

Diatom Indicators of Fluctuating/Intermittent Discharge from Springs in Two Bavarian Nature Conservation Areas

 , ,
, ,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
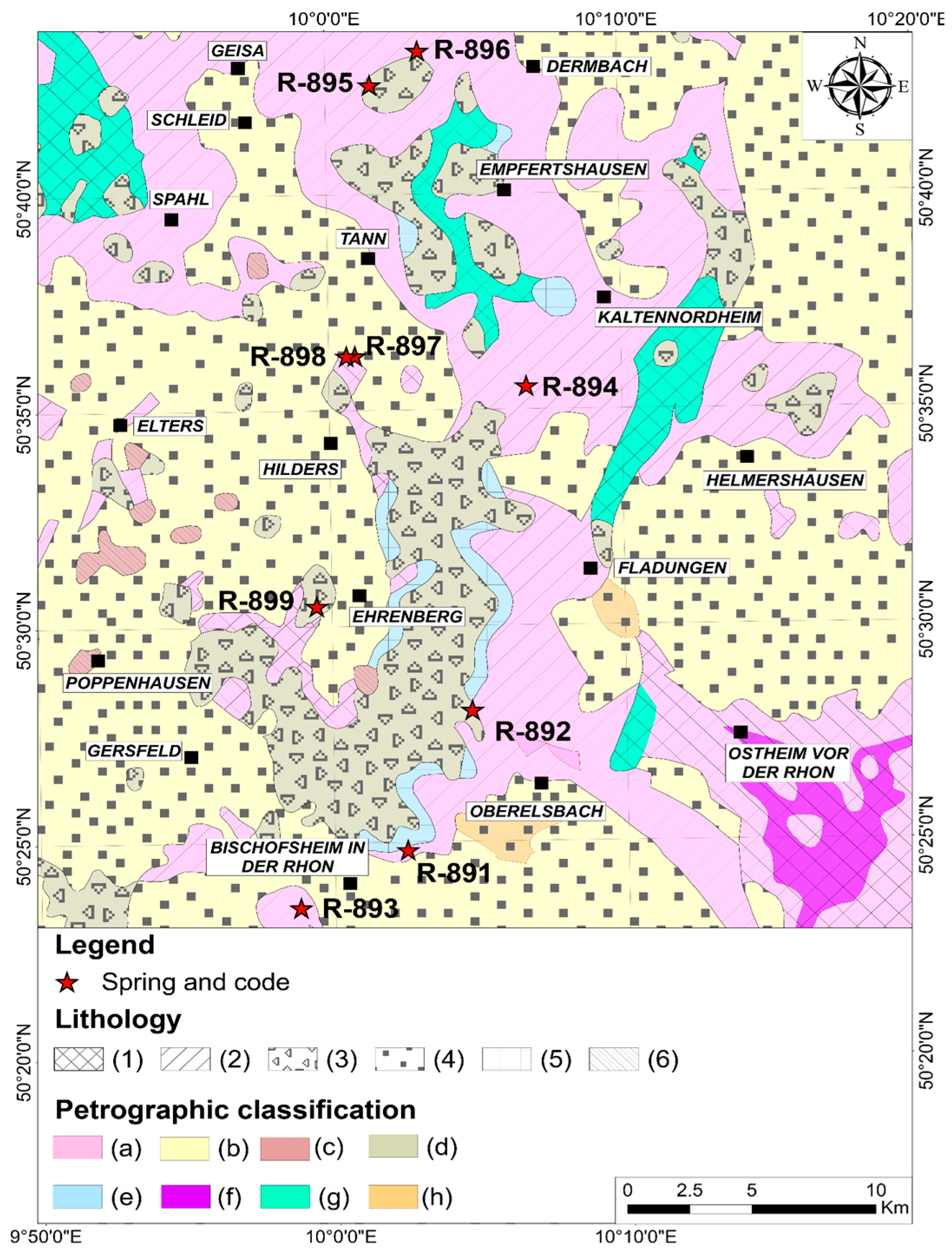
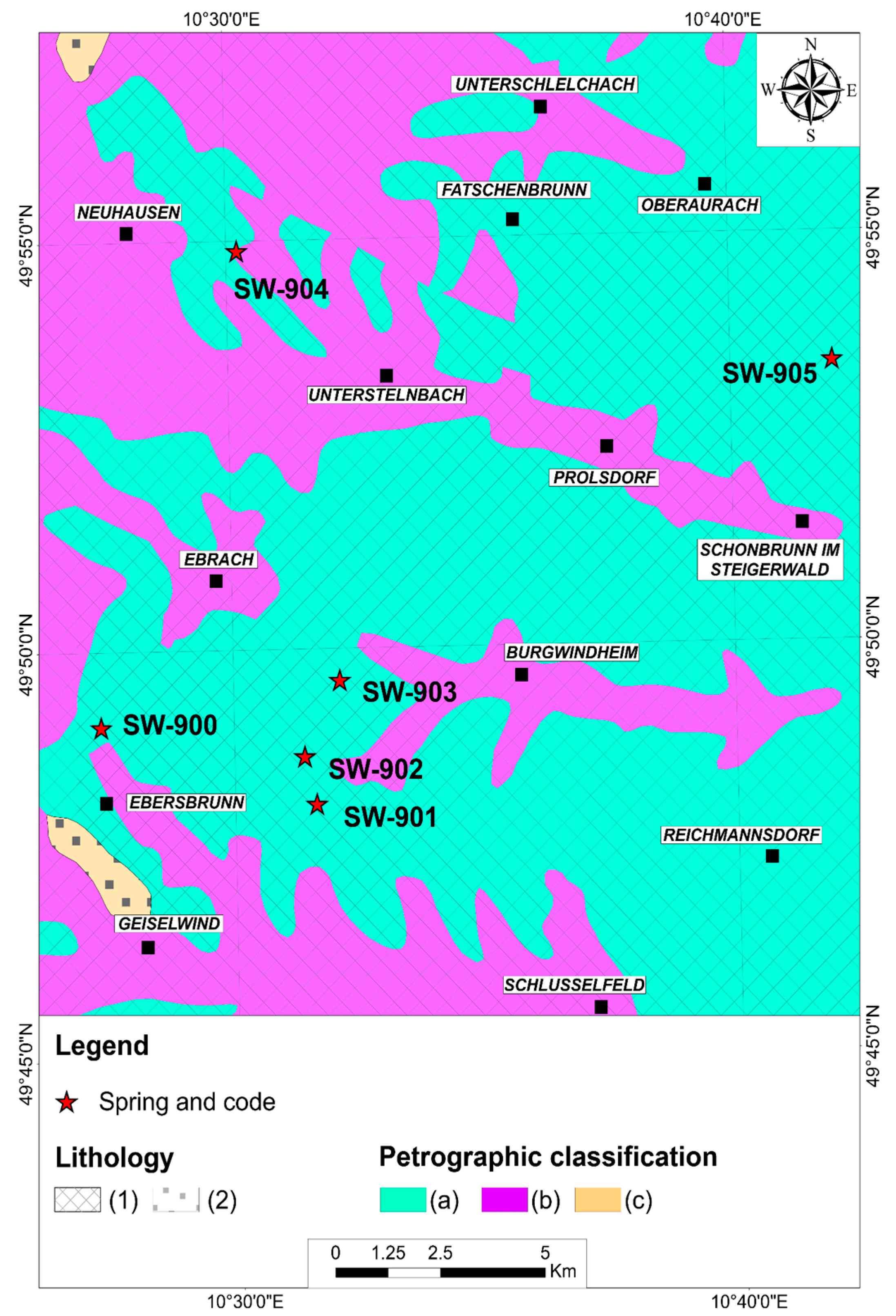
2.1. Study Areas
2.2. Field Work and Sampling
- Steinberg 39 (R891), Gangolfsberg 2 (R892), Kalktuff 1 (R893), Lahrbach 1 (R894), Mannsberg 9 (R895), Hoher Stern 28 (R896), Auersberg 133 (R897), Auersberg 119 (R898), and Buchenstrauch 14 (R899) for the Rhön.
- For the Steigerwald, 80 (SW900), 393 (SW901), 385 (SW902), 353 (SW903), 225 (SW904), and 528 (SW905). The fieldwork/sampling campaign was carried out from 27 June to 2 July 2021. The following parameters were measured: discharge, water temperature, conductivity, pH, and hydrochemistry (main ions and algal nutrients).
2.3. Geology and Hydrogeology
2.4. Hydrochemistry
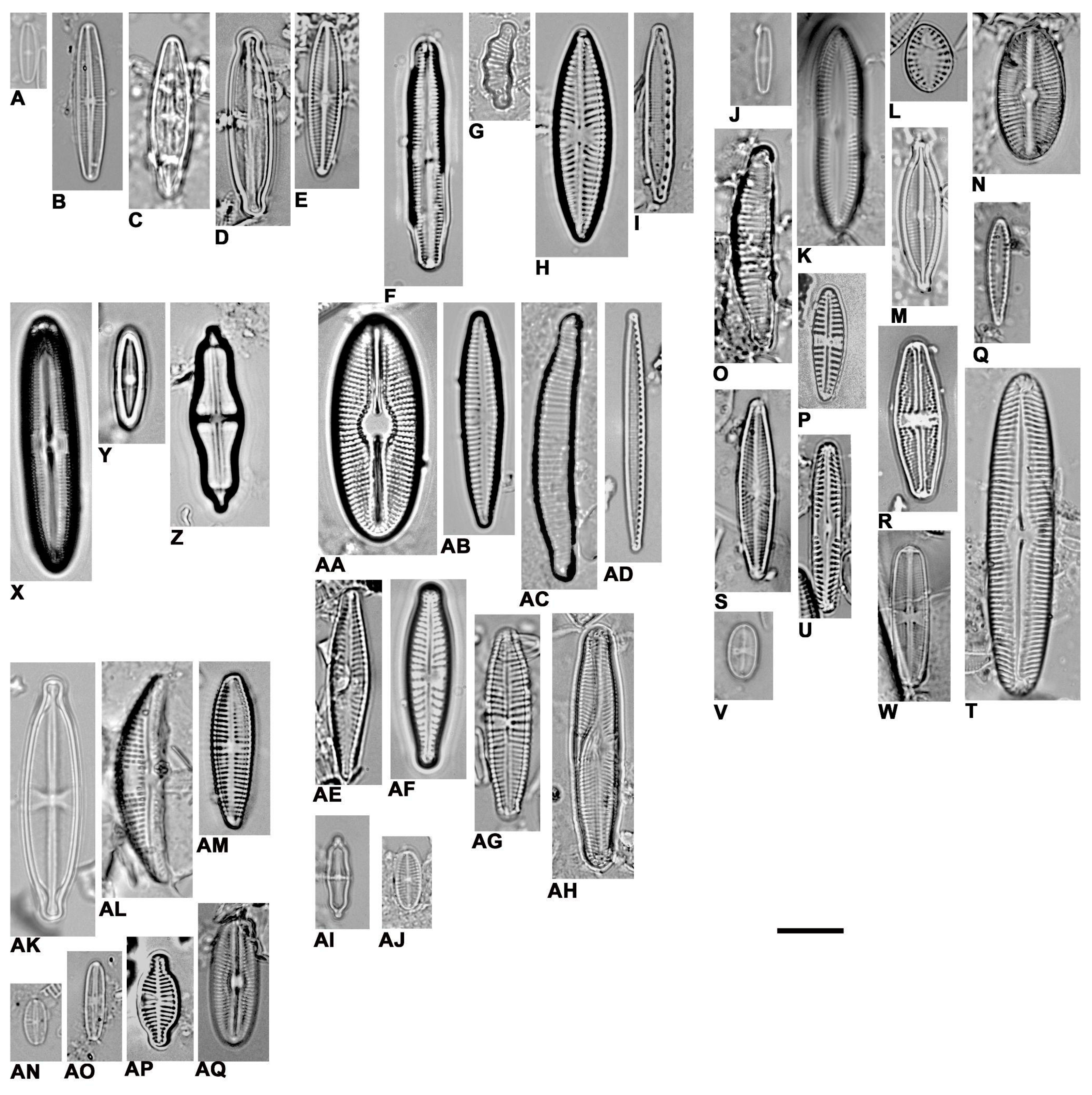
2.5. Diatom Sampling, Identification, and Quantification
2.6. Diatom Indicator and Metrics Data Validation with Invertebrates
2.7. Data Processing and Statistical Analyses
3. Results
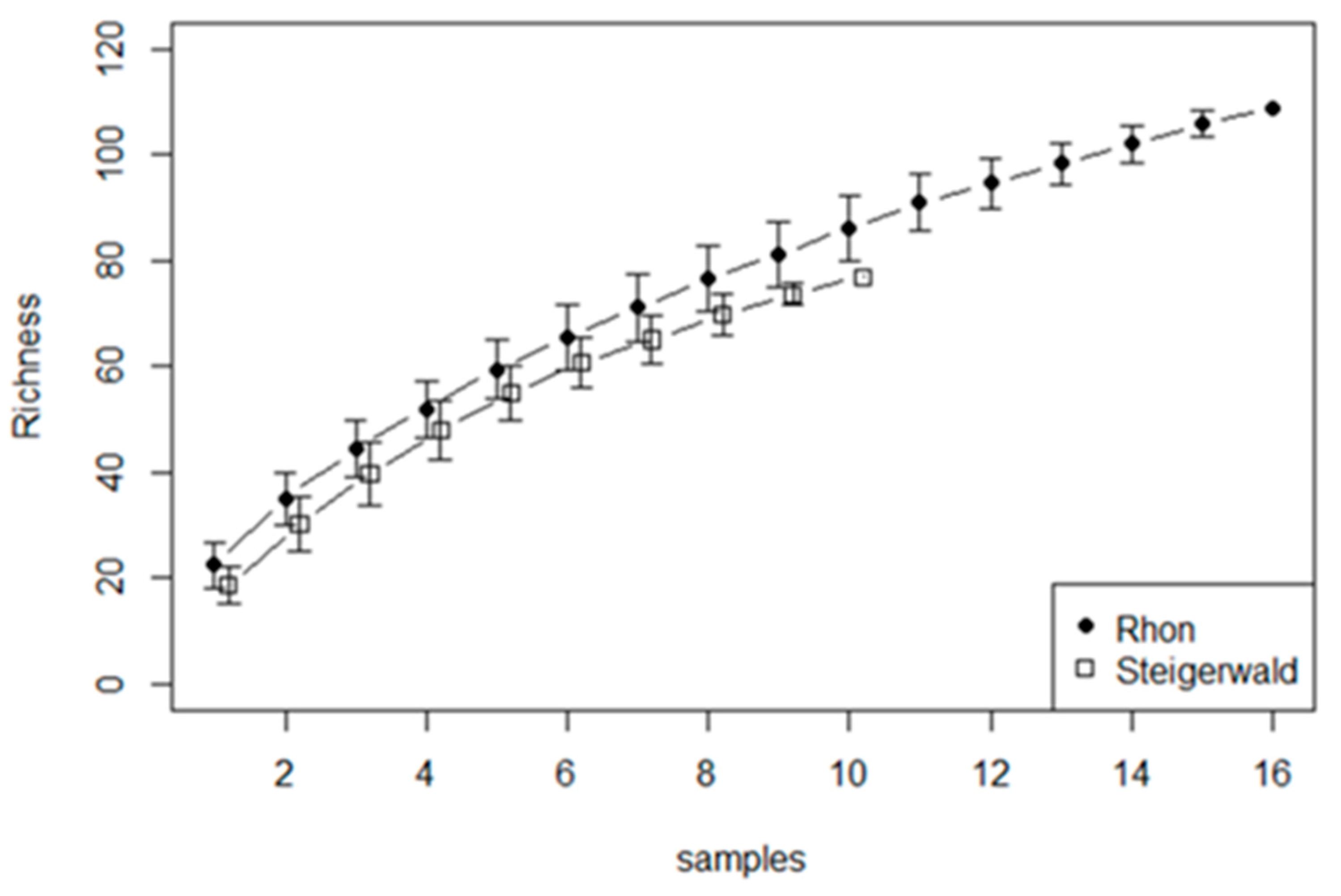
3.1. Samples Processed
3.2. The Diatoms Found in the Springs of the Two Nature Preserves
3.3. Representation of the Diatom Species Found in the Red List
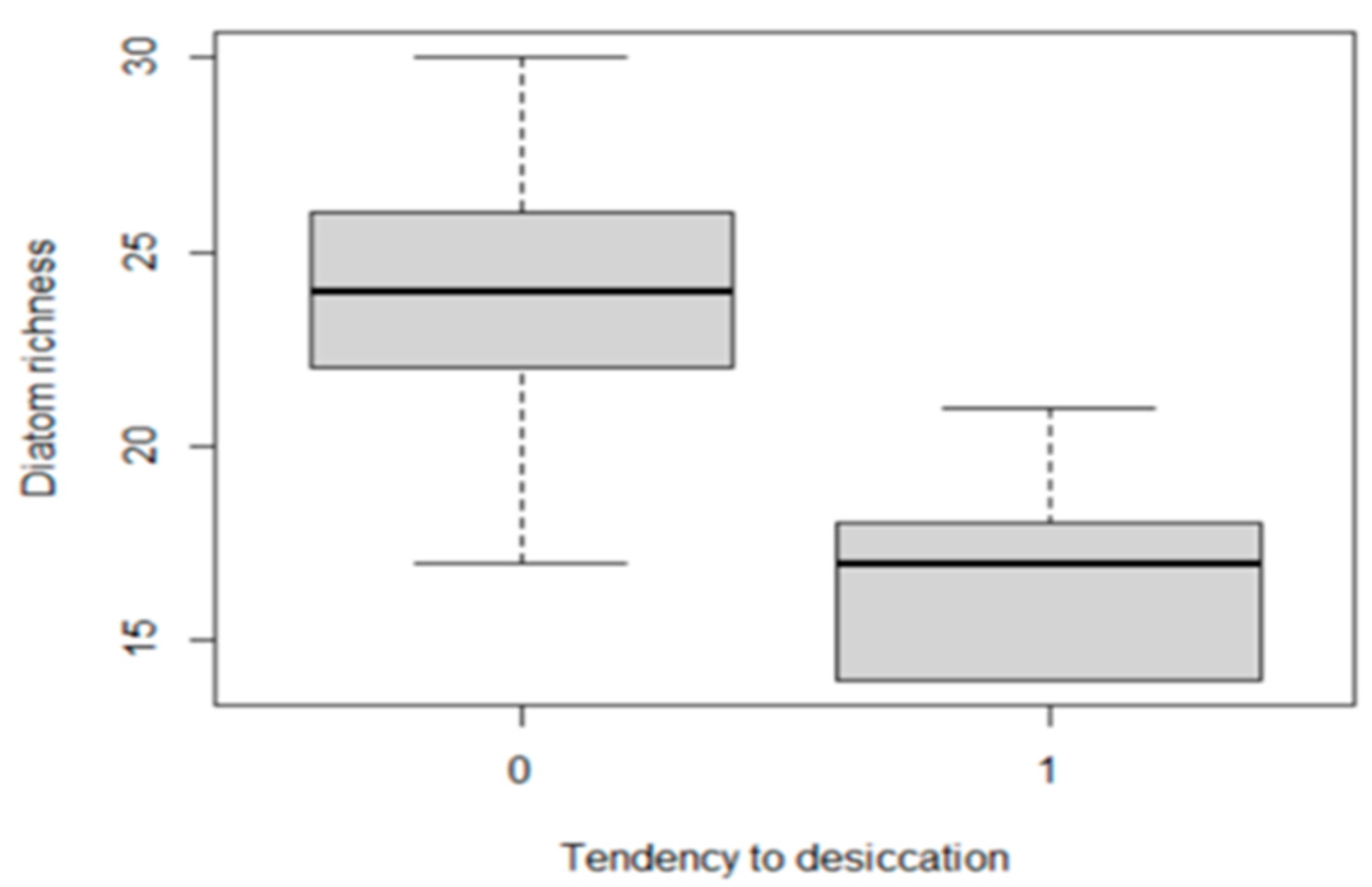
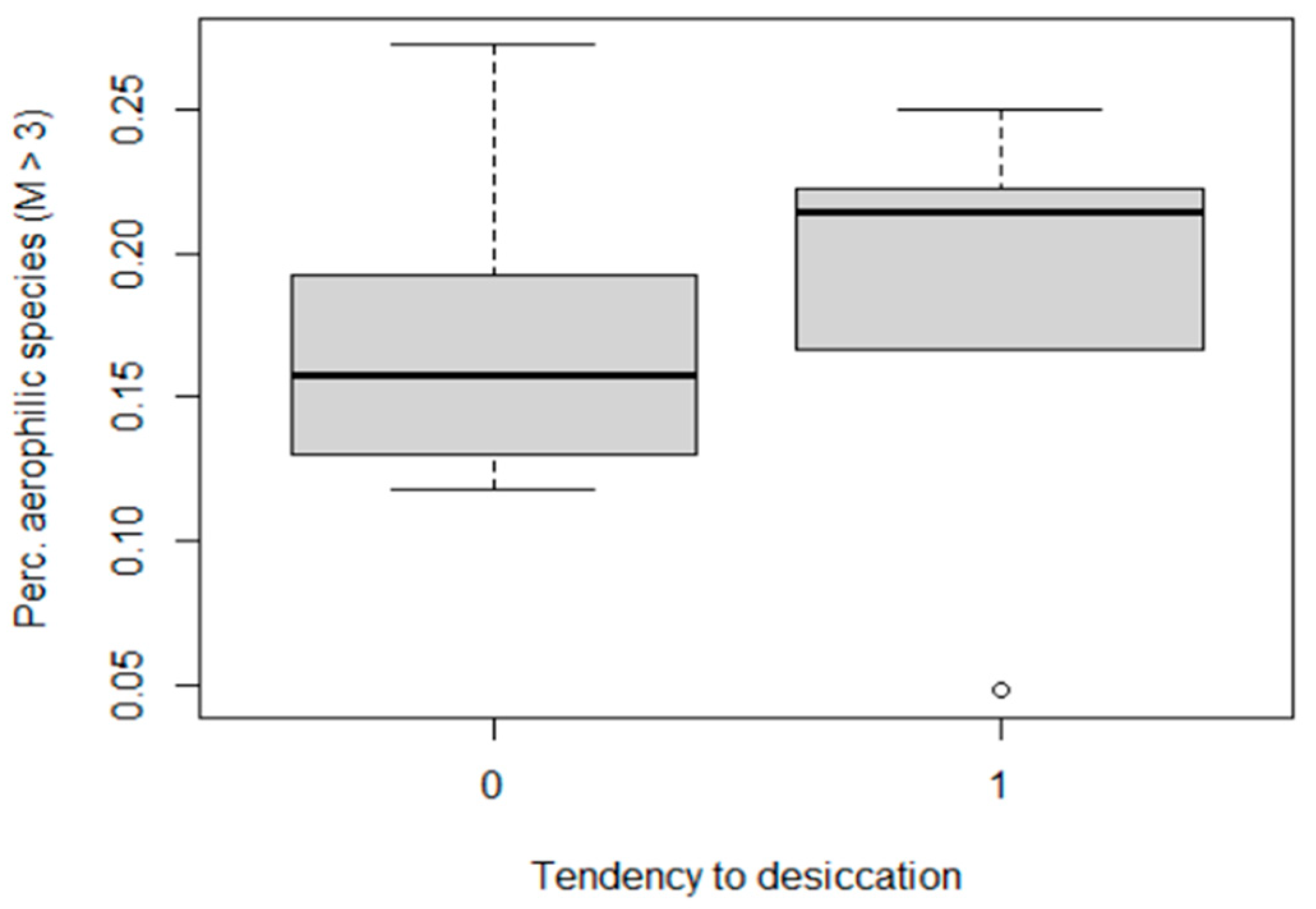
3.4. Ecological Attributes Associated with Discharge Fluctuation and Desiccation
3.5. Teratological Diatom Specimens
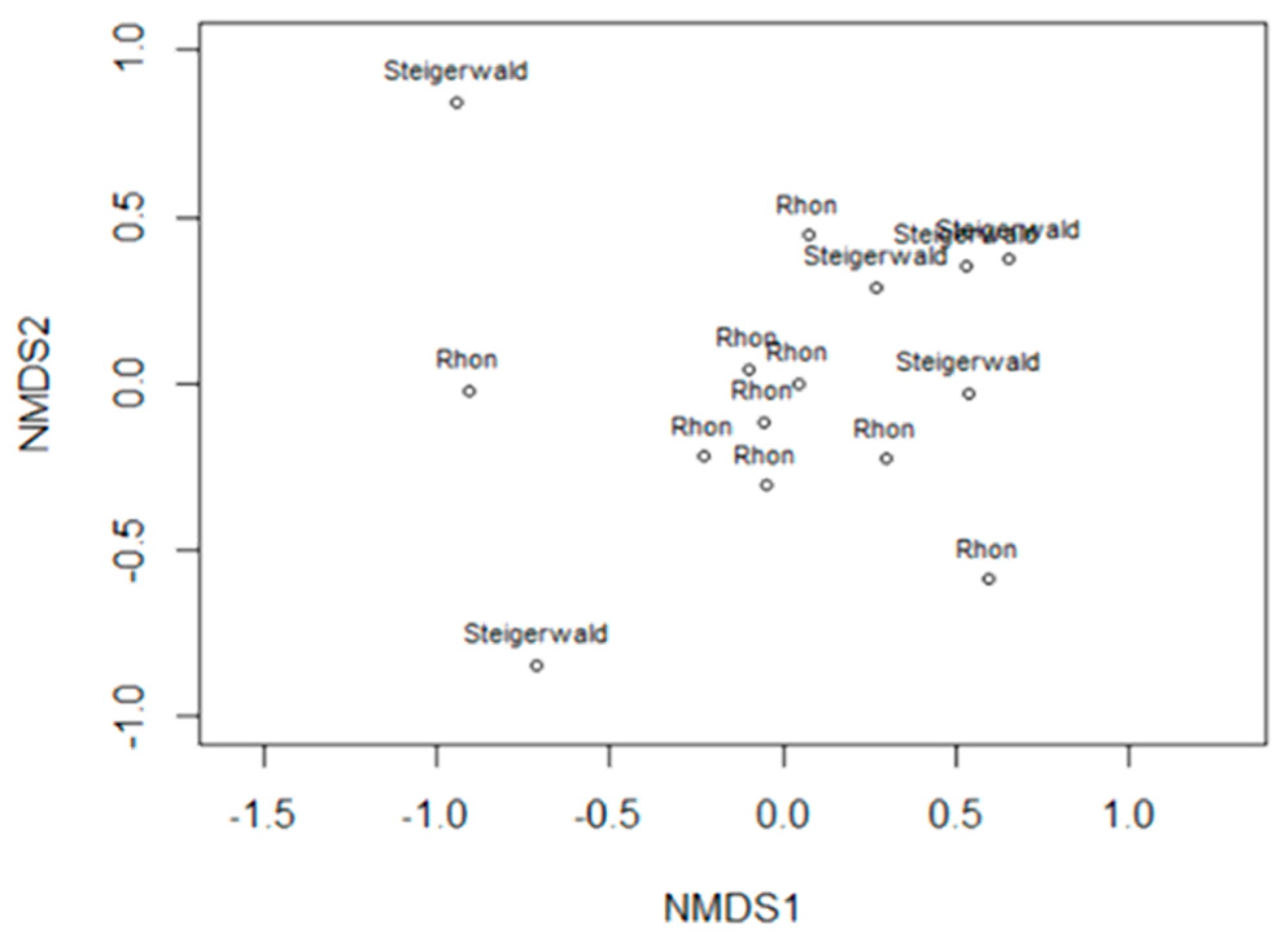
3.6. Results of Preliminary Statistical Analyses
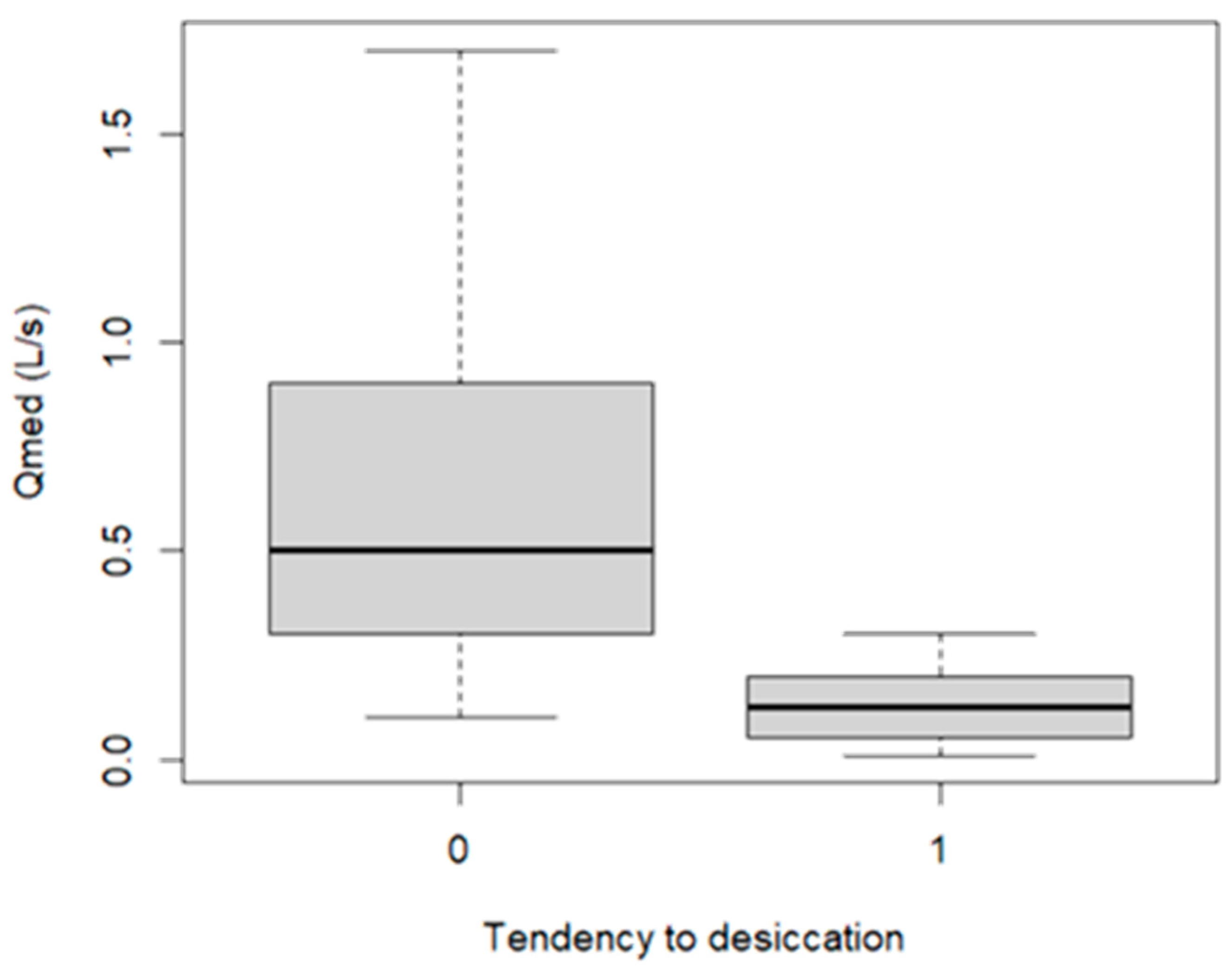
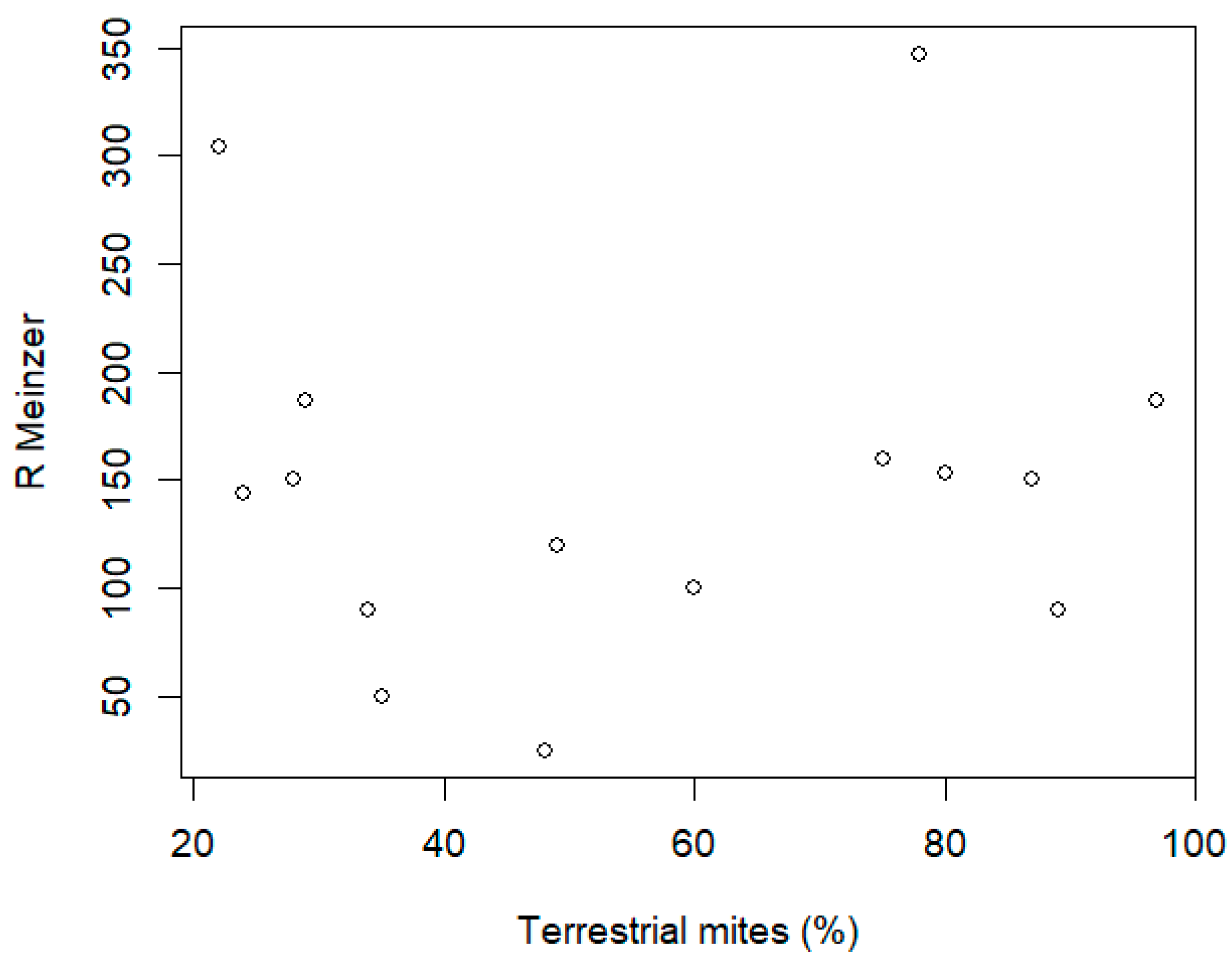
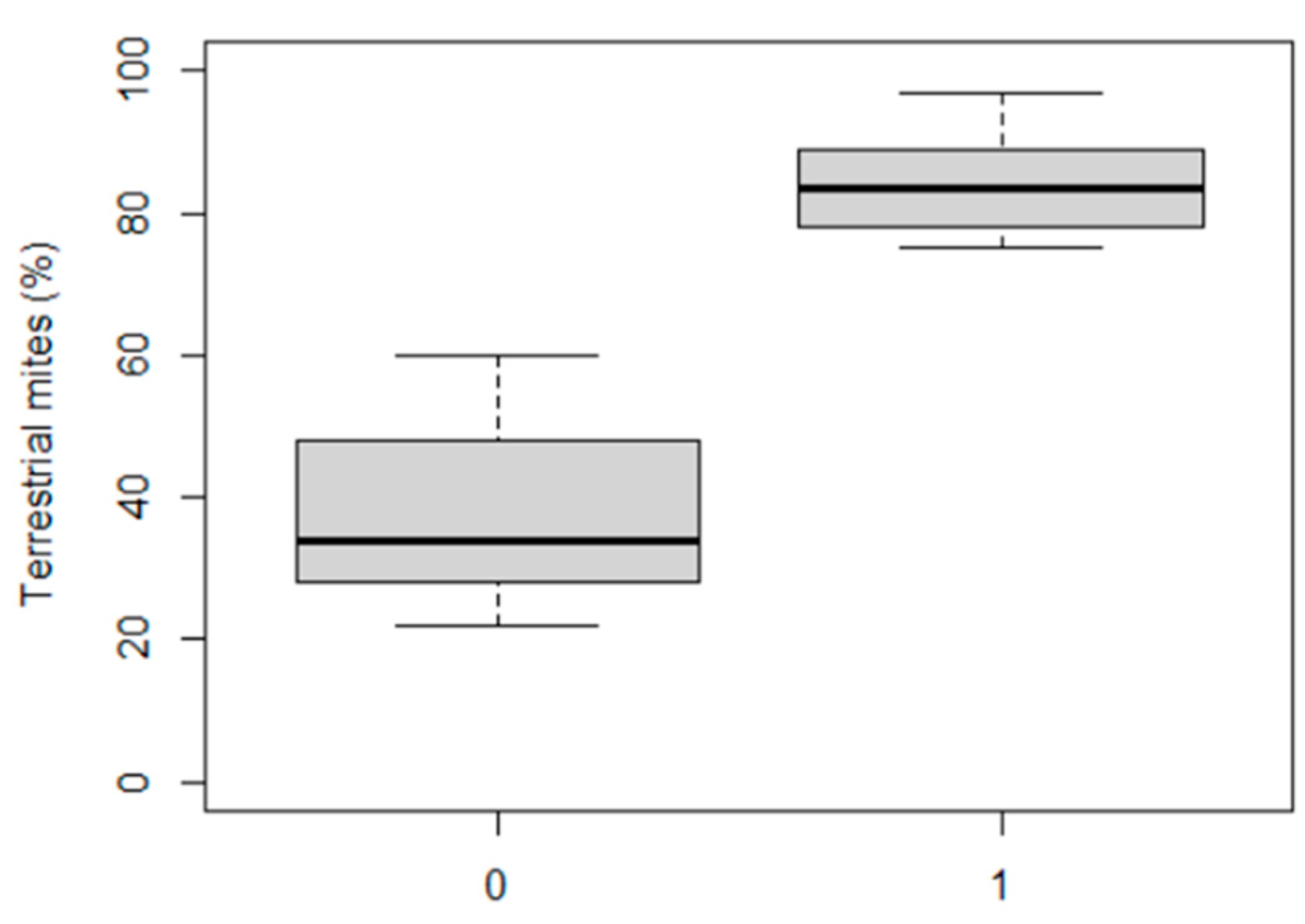
3.7. Faunal (Invertebrate) Desiccation Parameters to Validate Indicators/Metrics Based on Diatoms
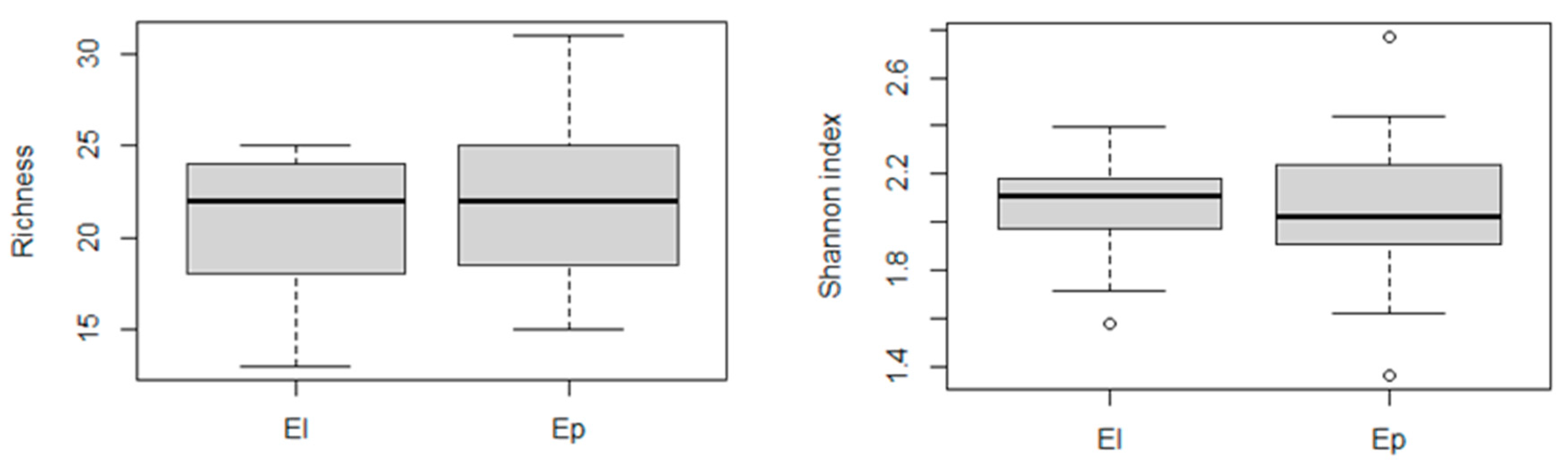
3.8. Diatom Species Richness in Stable and Fluctuating Discharge Springs
3.9. Diatom Red List Species in Stable and Fluctuating Discharge Springs
3.10. Aerial Diatom Species in Stable and Fluctuating Discharge Springs
3.11. Ecological Guilds in Stable and Fluctuating Discharge Springs
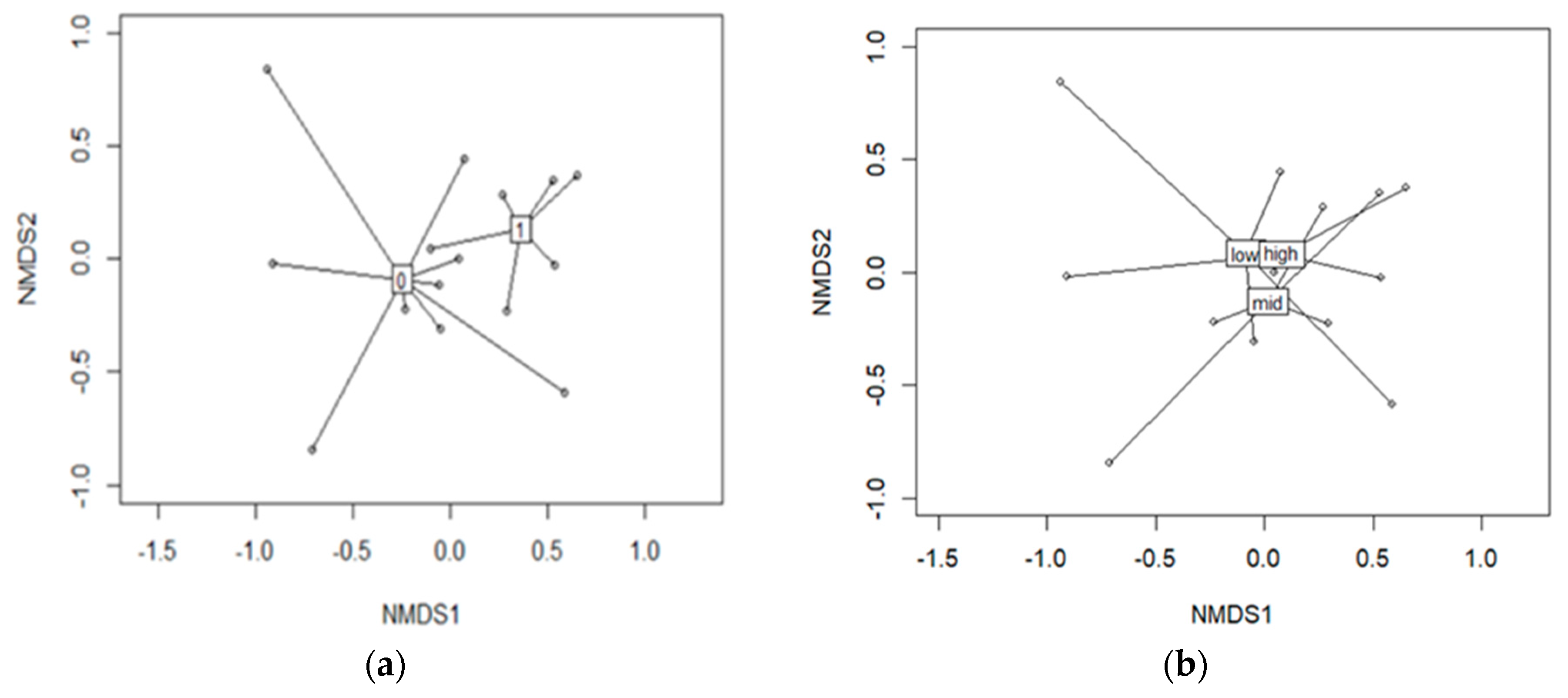
3.12. Diatom Communities in Stable and Fluctuating Discharge Springs
3.13. Diatom Indicators in Stable and Fluctuating Discharge Springs
4. Discussion
5. Conclusions
- -
- It is possible to use diatoms in spring ecosystems as indicators of water level / discharge fluctuations and desiccation probability.
- -
- Springs affected by discharge variability/desiccation have lower diatom species richness.
- -
- Springs affected by discharge variability/desiccation have distinct diatom communities.
- -
- Diatom indicators and metrics can be validated using other components of the spring biota, namely invertebrates.
- -
- Larger databases will be necessary to identify the most suitable diatom indicators of discharge variability/desiccation in springs.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Kamp, G. The hydrogeology of springs in relation to the biodiversity of spring fauna: A review. J. Kans. Entomol. 1995, 68, 4–17. [Google Scholar]
- Scarsbrook, M.; Barquín, J.; Gray, D. New Zeland coldwater springs and their biodiversity. Sci. Conserv. 2007, 278, 6. [Google Scholar]
- Glazier, D.S. Springs. In Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press Elsevier: Amsterdam, The Netherlands, 2009; Volume 1, pp. 734–755. [Google Scholar]
- Illies, J.; Botosaneanu, L. Problèmes et méthodes de la classification et de la zonation écologique des eaux courantes, considérées surtout du point de vue faunistique. Mitt. Int. Ver. Limnol. 1963, 12, 157. [Google Scholar] [CrossRef]
- Cantonati, M.; Füreder, L.; Gerecke, R.; Jüttner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 31, 463–480. [Google Scholar] [CrossRef]
- Cantonati, M.; Segadelli, S.; Spitale, S.; Gabrieli, J.; Gerecke, R.; Angeli, N.; De Nardo, M.T.; Ogata, K.; Wehr, J.D. Geological and hydrochemical prerequisites of unexpectedly high biodiversity in spring ecosystems at the landscape level. Sci. Total Environ. 2020, 740, 140157. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.G.; Vanormelingen, P. An Inordinate Fondness? The Number, Distributions, and Origins of Diatom Species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, D.G. The species concept in diatoms. Phycological. Reviews 18. Phycologia 1999, 38, 437–495. [Google Scholar] [CrossRef] [Green Version]
- Smol, J.P.; Stoermer, E.F. The Diatoms: Applications for the Environmental and Earth Sciences, 2nd ed.; Cambridge University Press: Cambridge, UK, 2010; pp. 1–667. [Google Scholar]
- Raven, J.A. The possible roles of algae in restricting the increase in atmospheric CO2 and global temperature. Eur. J. Appl. Physiol. 2017, 52, 506–522. [Google Scholar]
- Serôdio, J.; Lavaud, J. Diatoms and Their Ecological Importance. In Life Below Water; Encyclopedia of the UN Sustainable Development Goals; Leal Filho, W., Azul, A.M., Brandli, L., Lange Salvia, A., Wall, T., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–9. [Google Scholar]
- Hofmann, G.; Lange-Bertalot, H.; Werum, M.; Klee, R.; unter Mitarbeit von König, C.; Kusber, W.-H.; Metzeltin, D.; Reichardt, E. Rote Liste und Gesamtartenliste der limnischen Kieselalgen (Bacillariophyta) Deutschlands. In Rote Liste Gefährdeter Tiere, Pflanzen und Pilze Deutschlands; Band 7; Naturschutz und Biologische Vielfalt 70; Metzing, D., Hofbauer, N., Ludwig, G., Matzke Hajek, G., Eds.; Landwirtschaftsverlag: Münster, Germany, 2018; pp. 601–708. [Google Scholar]
- Cantonati, M.; Bilous, O.; Spitale, D.; Angeli, N.; Segadelli, S.; Bernabè, D.; Lichtenwöhrer, K.; Gerecke, R.; Saber, A.A. Diatoms from the Spring Ecosystems Selected for the Long-Term Monitoring of Climate-Change Effects in the Berchtesgaden National Park (Germany). Water 2022, 14, 381. [Google Scholar] [CrossRef]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar]
- Cantonati, M.; Angeli, N.; Bertuzzi, E.; Spitale, D.; Lange-Bertalot, H. Diatoms in springs of the Alps: Spring types, environmental determinants, and substratum. Freshw. Sci. 2012, 31, 499–524. [Google Scholar] [CrossRef]
- Cantonati, M.; Kelly, M.G.; Demartini, D.; Angeli, N.; Dörflinger, G.; Papatheodoulou, A.; Armanini, D. Overwhelming role of hydrology-related variables and river types in driving diatom species distribution and community assemblage in streams in Cyprus. Ecol. Indic. 2020, 117, 106690. [Google Scholar] [CrossRef]
- Falasco, E.; Piano, E.; Doretto, A.; Fenoglio, S.; Bona, F. Resilience of benthic diatom communities in Mediterranean streams: Role of endangered species. Mar. Freshw. Res. 2018, 70, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Artigas, J.; Fund, K.; Kirchen, S.; Morin, S.; Obst, U.; Romanı, A.M.; Sabater, S.; Schwartz, T. Patterns of biofilm formation in two streams from different bioclimatic regions: Analysis of microbial community structure and metabolism. Hydrobiologia 2012, 695, 83–96. [Google Scholar] [CrossRef]
- Calapez, A.R.; Elias, C.L.; Almeida, S.F.P.; Feio, M.J. Extreme drought effects and recovery patterns in the benthic communities of temperate streams. Limnetica 2014, 33, 281–296. [Google Scholar] [CrossRef]
- Piano, E.; Falasco, E.; Bona, F. How does water scarcity affect spatial and temporal patterns of diatom community assemblages in Mediterranean streams? Freshw. Biol. 2017, 62, 1276–1287. [Google Scholar] [CrossRef]
- Gerecke, R.; Cantonati, M.; Spitale, D.; Stur, E.; Wiedenbrug, S. The challenges of long-term ecological research in springs in the northern and southern Alps: Indicator groups, habitat diversity, and medium term change. J. Limnol. 2011, 70 (Suppl. S1), 168–187. [Google Scholar] [CrossRef]
- Deutscher Wetterdienst DWD: Climate Data Center (CDC), Annual Grids of Monthly Averaged Daily Air Temperature (2m) and Precipitation, Version v1.0. Available online: https://opendata.dwd.de/ (accessed on 22 July 2020).
- Verwaltungsstellen UNESCO-Biosphärenreservat Rhön in Bayern, Hessen und Thüringen: Neues Rahmenkonzept 2018 UNESCO-Biosphärenreservat Rhön Band I—Wo stehen wir? 2018, p. 541. Available online: https://www.biosphaerenreservat-rhoen.de/fileadmin/media/publikationen/Rahmenkonzept_Band_I.pdf (accessed on 2 August 2023).
- Die Waldklimastation Ebrach. 2023. Available online: https://www.lwf.bayern.de/boden-klima/umweltmonitoring/227196/index.php (accessed on 19 June 2023).
- Meinzer, O.E. Outline of Ground-Water Hydrology; Water Supply Paper; Department of the Interior, U.S. Geological Survey, Washington Government Printing Office: Washington, DC, USA, 1923; p. 494.
- Standard Methods for the Examination of Water and Wastewater, 20th ed.; APHA, AWWA & WEF, American Public Health Association: Washington, DC, USA, 2000; pp. 1–541.
- Cantonati, M.; Kelly, M.G.; Lange-Bertalot, H. (Eds.) Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Schmitten-Oberreifenberg, Germany, 2017; pp. 1–942. [Google Scholar]
- Guiry, M.D.; Guiry, G.M.; AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org (accessed on 20 June 2023).
- Kociolek, J.P.; Balasubramanian, K.; Blanco, S.; Coste, M.; Ector, L.; Liu, Y.; Kulikovskiy, M.; Lundholm, N.; Ludwig, T.; Potapova, M.; et al. DiatomBase. 2021. Available online: http://www.diatombase.org (accessed on 20 June 2023).
- Spaulding, S.A.; Bishop, I.W.; Edlund, M.B.; Lee, S.; Furey, P.; Jovanovska, E.; Potapova, M. Diatoms of North America. 2020. Available online: https://diatoms.org (accessed on 20 June 2023).
- Jüttner, I.; Carter, C.; Chudaev, D.; Cox, E.J.; Ector, L.; Jones, V.; Kelly, M.G.; Kennedy, B.; Mann, D.G.; Turner, J.A.; et al. Freshwater Diatom Flora of Britain and Ireland. Amgueddfa Cymru–National Museum Wales. 2021. Available online: https://naturalhistory.museumwales.ac.uk/diatoms (accessed on 20 June 2023).
- Lange-Bertalot, H. Rote Liste der Limnischen Kieselalgen (Bacillariophyceae) Deutschlands; Schriftenreihe für Vegetationskunde, Bundesamt fü Naturschutz: Bonn, Germany, 1996; Volume 28, pp. 633–677. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: https://www.R-project.org (accessed on 19 June 2023).
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan. 2019. Community Ecology Package. R Package Version 2.5–6. Available online: http://CRAN.Rproject.org/package=vegan (accessed on 19 June 2023).
- Lange-Bertalot, H.; Metzeltin, D. Indicators of Oligotrophy. 2. In Iconographia Diatomologica; Lange-Bertalot, H., Ed.; Koeltz Scientific Books: Königstein, Germany, 1996; pp. 1–390. [Google Scholar]
- Cantonati, M.; Virtanen, L.; Angeli, N.; Wojtal, A.; Gabrieli, J.; Falasco, E.; Lavoie, I.; Morin, S.; Marchetto, A.; Fortin, C.; et al. Achnanthidium minutissimum (Bacillariophyta) valve deformities as indicators of metal enrichment in diverse widely-distributed freshwater habitats. Sci. Total Environ. 2014, 475, 201–215. [Google Scholar] [CrossRef]
- Falasco, E.; Bona, F.; Badino, G.; Hoffmann, L.; Ector, L. Diatom teratological forms and environmental alterations: A review. Hydrobiologia 2009, 623, 1–35. [Google Scholar] [CrossRef]
- Falasco, E.; Ector, L.; Wetzel, C.E.; Badino, G.; Bona, F. Looking back, looking forward: A review of the new literature on diatom teratological forms (2010–2020). Hydrobiologia 2021, 848, 1675–1753. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Angeli, N.; Cantonati, M.; Spitale, D.; Lange-Bertalot, H. A comparison between diatom assemblages in two groups of carbonate, low-altitude springs with different levels of anthropogenic disturbances. J. Czech Phycol. Soc. 2010, 10, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Fritscher, J. Untersuchungen über Sulfi d–Schwefelquellen in Bayern. Ph.D. Thesis, University of Erlangen, Nürnberg, Germany, 2004; pp. 1–208. [Google Scholar]
- Cantonati, M.; Spitale, D. The role of environmental variables in structuring epiphytic and epilithic diatom assemblages in springs and streams of the Dolomiti Bellunesi National Park (South-Eastern Alps). Fundam. Appl. Limnol./Arch. Hydrobiol. 2009, 174, 117–133. [Google Scholar] [CrossRef]
- Taxböck, L.; Karger, D.N.; Kessler, M.; Spitale, D.; Cantonati, M. Diatom Species Richness in Swiss Springs Increases with Habitat Complexity and Elevation. Water 2020, 12, 449. [Google Scholar] [CrossRef] [Green Version]
- Passy, S.I. Diatom ecological guilds display distinct and predictable behavior along nutrient and disturbance gradients in running waters. Aquat. Bot. 2007, 86, 171–178. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spring Code | R-891 | R-892 | R-893 | R-894 | R-895 | R-896 | R-897 | R-898 | R-899 | SW-900 | SW-901 | SW-902 | SW-903 | SW-904 | SW-905 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Coordinates (WGS84) | 50.41383 N | 50.46764 N | 50.39182 N | 50.59260 N | 50.7092 N | 50.72227 N | 50.6046915 N | 50.6046092 N | 50.5081572 N | 49.818113N | 49.80184 N | 49.81156 N | 49.82695 N | 49.91474 N | 49.89039 N |
| 10.04189 E | 10.07887 E | 9.98014 E | 10.11219 E | 10.02535 E | 10.05276 E | 10.0149148 E | 10.0101308 E | 9.9913335 E | 10.45577 E | 10.52687 E | 10.5231 E | 10.53515 E | 10.50351 E | 10.70048 E | |
| Altitude, (m.a.s.l.) | 505 | 667 | 603 | 573 | 604 | 628 | 570 | 580 | 722 | 436 | 361 | 348 | 387 | 397 | 319 |
| Shading, (sc. 1–5) | 4–5 | 4–5 | 4–5 | 4 | 4–5 | 4–5 | 4–5 | 4–5 | 4–5 | 4 | 4–5 | 4–5 | 4–5 | 4–5 | 3 |
| Spring type | Helo | Helo | Rheo | Helo | Rheo | Helo | Helo | Rheo-helo | Helo | Rheo-helo | Rheo | Rheo | Rheo | Rheo | Rheo |
| Lithology | PCR (L,M,D) | MR (AB,BE,T) | PCR (L,M,D) | PCR (L,M,D) | MR (AB,BE,T) | PCR (L,M,D) | SS (SS,SI,M) | SS (SS,SI,M) | MR (AB,BE,T) | MCR (SS,MS,D) | MCR (SS,MS,D) | MCR (SS,MS,D) | MCR (SS,MS,D) | MCR (SS,MS,D) | MCR (SS,MS,D) |
| Disch., L s−1 | 0.6 (1.3/0.4) | 1.7 (5.2/0.04) | 0.3 (0.6/0.04) | 0.4 (0.4/0.3) | 0.2 (0.2/0.1) | 0.01 (0.02/0.005) | 0.1 (0.1/0.01) | 0.1 (0.1/0.01) | 0.9 (1.6/0.3) | 0.15 (0.02/0.3) | 0.5 (0.8/0.2) | 0.3 (0.6/0.14) | 0.2 (0.7/0.006) | 0.05 (0.1/0.02) | 0.9 (1.5/0.6) |
| MVID | 150 | 303.5 | 186.7 | 25 | 50 | 150 | 90 | 90 | 144.4 | 186.7 | 120 | 153.3 | 347 | 160 | 100 |
| Water T, °C | 8 (8.2/7.4) | 7.3 (7.5/7.1) | 7.8 (8.6/7.1) | 7.6 (7.6/7.6) | 7.6 (7.6/7.6) | N.R. | 8.4 (9.8/6.8) | 8.6 (10.1/7.3) | 8.1 (8.8/7.5) | 8.4 (10.7/6.9) | 9.2 (9.4/9) | 9.05 (9.5/8.7) | 9 (11.4/7.3) | 8.7 (9.4/8.3) | 9.2 (9.3/9.1) |
| Cond., µS cm−1 | 345 (433/287) | 189 (214/157) | 615 (673/559) | 315 (351/278) | 209 (215/200) | 379 (414/343) | 447 (471/427) | 573 (584/563) | 264 (293/236) | 88 (91/86) | 776 (856/714) | 744 (1022/90) | 91.5 (100/78) | 205 (353/85) | 745 (813/690) |
| pH | 7.7 (8/7.5) | 7.5 (7.7/7.4) | 7.7 (8.1/7.2) | 7.7 (7.9/7.4) | 8 (8.5/7.5) | 7.1 (7.2/7) | 8 (8.3/7.5) | 7.5 (7.9/7.3) | 7.8 (8/7.4) | 6.3 (7.1/5.8) | 7.7 (7.8/7.5) | 7.5 (8/6.1) | 6.7 (7.5/6.2) | 7.4 (8.1/6.8) | 7.7 (7.9/7.4) |
| Mg2+, mg L−1 | 8.9 | 7.8 | 4.1 | 6.9 | 10.4 | 5.7 | 6.5 | 7 | 3.9 | 2.9 | 38.6 | 15.6 | 2.3 | 3.8 | 46.1 |
| Ca2+, mg L−1 | 47.1 | 18 | 96.5 | 33.6 | 17.3 | 43.4 | 81 | 110 | 39.8 | 4 | 94.7 | 23 | 3.8 | 6.4 | 92 |
| Na+, mg L−1 | 2.1 | 2.5 | 24.6 | 2.1 | 5.7 | 2.2 | 2.3 | 2.2 | 2.2 | 4 | 6.1 | 4.9 | 2.4 | 3.3 | 4.9 |
| K+, mg L−1 | 0.6 | 0.6 | 0.7 | <0.5 | 1.1 | <0.5 | 0.7 | 1.9 | <0.5 | 0.8 | 1.6 | 2.1 | 1.3 | 1.3 | 1.3 |
| Cl−, mg L−1 | 2.3 | 2.3 | 48.5 | 2.6 | 3.9 | 2.9 | 2.7 | 3.2 | 2.2 | 4 | 9.4 | 3.1 | 2.1 | 2.9 | 5.6 |
| NO3−, mg L−1 | 1.7 | 1.7 | 2.6 | 5.5 | 11.5 | 16.2 | 5 | 4.2 | 2.9 | 9.4 | 15.9 | 4 | 2.3 | ||
| SO42−, mg L−1 | 8.2 | 8.2 | 6.2 | 6.1 | 14.5 | 10.7 | 8.3 | 8.1 | 4.9 | 11 | 140 | 41.1 | 4.5 | ||
| PO43−, mg L−1 | 0.069 | 0.2 | <0.03 | 0.093 | 0.14 | 0.04 | 0.069 | 0.21 | 0.14 | 0.088 | 0.038 | 0.092 | 0.069 | ||
| TP, mg L−1 | 0.028 | 0.071 | <0.010 | 0.036 | 0.047 | 0.023 | 0.039 | 0.071 | 0.047 | - | - | - | - |
| Taxa | M.A. | N.O. | RL18 | RL96 | Evolution | E c o l o g y | M | Ep | El | Ma |
|---|---|---|---|---|---|---|---|---|---|---|
| Achnanthidium affine (Grunow) Czarn. | 7.3% | 8 | * | * | = | ? | - | x | x | xx |
| Achnanthidium dolomiticum Cantonati et Lange-Bert. | 19.3% | 20 | 2 | - | ? | xxx | xx | x | ||
| Achnanthidium gracillimum (Meister) Lange-Bert. in Krammer et Lange-Bert. | 0.50% | 1 | 2 | 3 | - | oc | - | x | ||
| Achnanthidium lineare W.Sm. | 51.8% | 24 | G | ♦° | eu | - | xx | xxx | xx | |
| Achnanthidium minutissimum (Kütz.) Czarn. | 24.00% | 14 | * | **° | = | ? | 3 | xx | x | xx |
| Achnanthidium pfisteri Lange-Bert. | 7.50% | 7 | D | - | ? | xx | xx | |||
| Achnantidium polonicum Van de Vijver, Wojtal, Morales et L.Ector | 2.00% | 1 | x | |||||||
| Achnanthidium subatomus (Hust.) Lange-Bert. | 1.00% | 1 | V | * | – | od | 1 | x | ||
| Adlafia bryophila (Petersen) Lange-Bert. in Moser et al. | 3.00% | 6 | * | V | + | ? | 5 | x | x | x |
| Adlafia langebertalotii Monnier et L.Ector | 0.25% | 2 | * | – | ? | x | x | |||
| Adlafia minuscula (Grunow) Lange-Bert. in Lange-Bert. et Genkal | 2.00% | 10 | * | * | = | ? | 4 | x | x | x |
| Amphora indistincta Levkov | 3.50% | 3 | * | - | ? | x | x | |||
| Amphora lange-bertalotii var. tenuis Levkov et Metzeltin | 0.25% | 2 | D | - | o | x | ||||
| Amphora micra Levkov | 49.00% | 23 | xx | xx | xx | |||||
| Amphora pediculus (Kütz.) Grunow in A.Schmidt et al. | 39.3% | 15 | * | ** | = | ? | 3 | xx | xx | xx |
| Brachysira calcicola var. calcicola Lange-Bert. | 0.25% | 1 | 2 | ♦° | oc | x | ||||
| Caloneis constans E.Reichardt | 4.25% | 2 | R | – | oc | x | ||||
| Caloneis fontinalis (Grunow) Lange-Bert. et Reichardt | 15.3% | 7 | * | – | ? | x | x | |||
| Caloneis lancettula (Schulz-Danzing) Lange-Bert. et Witkowski | 43.5% | 9 | * | ♦° | eu | x | x | |||
| Caloneis lauta J.R.Carter | 0.25% | 1 | G | G | = | o | 1 | x | ||
| Caloneis tenuis (W.Greg.) Krammer | 0.50% | 2 | 3 | G | o | 4 | x | x | ||
| Caloneis vasileyevae Lange-Bert., Genkal et Vekhov | 2.00% | 2 | R | - | ? | x | ||||
| Cymbella compacta Østrup | 0.75% | 1 | * | * | = | eu | x | |||
| Cymbopleura austriaca (Grunow) Krammer | 0.50% | 1 | 2 | V | – | ae/oc | x | |||
| Cocconeis euglypta Ehrenb. | 4.80% | 15 | * | ** | = | ? | 2 | xx | x | x |
| Cocconeis lineata Ehrenb. | 9.50% | 11 | * | ** | = | ? | 2 | x | x | x |
| Cocconeis pseudolineata (Geitler) Lange-Bert. | 7.30% | 8 | * | D | ? | 2 | x | x | ||
| Cocconeis pseudothumensis E.Reichardt | 1.00% | 2 | G | 3 | oc | 3 | x | x | ||
| Craticula minusculoides (Hust.) Lange-Bert. | 0.50% | 1 | * | * | = | eu | 2 | x | ||
| Craticula vixnegligenda Lange-Bert. | 0.50% | 1 | G | R | - | o | x | |||
| Denticula tenuis Kütz. | 1.80% | 4 | * | * | = | o | 3 | x | x | |
| Diploneis fontanella Lange-Bert. | 1.30% | 2 | G | - | o | x | ||||
| Diploneis krammeri Lange-Bert. et E.Reichardt | 0.50% | 2 | V | ♦° | oc | x | ||||
| Diploneis separanda Lange-Bert. | 1.00% | 3 | D | ♦° | oc | x | x | |||
| Encyonopsis falaisensis (Grunow) Krammer | 1.00% | 1 | G | G | = | o | 3 | x | ||
| Encyonopsis fonticola (Hust.) Krammer | 0.25% | 1 | x | |||||||
| Eunotia boreoalpina Lange-Bert. et Nörpel-Schempp in Lange-Bert. et Metzeltin | 0.50% | 1 | G | - | od | 2 | x | |||
| Eunotia minor (Kütz.) Grunow | 14.2% | 5 | V | * | – | ? | - | x | xxx | |
| Eunotia paratridentula Lange-Bert. et Kulikovskiy | 1.50% | 1 | 3 | * | – | od | x | |||
| Eunotia soleirolii (Kütz.) Rabenh. | 7.25% | 3 | G | G | = | od | 1 | x | x | |
| Frustulia vulgaris (Thwaites) De Toni | 0.25% | 2 | * | ** | = | ? | 3 | x | ||
| Gogorevia exilis (Grunow) Czarn. | 0.25% | 1 | 2 | x | ||||||
| Gomphonema angustum C.Agardh | 18.00% | 12 | G | V | - | oc | - | xx | x | x |
| Gomphonema cuneolus E.Reichardt | 10.00% | 1 | G | - | oc | x | ||||
| Gomphonema cymbelliclinum E.Reichardt et Lange-Bert. | 3.00% | 1 | * | - | ? | x | ||||
| Gomphonema elegantissimum E.Reichardt et Lange-Bert. | 22.3% | 4 | * | - | oc | x | x | |||
| Gomphonema exilissimum (Grunow) Lange-Bert. et Reichardt | 1.00% | 1 | V | V | = | od | - | x | ||
| Gomphonema extentum E.Reichardt et Lange-Bert. | 4.3% | 4 | * | ** | = | hal | x | x | xx | |
| Gomphonema hebridense W.Greg. | 0.50% | 1 | V | V | = | ? | x | |||
| Gomphonema innocens E.Reichardt | 0.75% | 2 | * | - | ? | x | ||||
| Gomphonema micropus Kütz. | 4.00% | 10 | * | ♦° | ? | 3 | x | x | x | |
| Gomphonema minutum (C.Agardh) C.Agardh | 3.25% | 1 | * | ** | = | eu | - | x | ||
| Gomphonema productum (Grunow) Lange-Bert. et Reichardt in Lange-Bert. | 2.00% | 6 | V | V | = | ? | x | x | ||
| Gomphonema pseudotenellum Lange-Bert. in Krammer et Lange-Bert. | 1.50% | 2 | G | 3 | o | x | x | |||
| Gomphonema pumilum (Grunow) E.Reichardt et Lange-Bert. | 9,75% | 4 | * | ♦° | ? | - | x | |||
| Gomphonema sarcophagus W.Greg. | 5.00% | 3 | V | V | = | ? | 3 | x | x | |
| Gomphonema subclavatum (Grunow) Grunow | 3.30% | 6 | * | ♦° | ? | 3 | x | x | x | |
| Gomphonema utae Lange-Bert. et E.Reichardt in Reichardt | 16.8% | 4 | * | D | ? | x | x | xx | ||
| Gomphonema varioreduncum Jüttner, L.Ector, E.Reichardt, Van de Vijver et E.J.Cox in Jüttner et al. | 0.50% | 2 | D | – | od | x | ||||
| Grunowia sinuata (Thwaites) Rabenh. | 0.50% | 2 | x | x | ||||||
| Halamphora normanii (Rabenh.) Levkov | 7.00% | 5 | D | ♦° | ae | 4 | x | |||
| Hantzschia calcifuga E.Reichardt et Lange-Bert. | 1.25% | 1 | D | - | od | x | ||||
| Humidophila brekkaensis (Petersen) R.LLowe et al. | 1.00% | 6 | * | V | + | ae | x | x | x | |
| Humidophila contenta (Grunow) R.LLowe, Kociolek, Johansen, Van de Vijver, Lange-Bert. et Kopalová | 12.2% | 12 | D | ♦° | ae | 4 | xx | x | ||
| Humidophila perpusilla (Grunow) R.L.Lowe, Kociolek, J.R.Johans., Van de Vijver, Lange-Bert. et Kopalová | 53.00% | 22 | * | ** | = | ae/o | 5 | xxx | xx | x |
| Karayevia clevei (Grunow) Bukht. | 6.80% | 3 | * | * | = | eu | 1 | x | x | |
| Kobayasiella subtilissima (Cleve) Lange-Bert. | 0.75% | 1 | 2 | 2 | = | od | 3 | x | ||
| Luticola acidoclinata Lange-Bert. | 0.50% | 1 | G | D | od | x | ||||
| Luticola goeppertiana (Bleisch) D.G.Mann | 0.50% | 1 | * | ♦° | eu | 3 | x | |||
| Mayamaea fossalis var. fossalis (Krasske) Lange-Bert. | 2.50% | 4 | * | * | = | ? | 4 | x | x | x |
| Mayamaea permitis (Hust.) K.Bruder et Medlin | 5.75% | 6 | * | ** | = | eu | 3 | x | ||
| Meridion circulare (Grév.) C.Agardh | 27.3% | 18 | * | ** | = | ? | 1 | x | xx | xx |
| Meridion constrictum Ralfs | 1.50% | 1 | * | ** | = | o | 2 | x | x | |
| Meridion infirmatum E.Reichardt | 0.25% | 2 | x | |||||||
| Microfissurata paludosa Cantonati et Lange-Bert. | 0.25% | 1 | x | |||||||
| Navicula antonii Lange-Bert. in Rumrich et al. | 2.25% | 1 | * | ** | = | eu | - | x | ||
| Navicula cryptocephala Kütz. | 0.50% | 3 | * | ** ᷉ | = | eu | 2 | x | x | x |
| Navicula cryptotenelloides Lange-Bert. | 0.25% | 1 | * | ** | = | ? | x | |||
| Navicula exilis Kütz. | 0.75% | 3 | 3 | G | od | - | x | x | ||
| Navicula gregaria Donkin | 1.75% | 1 | * | ** | = | ? | 5 | x | ||
| Navicula moenofranconica Lange-Bert. | 0.75% | 1 | 3 | 3 | = | ? | x | |||
| Navicula notha J.H.Wallace | 2.25% | 3 | 2 | G | od | 2 | x | x | ||
| Navicula tenelloides Hust. | 1.00% | 4 | * | * | = | eu | 4 | x | x | |
| Navicula tripunctata (O.F.Müll.) Bory | 1.25% | 4 | * | ** | = | eu | 3 | x | x | x |
| Navicula wygaschii Lange-Bert. | 0.25% | 1 | G | - | oc | x | ||||
| Nitzschia acidoclinata Lange-Bert. | 4.25% | 3 | V | * | – | ? | 3 | x | x | |
| Nitzschia alpina Hust. | 1.00% | 3 | 3 | G | o | - | x | x | ||
| Nitzschia cf. communis Rabenh. | 0.25% | 1 | * | ** | = | eu | 5 | x | ||
| Nitzschia dissipata (Kütz.) Grunow | 1.50% | 3 | * | ** | = | eu | 3 | x | x | |
| Nitzschia fonticola Grunow in Cleve et Möller | 0.75% | 3 | * | ** | = | eu | 1 | x | x | |
| Nitzschia hantzschiana Rabenh. | 2.25% | 9 | G | * | – | ? | 4 | x | x | |
| Nitzschia intermedia Hantzsch in Cleve et Grunow | 2.00% | 2 | * | * | = | eu | 1 | x | ||
| Nitzschia linearis (C.Agardh) W.Sm. | 2.50% | 10 | * | ** | = | eu | 3 | x | x | x |
| Nitzschia perminuta (Grunow) H.Peragallo | 1.00% | 5 | * | * | = | ? | 3 | x | x | x |
| Nitzschia pura Hust. | 0.25% | 1 | * | *° | = | o | - | x | ||
| Odontidium hyemale (Roth) Kütz. | 1.00% | 1 | 2 | x | ||||||
| Odontidium mesodon (Ehrenb.) Ralfs | 3.00% | 5 | 2 | x | x | x | ||||
| Orthoseira roeseana (Rabenh.) O’Meara | 2.00% | 1 | D | V | ae | x | ||||
| Pinnularia microstauron (Ehrenb.) Cleve | 0.25% | 1 | V | V | = | od | 7 | x | ||
| Pinnularia obscura Krasske | 1.00% | 1 | * | ** | = | ae | 4 | x | ||
| Pinnularia perirrorata Krammer | 1.75% | 3 | * | – | od | x | x | |||
| Pinnularia rupestris Hantzsch | 0.25% | 1 | G | ♦° ᷉ | od | 1 | x | |||
| Pinnularia schoenfelderi Krammer | 1.00% | 1 | G | G | = | od | x | |||
| Planothidium dubium (Grunow) Round et Bukht. | 0.50% | 1 | * | * | = | eu | x | |||
| Planothidium frequentissimum (Lange-Bert.) Lange-Bert. | 16.2% | 27 | * | ** | = | eu | - | xxx | xx | xxx |
| Planothidium lanceolatum (Bréb. ex Kütz.) Lange-Bert. | 76.1% | 35 | * | ** | = | ? | 3 | xxx | xxx | xxx |
| Planothidium reichardtii Lange-Bert. et Werum in Werum et Lange-Bert. | 5.00% | 9 | D | - | ? | x | x | x | ||
| Planothidium rostratoholoarticum Lange-Bert. et Bąk | 0.50% | 2 | * | D | eu | 3 | x | x | ||
| Psammothidium grischunum (Wuthrich) Bukht. et Round | 31.00% | 14 | V | * | - | ? | - | xx | x | xx |
| Psammothidium subatomoides (Hust.) Bukht. et Round | 12.7% | 8 | G | G | = | od | 1 | x | x | x |
| Reimeria sinuata (W.Greg.) Kociolek et Stoermer | 2.50% | 2 | * | ♦° | ? | 3 | x | |||
| Rhoicosphenia abbreviata (C.Agardh) Lange-Bert. | 6.50% | 3 | * | ** | = | eu | 2 | x | ||
| Rhoicosphenia tenuis Levkov et T.Nakov | 11.2% | 1 | x | |||||||
| Sellaphora atomoides (Grunow) Wetzel et Van de Vijver | 4.75% | 8 | x | x | ||||||
| Sellaphora nigri (De Not.) C.E.Wetzel et L.Ector in C.E.Wetzel et al. | 27.7% | 28 | 3 | xxx | xxx | xxx | ||||
| Sellaphora pupula (Kütz.) Mereschk. | 0.25% | 1 | D | **° | eu | 4 | x | |||
| Sellaphora pseudopupula (Krasske) Lange-Bert. | 1.50% | 1 | G | G | = | od | x | |||
| Sellaphora saugerresii (Desm.) C.E.Wetzel et D.G. Mann | 5.50% | 11 | 3 | xx | x | x | ||||
| Sellaphora seminulum (Grunow) D.G.Mann | 6.00% | 4 | * | ** | = | eu | 3 | x | x | x |
| Sellaphora stroemii (Hust.) D.G.Mann | 1.30% | 2 | 2 | 3 | - | oc | 4 | x | x | |
| Stauroneis kriegeri R.M.Patrick | 0.50% | 1 | * | * | = | ? | 4 | x | ||
| Stauroneis parathermicola Lange-Bert. | 0.50% | 1 | * | – | ? | x | ||||
| Stauroneis separanda Lange-Bert. et Werum | 0.50% | 1 | V | - | oc | x | ||||
| Stauroneis silvahassiaca Lange-Bert. et Werum | 0.25% | 1 | D | – | od | x | ||||
| Stauroneis smithii Grunow | 0.50% | 3 | R | ♦° | ? | 3 | x | x | ||
| Stauroneis thermicola (J.B.Petersen) Lund | 2.50% | 5 | * | * | = | ae | 4 | x | x | |
| Staurosirella pinnata (Ehrenb.) D.M.Williams et Round | 26.5% | 3 | * | ** | = | ? | 7 | x | x | |
| Surirella terricola Lange-Bert. et Alles | 0.75% | 2 | D | * | ae | x | x |
| Spring Code | % Terrestrial Mites | % Zoobenthos Species | Desiccation Probability |
|---|---|---|---|
| R-891 | 28 | 54 | 0 |
| R-892 | 22 | 60 | 0 |
| R-893 | 29 | 41 | 0 |
| R-894 | 48 | 47 | 0 |
| R-895 | 35 | 58 | 0 |
| R-896 | 87 | 54 | 1 |
| R-897 | 34 | 39 | 0 |
| R-898 | 89 | 38 | 1 |
| R-899 | 24 | 51 | 0 |
| SW-900 | 97 | 47 | 1 |
| SW-901 | 49 | 60 | 0 |
| SW-902 | 80 | 54 | 1 |
| SW-903 | 78 | 55 | 1 |
| SW-904 | 75 | 51 | 1 |
| SW-905 | 60 | 54 | 0 |
| Spring Type 0 (Perennial) | Type 1 | Total | ||
|---|---|---|---|---|
| High profile (N of sp) | Observed | 7 | 9 | 16 |
| Expected | 10.8 | 5.2 | ||
| Low profile | Observed | 84 | 40 | 124 |
| Expected | 83.4 | 40.6 | ||
| Motile | Observed | 71 | 30 | 101 |
| Expected | 69.9 | 33.1 | ||
| Total Observed | 162 | 79 | 241 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantonati, M.; Casoria, C.; Gerecke, R.; Bilous, O.P.; Maisto, G.; Segadelli, S.; Spitale, D.; Steinbauer, A.; Vogel, S.; Saber, A.A. Diatom Indicators of Fluctuating/Intermittent Discharge from Springs in Two Bavarian Nature Conservation Areas. Diversity 2023, 15, 915. https://doi.org/10.3390/d15080915
Cantonati M, Casoria C, Gerecke R, Bilous OP, Maisto G, Segadelli S, Spitale D, Steinbauer A, Vogel S, Saber AA. Diatom Indicators of Fluctuating/Intermittent Discharge from Springs in Two Bavarian Nature Conservation Areas. Diversity. 2023; 15(8):915. https://doi.org/10.3390/d15080915
Chicago/Turabian StyleCantonati, Marco, Chiara Casoria, Reinhard Gerecke, Olena P. Bilous, Giulia Maisto, Stefano Segadelli, Daniel Spitale, Alana Steinbauer, Sebastian Vogel, and Abdullah A. Saber. 2023. "Diatom Indicators of Fluctuating/Intermittent Discharge from Springs in Two Bavarian Nature Conservation Areas" Diversity 15, no. 8: 915. https://doi.org/10.3390/d15080915