug Metab. 2021, 22, 709–725. [Google Scholar] [CrossRef]
- Legendre, C.; Hori, T.; Loyer, P.; Aninat, C.; Ishida, S.; Glaise, D.; Morel, F. Drug-metabolising enzymes are down-regulated by hypoxia in differentiated human hepatoma HepaRG cells: HIF-1α involvement in CYP3A4 repression. Eur. J. Cancer 2009, 45, 2882–2892. [Google Scholar] [CrossRef]
- Duan, Y.; Zhu, J.; Yang, J.; Liu, G.; Bai, X.; Qu, N.; Li, X. Regulation of High-Altitude Hypoxia on the Transcription of CYP450 and UGT1A1 Mediated by PXR and CAR. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Duan, Y.; Bai, X.; Yang, J.; Zhou, Y.; Gu, W.; Liu, G.; Li, X. Exposure to High-Altitude Environment Is Associated with Drug Transporters Change: microRNA-873-5p-Mediated Alteration of Function and Expression Levels of Drug Transporters under Hypoxia. Drug Metab. Dispos. 2022, 50, 174–186. [Google Scholar] [CrossRef]
- Li, H.; He, J.; Jia, W. The influence of gut microbiota on drug metabolism and toxicity. Expert Opin. Drug Metab. Toxicol. 2015, 12, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhang, J.; Wang, R.; Jia, Z. Effects of gut microbiota on drug metabolism and guidance for rational drug use under hypoxic conditions at high altitudes. Curr. Drug Metab. 2019, 20, 155–165. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Y.; Sun, Y.; Wang, R.; Zhang, J.; Jia, Z. Plateau hypoxia attenuates the metabolic activity of intestinal flora to enhance the bioavailability of nifedipine. Drug Deliv. 2018, 25, 1175–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Bai, X.; Duan, Y.; Zhu, J.; Yang, J.; Wang, Q.; Li, X. Changes in the intestinal flora of rats under high altitude hypoxia. Acta Pharm. Sin. 2021, 56, 1100–1108. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 810.05 ± 146.22 | 1127.94 ± 317.36 | 1268.96 ± 177.10 | 1239.53 ± 566.75 |
| AUC(0–∞), h·ug/L | 842.00 ± 148.01 | 1191.85 ± 334.85 | 1377.89 ± 199.13 a | 1317.55 ± 594.18 |
| AUMC(0–24), h2·ug/L | 5124.82 ± 992.08 | 8041.01 ± 2385.49 | 8895.72 ± 1463.89 a | 8854.69 ± 4261.74 a |
| AUMC(0–∞), h2·ug/L | 6419.60 ± 1258.50 | 11,267.41 ± 3647.81 a | 12,561.54 ± 2533.47 a | 11,797.45 ± 5201.98 a |
| MRT(0–24), h | 6.32 ± 0.60 | 7.15 ± 0.86 a | 7.00 ± 0.54 | 7.08 ± 0.70 a |
| MRT(0–∞), h | 7.63 ± 0.89 | 9.53 ± 2.06 a | 9.07 ± 0.98 a | 9.02 ± 1.26 |
| t1/2z, h | 4.99 ± 1.16 | 5.37 ± 1.11 | 6.55 ± 0.92 ab | 5.61 ± 0.93 |
| Tmax, h | 1.75 ± 0.46 | 1.44 ± 0.62 | 2.50 ± 0.93 b | 2.25 ± 1.16 |
| CLz/F, L/h/kg | 12.29 ± 2.73 | 8.90 ± 2.14 a | 7.42 ± 1.33 a | 9.04 ± 3.91 a |
| Vz/F, L/kg | 89.22 ± 29.43 | 68.84 ± 19.49 | 69.76 ± 12.46 | 72.80 ± 29.95 |
| λz, 1/h | 0.15 ± 0.05 | 0.13 ± 0.03 | 0.11 ± 0.02 a | 0.13 ± 0.02 |
| Cmax, ug/L | 107.82 ± 30.16 | 133.29 ± 34.51 | 156.23 ± 26.82 a | 134.74 ± 50.76 |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 242.33 ± 55.06 | 289.41 ± 77.21 | 533.64 ± 76.15 ab | 382.52 ± 62.29 abc |
| AUC(0–∞), h·ug/L | 284.51 ± 63.54 | 364.93 ± 113.48 | 560.05 ± 84.85 ab | 399.30 ± 64.67 ac |
| AUMC(0–24), h2·ug/L | 1034.71 ± 226.74 | 1325.11 ± 374.48 | 3398.47 ± 734.33 ab | 2262.16 ± 552.26 abc |
| AUMC(0–∞), h2·ug/L | 1860.09 ± 554.91 | 2635.37 ± 1238.53 | 4377.03 ± 1158.46 ab | 3043.17 ± 1072.49 ac |
| MRT(0–24), h | 4.27 ± 0.31 | 4.58 ± 0.30 | 6.33 ± 0.63 ab | 5.91 ± 1.10 ab |
| MRT(0–∞), h | 6.55 ± 1.29 | 7.03 ± 1.05 | 7.75 ± 1.06 | 7.60 ± 2.34 |
| t1/2z, h | 4.68 ± 1.27 | 5.00 ± 0.82 | 5.44 ± 1.54 | 5.22 ± 1.37 |
| Tmax, h | 2.00 ± 0.00 | 1.88 ± 0.35 | 2.12 ± 0.83 | 2.12 ± 0.84 |
| CLz/F, L/h/kg | 36.69 ± 7.97 | 30.62 ± 13.14 | 18.22 ± 2.82 ab | 25.53 ± 3.44 a |
| Vz/F, L/kg | 249.82 ± 97.60 | 213.18 ± 70.26 | 143.73 ± 47.89 a | 190.51 ± 52.21 |
| λz, 1/h | 0.16 ± 0.04 | 0.14 ± 0.02 | 0.14 ± 0.05 | 0.15 ± 0.06 |
| Cmax, ug/L | 41.29 ± 13.14 | 50.38 ± 14.76 | 73.94 ± 10.53 ab | 63.20 ± 17.89 a |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 37.46 ± 6.68 | 63.56 ± 8.71 a | 73.62 ± 13.42 a | 65.40 ± 13.34 a |
| AUC(0–∞), h·ug/L | 42.88 ± 7.86 | 76.85 ± 8.14 a | 80.61 ± 17.50 a | 73.91 ± 16.71 a |
| AUMC(0–24), h2·ug/L | 81.84 ± 16.63 | 141.81 ± 27.98 a | 175.85 ± 46.60 a | 120.72 ± 34.95 c |
| AUMC(0–∞), h2·ug/L | 145.62 ± 47.35 | 337.77 ± 121.66 a | 310.52 ± 171.71 a | 261.06 ± 179.05 |
| MRT(0–24), h | 2.19 ± 0.27 | 2.23 ± 0.32 | 2.37 ± 0.33 | 1.85 ± 0.38 abc |
| MRT(0–∞), h | 3.42 ± 1.04 | 4.36 ± 1.35 | 3.73 ± 1.31 | 3.39 ± 1.52 |
| t1/2z, h | 3.22 ± 1.06 | 3.44 ± 1.03 | 3.69 ± 0.88 | 3.60 ± 1.85 |
| Tmax, h | 0.94 ± 0.12 | 0.97 ± 0.09 | 0.94 ± 0.12 | 0.97 ± 0.09 |
| CLz/F, L/h/kg | 240.34 ± 45.04 | 13,150 ± 14.78 a | 129.83 ± 31.14 a | 141.55 ± 32.28 a |
| Vz/F, L/kg | 1096.51 ± 346.56 | 651.80 ± 190.65 a | 677.15 ± 164.99 a | 722.91 ± 368.21 a |
| λz, 1/h | 0.23 ± 0.07 | 0.22 ± 0.08 | 0.20 ± 0.05 | 0.23 ± 0.09 |
| Cmax, ug/L | 27.64 ± 6.70 | 49.89 ± 8.28 a | 58.03 ± 10.66 a | 58.06 ± 16.10 a |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 39.30 ± 7.26 | 42.36 ± 9.23 | 71.30 ± 8.60 ab | 54.45 ± 9.51 abc |
| AUC(0–∞), h·ug/L | 50.11 ± 9.99 | 53.03 ± 15.10 | 80.34 ± 12.41 ab | 69.36 ± 11.86 ab |
| AUMC(0–24), h2·ug/L | 119.08 ± 18.82 | 117.10 ± 36.76 | 254.22 ± 39.18 ab | 156.50 ± 28.71 abc |
| AUMC(0–∞), h2·ug/L | 264.43 ± 91.40 | 270.03 ± 160.34 | 462.33 ± 157.79 ab | 374.28 ± 169.66 |
| MRT(0–24), h | 3.05 ± 0.22 | 2.74 ± 0.27 a | 3.56 ± 0.28 ab | 2.88 ± 0.13 c |
| MRT(0–∞), h | 5.19 ± 1.07 | 4.88 ± 1.30 | 5.64 ± 1.22 | 5.29 ± 1.77 |
| t1/2z, h | 3.56 ± 0.92 | 3.67 ± 1.12 | 3.66 ± 1.21 b | 3.81 ± 1.62 |
| Tmax, h | 0.91 ± 0.13 | 0.94 ± 0.12 | 0.81 ± 0.22 | 0.97 ± 0.28 |
| CLz/F, L/h/kg | 207.02 ± 43.32 | 198.12 ± 39.05 | 127.05 ± 19.20 ab | 148.14 ± 26.80 ab |
| Vz/F, L/kg | 1038.91 ± 239.58 | 1019.96 ± 262.29 | 646.96 ± 159.90 ab | 793.51 ± 280.15 |
| λz, 1/h | 0.21 ± 0.07 | 0.20 ± 0.06 | 0.21 ± 0.08 | 0.20 ± 0.06 |
| Cmax, ug/L | 12.08 ± 4.34 | 17.16 ± 3.82 | 22.12 ± 7.83 a | 19.61 ± 4.03 a |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 511.92 ± 56.80 | 611.57 ± 75.70 a | 799.00 ± 60.34 ab | 811.72 ± 71.18 ab |
| AUC(0–∞), h·ug/L | 544.41 ± 67.37 | 664.25 ± 69.52 a | 881.48 ± 71.37 ab | 896.95±77.35 ab |
| AUMC(0–24), h2·ug/L | 1820.82 ± 279.73 | 2247.74 ± 286.46 a | 3328.84 ± 277.18 ab | 3471.50 ± 395.30 ab |
| AUMC(0–∞), h2·ug/L | 2315.83 ± 514.25 | 3173.46 ± 320.66 a | 4719.59 ± 674.31 ab | 4904.58 ± 493.11 ab |
| MRT(0–24), h | 3.54 ± 0.24 | 3.68 ± 0.26 | 4.17 ± 0.18 ab | 4.27 ± 0.21 ab |
| MRT(0–∞), h | 4.21 ± 0.56 | 4.81 ± 0.57 a | 5.34 ± 0.51 ab | 5.46 ± 0.20 ab |
| t1/2z, h | 2.75 ± 0.47 | 3.29 ± 0.55 a | 3.41 ± 0.48 a | 3.40 ± 0.29 a |
| Tmax, h | 0.94 ± 0.17 | 1.06 ± 0.29 | 1.09 ± 0.26 | 1.03 ± 0.21 |
| CLz/F, L/h/kg | 3.73 ± 0.50 | 3.04 ± 0.28 a | 2.28 ± 0.175 ab | 2.24 ± 0.20 ab |
| Vz/F, L/kg | 14.64 ± 2.30 | 14.52 ± 3.36 | 11.21 ± 1.67 ab | 10.98 ± 1.19 ab |
| λz, 1/h | 0.26 ± 0.05 | 0.22 ± 0.03 a | 0.21 ± 0.03 a | 0.20 ± 0.02 a |
| Cmax, ug/L | 122.36 ± 11.87 | 145.16 ± 25.74 | 162.14 ± 20.13 a | 143.50 ± 11.51 |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 7.72 ± 0.71 | 11.17 ± 1.26 a | 19.06 ± 2.46 ab | 15.43 ± 2.45 abc |
| AUC(0–∞), h·ug/L | 8.20 ± 0.79 | 12.19 ± 1.18 a | 21.42 ± 3.14 ab | 17.17 ± 3.58 abc |
| AUMC(0–24), h2·ug/L | 27.29 ± 3.17 | 42.90 ± 5.49 a | 80.12 ± 11.97 ab | 63.61 ± 10.37 abc |
| AUMC(0–∞), h2·ug/L | 34.01 ± 8.78 | 61.65 ± 10.54 a | 119.36 ± 24.90 ab | 87.16 ± 22.26 abc |
| MRT(0–24), h | 3.54 ± 0.30 | 3.84 ± 0.21 a | 4.20 ± 0.15 ab | 4.14 ± 0.29 ab |
| MRT(0–∞), h | 4.13 ± 0.89 | 5.05 ± 0.71 a | 5.54 ± 0.58 a | 5.09 ± 0.73 a |
| t1/2z, h | 2.68 ± 0.95 | 3.22 ± 0.80 | 3.54 ± 0.57 | 3.25 ± 1.16 |
| Tmax, h | 0.91 ± 0.13 | 1.16 ± 0.30 a | 1.03 ± 0.21 | 1.03 ± 0.21 |
| CLz/F, L/h/kg | 245.99 ± 25.34 | 165.37 ± 15.95 a | 95.13 ± 13.88 ab | 120.46 ± 22.30 abc |
| Vz/F, L/kg | 938.91 ± 301.42 | 772.88 ± 227.86 | 480.24 ± 75.72 a | 553.99 ± 190.27 a |
| λz, 1/h | 0.29 ± 0.11 | 0.23 ± 0.06 | 0.20 ± 0.04 a | 0.24 ± 0.10 |
| Cmax, ug/L | 2.31 ± 0.46 | 2.91 ± 0.27 | 3.98 ± 0.37 ab | 3.20 ± 0.76 ac |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 581.65 ± 160.97 | 649.88 ± 97.27 | 1187.88 ± 216.95 ab | 926.65 ± 157.68 abc |
| AUC(0–∞), h·ug/L | 613.97 ± 161.62 | 692.88 ± 120.78 | 1308.92 ± 241.35 ab | 994.02 ± 153.67 abc |
| AUMC(0–24), h2·ug/L | 4243.09 ± 1550.07 | 4196.97 ± 573.20 | 9506.62 ± 2176.34 ab | 7653.48 ± 1632.40 ab |
| AUMC(0–∞), h2·ug/L | 6180.89 ± 2829.55 | 5881.39 ± 1509.27 | 14,668.03 ± 3880.91 ab | 11,619.75 ± 3562.03 ab |
| MRT(0–24), h | 7.19 ± 0.68 | 6.52 ± 0.84 | 7.95 ± 0.72 ab | 8.22 ± 0.45 ab |
| MRT(0–∞), h | 9.79 ± 1.73 | 8.48 ± 1.39 | 11.08 ± 1.36 b | 11.51 ± 1.81 ab |
| t1/2z, h | 5.49 ± 1.77 | 5.64 ± 2.05 | 6.68 ± 1.83 | 6.19 ± 1.03 |
| Tmax, h | 3.25 ± 0.46 | 2.88± 0.35 | 3.12 ± 0.64 | 3.62 ± 0.52 b |
| CLz/F, L/h/kg | 34.31 ± 7.70 | 29.68 ± 5.45 | 15.74 ± 2.89 ab | 20.53 ± 3.02 ab |
| Vz/F, L/kg | 270.40 ± 111.22 | 232.04 ± 63.22 | 150.37 ± 42.16 a | 184.23 ± 45.28 |
| λz, 1/h | 0.14 ± 0.05 | 0.14 ± 0.06 | 0.11 ± 0.04 | 0.12 ± 0.02 |
| Cmax, ug/L | 82.49 ± 27.35 | 98.11 ± 23.74 | 135.30 ± 28.58 ab | 107.07 ± 19.24 c |
| Parameters | Normoxia | Acute Hypoxia | Chronic Hypoxia | Chronic Hypoxia to Normoxia |
|---|---|---|---|---|
| AUC(0–24), h·ug/L | 16.90 ± 3.33 | 26.10 ± 6.43 a | 48.12 ± 7.13 ab | 32.67 ± 10.41 ac |
| AUC(0–∞), h·ug/L | 17.52 ± 3.51 | 27.37 ± 6.51 a | 51.27 ± 6.16 ab | 34.52 ± 11.72 ac |
| AUMC(0–24), h2·ug/L | 109.00 ± 28.12 | 159.46 ± 48.60 a | 299.63 ± 37.11 ab | 183.96 ± 61.42 ac |
| AUMC(0–∞), h2·ug/L | 151.43 ± 39.09 | 231.94 ± 98.27 | 424.66 ± 67.64 ab | 261.46 ± 135.34 ac |
| MRT(0–24), h | 6.41 ± 0.56 | 6.16 ± 1.34 | 6.27 ± 0.46 | 5.65 ± 0.67 |
| MRT(0–∞), h | 8.67 ± 1.56 | 8.62 ± 3.26 | 8.41 ± 1.93 | 7.35 ± 1.94 |
| t1/2z, h | 5.04 ± 2.66 | 5.60 ± 2.30 | 5.55 ± 2.33 | 5.28 ± 3.23 |
| Tmax, h | 3.25 ± 0.46 | 3.00 ± 0.93 | 2.62 ± 0.52 | 3.25 ± 0.46 |
| CLz/F, L/h/kg | 1183.16 ± 239.74 | 766.53 ± 176.85 a | 395.84 ± 54.95 ab | 667.57 ± 322.49 ac |
| Vz/F, L/kg | 8636.86 ± 5128.39 | 6360.02 ± 3074.79 | 3221.34 ± 1700.78 a | 4577.55 ± 2353.37 |
| λz, 1/h | 0.17 ± 0.08 | 0.15 ± 0.09 | 0.15 ± 0.07 | 0.18 ± 0.11 |
| Cmax, ug/L | 3.57 ± 1.06 | 5.22 ± 1.43 | 7.92 ± 1.49 ab | 6.79 ± 2.51 a |
| Time (min) | Acetonitrile | 0.2% Formic Acid-Water |
|---|---|---|
| 0.0 | 5 | 95 |
| 5.0 | 5 | 95 |
| 10.0 | 100 | 0 |
| 10.1 | 5 | 95 |
| 12.0 | 5 | 95 |
| Time (min) | Acetonitrile | 0.2% Formic Acid-Water |
|---|---|---|
| 0.0 | 5 | 95 |
| 5.0 | 5 | 95 |
| 10.0 | 100 | 0 |
| 10.1 | 5 | 95 |
| 12.0 | 5 | 95 |
| Time (min) | Acetonitrile | 0.2% Formic Acid-Water |
|---|---|---|
| 0.0 | 5 | 95 |
| 3.0 | 5 | 95 |
| 5.0 | 95 | 5 |
| 7.0 | 95 | 5 |
| 7.1 | 5 | 95 |
| Time (min) | Acetonitrile | 0.2% Formic Acid-Water |
|---|---|---|
| 0.0 | 10 | 90 |
| 1.0 | 10 | 90 |
| 6.0 | 100 | 0 |
| 7.0 | 100 | 0 |
| 7.1 | 10 | 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Duan, Y.; Duo, D.; Yang, J.; Bai, X.; Liu, G.; Wang, Q.; Wang, X.; Qu, N.; Zhou, Y.; et al. High-altitude Hypoxia Influences the Activities of the Drug-Metabolizing Enzyme CYP3A1 and the Pharmacokinetics of Four Cardiovascular System Drugs. Pharmaceuticals 2022, 15, 1303. https://doi.org/10.3390/ph15101303
Zhu J, Duan Y, Duo D, Yang J, Bai X, Liu G, Wang Q, Wang X, Qu N, Zhou Y, et al. High-altitude Hypoxia Influences the Activities of the Drug-Metabolizing Enzyme CYP3A1 and the Pharmacokinetics of Four Cardiovascular System Drugs. Pharmaceuticals. 2022; 15(10):1303. https://doi.org/10.3390/ph15101303
Chicago/Turabian StyleZhu, Junbo, Yabin Duan, Delong Duo, Jianxin Yang, Xue Bai, Guiqin Liu, Qian Wang, Xuejun Wang, Ning Qu, Yang Zhou, and et al. 2022. "High-altitude Hypoxia Influences the Activities of the Drug-Metabolizing Enzyme CYP3A1 and the Pharmacokinetics of Four Cardiovascular System Drugs" Pharmaceuticals 15, no. 10: 1303. https://doi.org/10.3390/ph15101303