1. Introduction
The global expansion of the wind energy industry continues apace [
1,
2,
3]. There are potentially beneficial effects of wind energy capture for biodiversity as a largely sustainable and renewable source of energy supply mitigating human-induced climate change [
4,
5], but there are environmental concerns [
1,
6].
Birds’ interactions with wind turbines are a concern, and research has often focused on large raptors. This focus is largely on large raptors’ demonstrable or purported vulnerability to collision with turbine blades [
7,
8,
9,
10,
11,
12,
13,
14,
15,
16,
17,
18,
19].
Large raptors also possess life history traits sensitive to additive mortality from anthropogenic sources [
20,
21,
22], notably in older individuals [
7,
23,
24,
25,
26,
27,
28]. Alarms have been raised on collision mortality potentially affecting population viability in several species [
28,
29,
30,
31,
32,
33].
Another potentially adverse effect is avoidance [
34], including avoidance of turbine arrays (wind farms) or individual turbines (macro- or meso-/micro-avoidance, respectively: [
34]), through disturbance and displacement [
35,
36,
37,
38] or wariness [
39,
40]. This can produce functional habitat loss [
35,
36,
37,
38,
39,
40].
Despite an erstwhile predominance of recorded or assumed collision mortality, several studies have nonetheless concluded avoidance as the main response of large raptors to wind turbines, highlighting the consequential functional (indirect) habitat loss [
37,
39,
40,
41,
42,
43,
44]. Functional habitat loss can incur serious impacts on large raptor populations. It may potentially affect territory occupancy and reproductive output [
45,
46] in part because individuals occur at low densities with extensive habitat requirements [
47]. The significance of any habitat loss to territorial birds will depend not just on the amount of lost habitat but also on whether there is scope for compensatory territorial boundary reconfiguration which will, in turn, depend on the adjacent landscape suitability and the proximity of neighbouring pairs [
46]. The use of key resources by migratory individuals may also be affected detrimentally [
37,
42].
Simplistically, avoidance is antagonistic to collision. A bird that does not enter a wind farm or avoids flying close to turbine blades cannot collide with a turbine blade and be killed. The two processes are not necessarily mutually exclusive in risk, however [
34,
36,
37,
39,
40,
41]. The observed balance in risk between avoidance (towards functional habitat loss) and collision (towards fatality) appears complex, involving several factors [
13,
15,
34,
39,
40]. Understanding these risk factors to predict the most likely adverse impact is critical in environmental impact assessments of future wind farm proposals [
38,
39,
40,
48].
The golden eagle
Aquila chrysaetos is well-studied in wind farm research. In North America, the consensus is that both non-territorial and territorial birds are mostly vulnerable to collision and consequent fatalities [
7,
14,
15,
18,
19,
28,
49,
50]. Avoidance may be more likely in migrating eagles [
42] through the use of different sources of wind energy uplift to some resident birds [
51,
52]. First-year (juvenile) eagles at Altamont in California, USA were less likely collision victims [
28].
In Europe, the spatial coincidence between wind farm development and golden eagle distribution has been described repeatedly [
53,
54,
55]. To our knowledge, however, there are no reports of golden eagle collision fatality rates from European wind farms comparable to those from North America. Scottish studies empirically concluded avoidance as the substantive response of golden eagles to wind turbines [
39,
40,
56,
57,
58].
The risk of collision via proximity to turbines for non-territorial birds during natal dispersal in Scotland was increased if turbines were placed in a more preferred habitat, and at higher wind speeds, but not according to the age of turbines and their specifications or birds’ age [
39,
40]. Given the number of turbines, the duration of their operation, and their coincidence with the distribution of numerous eagles [
40,
53], however, reported collision fatalities were very low [
39]. In confirmation, a subsequent update (NatureScot unpublished data) lists only five collisions over a 10-year period.
This observation shows the balance between avoidance and collision is not either/or [
36,
37,
39,
41]. It is independently consistent, nevertheless, with the predominant reactions of GPS-tagged dispersing Scottish golden eagles to the presence of turbines, to avoid flying near them [
39,
40,
56,
57,
58]. This evidence from movement behaviour included records of spatial use before and after turbine presence [
40].
Most Scottish studies have involved non-territorial eagles. Territorial eagles may not show the same degree of avoidance as non-territorial dispersing birds because they are more spatially constrained to a territory. Non-territorial birds have a greater capacity to compensate for functional habitat loss by being able to utilise more areas free of turbines. An inherent cost of wariness in avoiding turbines, which occupy intrinsically preferred habitats [
40], should be greater for territorial birds. Moreover, despite no evidence suggesting habituation in non-territorial eagles [
40], repeated exposure to the same turbines may favour habituation in territorial birds (see also [
59]). This, again, could lessen avoidance responses so that collision may be more likely. Territorial birds also engage in flight displays, pair interactions, and territory defence behaviours that may distract from their ability to detect and respond to spinning turbine blades. We could expect, therefore, that territorial birds differ in their reaction to turbines and are less wary.
The primary study objective was to examine the responses of territorial golden eagles to operational wind farms in Scotland utilising GPS-telemetry data from tagged individuals, to add to previous studies of non-territorial birds [
39,
40,
57,
58]. Underlying this objective, we also sought to contrast the responses of territorial occupants against those of non-territorial intruders to the same turbines, in the expectation (see above) that non-territorial birds would be more wary of turbines. This accounted for any potential effects involving different wind farms. Additionally, we considered the potential influence of the habitat suitability of turbine locations and the surrounding habitat when this had been shown previously to influence eagles’ proximity to operational wind turbines [
39].
3. Results
3.1. Descriptive Statistics
Both bird classes (territorial and non-territorial intruders) were rarely recorded close to turbines at any of the 11 wind farms (
Table 3). Although there were differences between wind farms (
Figure 2), even the closest mean approach distance (368 m for dispersing birds at Blary Hill) was a considerable distance from a potential turbine tip which has a radius of 50.5 m from the hub. If 2.5% quantiles are considered, the closest distance was 100 m by a territorial bird at Beinn Ghlas, where the turbine radius was only 22 m (
Table 3). The mean approach distance was greater for territorial birds (656 m v 572 m), but there was considerable overlap with non-territorial birds (95% empirical confidence limits 152 m–998 m for territorial birds and 175 m–979 m for non-territorial birds).
Twenty-point four percent (22,472/110,267) of random points were “inside” wind farm boundaries in 2D space. Only 3.4% (mean % per bird, sd 3.91%, range 0.4–12.9%) of territorial bird records were “inside” wind farms from 2D data, and this reduced to 1.3% (mean % per bird, sd 1.68%, range 0–4.7%) after adjustment for 3D measures. After adjusting the random points, using the proportion of territorial bird records inside 3D space (790/1725 = 0.458), the number of random points inside wind farms reduced to 10,292 or 9.3% of the total. Therefore, the proportion of territorial bird records inside the wind farms’ 3D volumes was considerably smaller than the proportion of random points.
For non-territorial birds, inside wind farms on 2D measures, the comparable percentage of records was slightly larger than for territorial birds, 5.9% (mean % per bird, sd 5.99%, range 0.4–19.5%), or 4.0% (mean % per bird, sd 5.85%, range 0–18.7%) when adjusted for 3D measures. After adjusting the random points, using the proportion of dispersing bird records inside 3D space (20,198/21,065 = 0.959), the number of random points inside wind farms’ 3D volumes reduced to 21,55 or 19.5% of the total. The proportion of non-territorial bird records inside wind farms’ 3D volumes was again considerably smaller than that of random points, but the difference was less than that for territorial birds.
Descriptively, these results suggest that bird records of both classes inside wind farms were lower than expected if they were randomly distributed. This was against additional expectations that territorial birds could show less evidence of being wary of turbine locations. The range of percentages of records inside the wind farm highlights the importance of differences between individuals which is also highlighted by the statistical model (
Section 3.2). The tagged eagle with the second highest proportion of records inside a wind farm, as a non-territorial bird, was the same individual with the highest proportion of records inside a wind farm for a territorial bird after later settling on a territory (tag 1025 consequently appears in both data sets,
Table 2).
Restricting the analyses to only those locations over topographically preferred habitats did not change the interpretation. In 2D space, 15.5% (5089/32,768) of the random points were inside wind farm boundaries while only 3.9% (mean % per bird, sd 4.54%, range 0.0–12.2%) of territorial bird records were “inside” wind farms, and this reduced to 1.8% (mean % per bird, sd 3.04%, range 0–9.1%) after adjustment for 3D measures. After adjusting the random points, using the proportion of territorial bird records inside 3D space (790/1725 = 0.458), the number of random points inside wind farms reduced to 10,292 or 9.3% of the total. Therefore, the proportion of territorial bird records inside the wind farms’ 3D volumes was considerably smaller than the proportion of random points. This magnitude of difference suggested that territorial birds’ responses to turbines were more likely due to the presence of turbines, rather than less preferred habitat within wind farms.
In the analyses restricted only to preferred habitat locations, the percentage of records from non-territorial birds inside wind farms in 2D space was similar to that for territorial birds, 4.2% (mean % per bird, sd 4.79%, range 0.3–14.1%), or 3.1% (mean % per bird, sd 4.59%, range 0.0–13.7%) when adjusted for 3D measures. After adjusting the random points, using the proportion of dispersing bird records inside 3D space (10,975/11,483 = 0.96), the number of random points inside wind farms’ 3D volumes reduced to 4885 or 15.0% of the total. Therefore, as with the full data set, the proportion of non-territorial bird records inside wind farms’ 3D volumes was smaller than that of random points and similar to that for territorial birds.
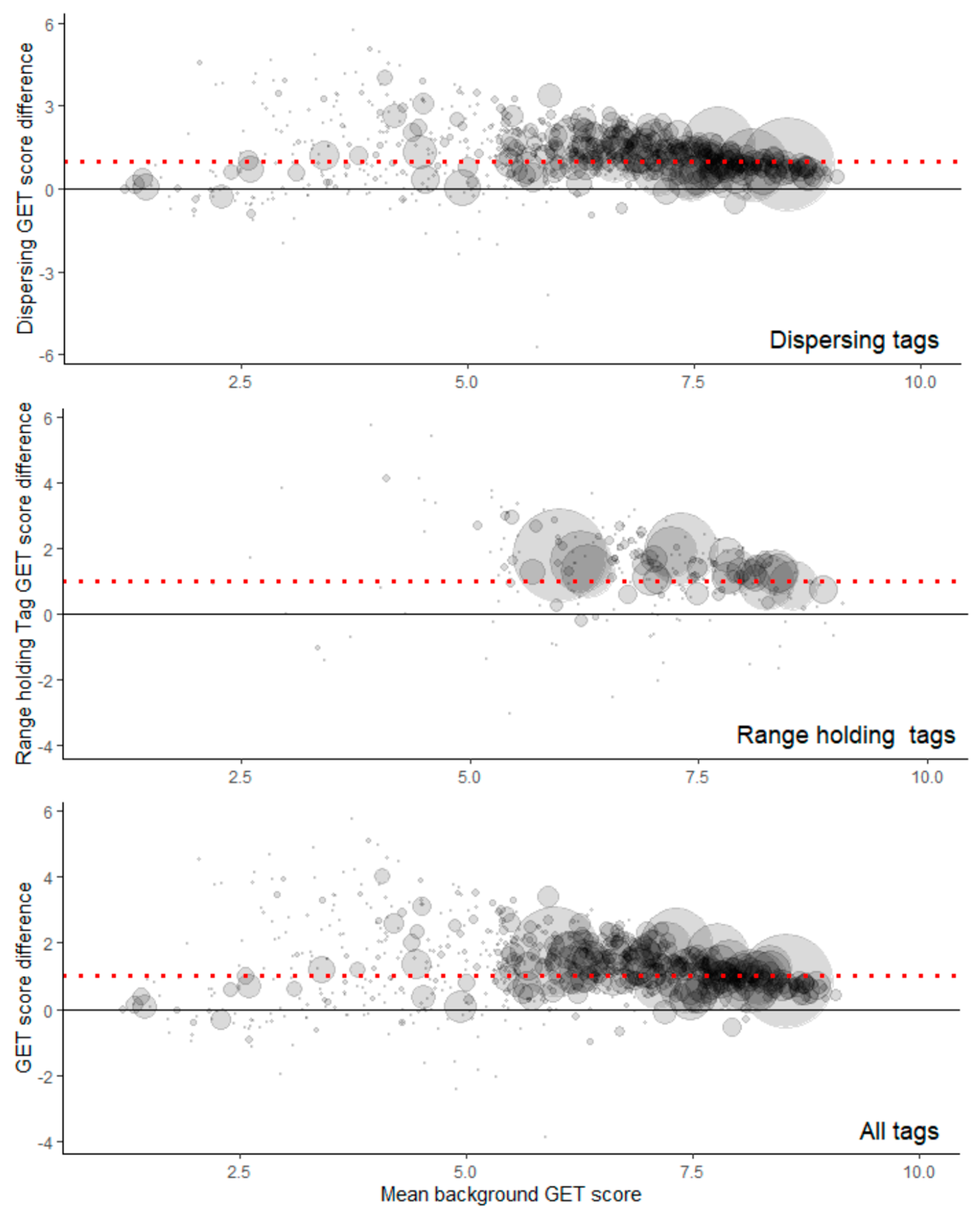
GET values also did not suggest subjectively that this substantial difference in random vs. empirical records inside wind farms was due to locational disparities in underlying habitat preference (
Table 4). Data on GET values again suggest little difference between territorial and non-territorial birds’ records, either inside or outside wind farms. Outside of wind farms, there were indications that both classes of birds were prone to select areas of higher habitat preference compared with random locations (
Table 4).
3.2. Analytical Statistical Results
Summaries of the four model structures are given in
Table 5, and
Table 6 shows the model comparison results. The model comparison results suggest that birds’ ‘class’ (territorial or non-territorial) did not improve the model fit, compared with the null model (m0 v m1,
p = 0.255:
Table 6). This confirmed that approach distances to the same turbines, by territorial range holders and intruding non-territorial birds, did not have a statistically significant difference.
However, including the GET difference predictor resulted in a significant improvement in the model fit (m0 v m2,
p < 0.001) (
Table 6). Although the model with both fixed effects improved the fit compared with the null model (m0 v m3,
p < 0.001), adding class (birds’ territorial status) did not improve the model compared with the single predictor GET difference model (m2 v m3,
p = 0.112). Conversely, adding GET difference to a model containing class (birds’ territorial status) improved the model compared with the class-only model (m1 v m3,
p < 0.001).
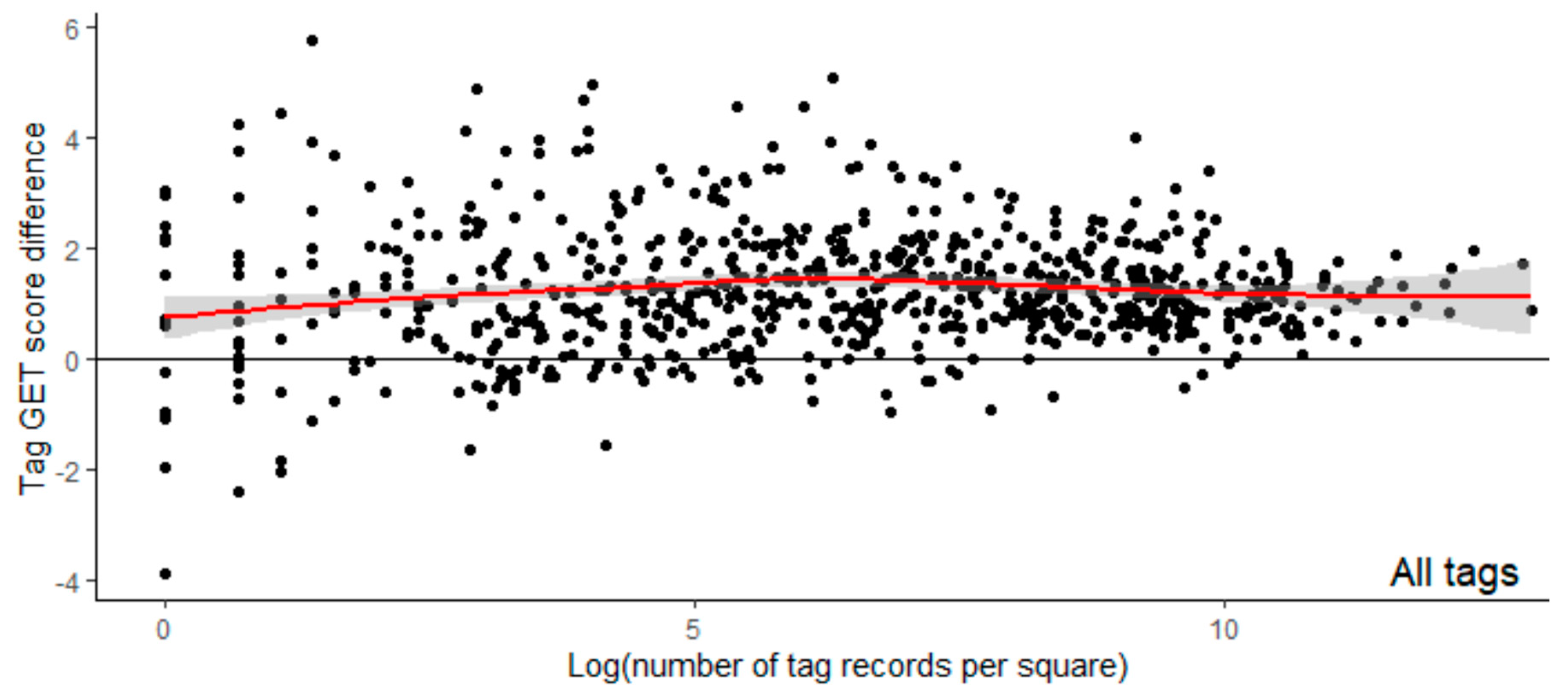
The best model (via AIC and model comparisons) was, therefore, the model containing the difference in GET scores as a single fixed effect (
Table 6). The difference in GET scores (averaged over 100 m) was significantly associated with the distance from an eagle to the nearest turbine (χ
2 = 7251.6,
p < 0.001), increasing the distance by ~24 m ± 15.1 m (standard errors) for every unit increase in the GET value difference (
Table 6).
Although the model including the GET difference had a significantly improved fit compared with the null model, the extra deviance explained by the GET difference was small compared with that explained by the random factors (
Table 6). The marginal coefficient of determination (deviance attributable to the fixed factors) was 1.9% (marginal R
2) compared with 43.0% (conditional R
2) for the conditional coefficient of determination (
Table 5). Therefore, although significant, the GET difference fixed effect added little additional explanation to the change in distance to a turbine when compared with that explained by the two random factors: the identity of the eagle (independent of territorial occupancy class) and the wind farm.
4. Discussion
We found that territorial eagles did not differ significantly from non-territorial eagles in their approach distances to the same wind turbines at 11 wind farms. Their response, consistent with previous research in Scotland (only on dispersing non-territorial eagles: [
39,
40]) was suubstantially avoidance that was conditional on the relative attractiveness of habitat adjacent to and within the wind farm.
In the present study, through focusing on territorial eagles, we did not have the benefit of comparing eagles’ use of a wind farm location before and after the turbines’ operation, as was available previously for non-territorial eagles [
39,
40]. Such before and after data give greater assurance that any paucity of eagle records within a wind farm or close to its turbines is due to the turbines and not any intrinsic difference in habitat suitability where turbines were placed. A conclusion towards avoidance, rather than wind farms being in less suitable habitats, however, was supported by only including pruned random and eagle use locations in preferred habitat locations. Additional support to this conclusion was also provided by the habitat preference scores inside and outside wind farms (
Table 4).
There was an expectation that avoidance could be less for territorial birds. This expectation included greater constraints on habitat availability (including spatial use) imposed by territoriality so that the preferred habitat around wind turbines would be proportionately more valuable. Moreover, repeated exposure to the same turbines, through spatial constraints, combined with (typically) the greater age of territorial birds could increase the prospect of habituation in territorial birds. Behavioural differences imposed by territoriality could also lead to closer approaches to turbine blades.
Despite this expectation, we found no evidence for closer approaches towards turbines by territorial eagles. Rather, while not significant, the analytical results erred towards non-territorial birds being more prone to closer approaches to the same turbines. Predominantly, therefore, the main impact of wind turbines on territorial eagles in Scotland—as in non-territorial conspecifics—was functional habitat loss through avoidance of displacing birds from otherwise suitable habitats.
However, in keeping with previous studies based only on non-territorial Scottish eagles [
39,
40], we found that the degree of territorial birds’ avoidance was conditional on the attractiveness of both the immediate habitat surrounding turbines and turbines’ locations in a wind farm. Despite the primary overarching tendency towards avoidance and functional habitat loss, the risk of collision (proxied by approach distances to turbine blades) increased when turbines were in locations of and surrounded by preferred habitats.
Thus, the two primary potentially adverse effects (collision vs. avoidance) are not “either/or” but represent antagonistic extremities on a continuum [
34,
37,
39,
40,
41]. Different species, populations, and individuals may occupy different places on this continuum, which may shift according to the stage of the life cycle, time, or several other factors [
13,
34].
On such factors, the present study showed that individual birds’ behaviour and different wind farms were strongly influential on eagles’ proximity to wind turbines. The locations of individual turbines and wind farms have been documented previously as influential in studies of griffon vultures
Gyps fulvus by dint of collision fatalities coinciding with modelling of topographic and weather-related drivers [
101,
102]. This Spanish vulture research prefaces our findings from Scottish golden eagles; although, the prospect of collision appears higher in the Spanish vultures, likely because of different species’ flight modes [
39].
Previous Scottish studies found no suggestion that non-territorial golden eagles changed their response to wind turbines with temporal exposure, either via their age or the age of the turbines [
40]. The present study also confirmed that there was no evidence of territorial birds’ habituation in their responses to turbines (see also [
38,
44,
103]). While we did not examine the age of birds per se explicitly, it was implicit in the comparisons between territorial and dispersing birds, since the former were typically older (
Table 2) and incurred greater repeated exposure to the same turbines, several of which had been operational for many years (
Table 1).
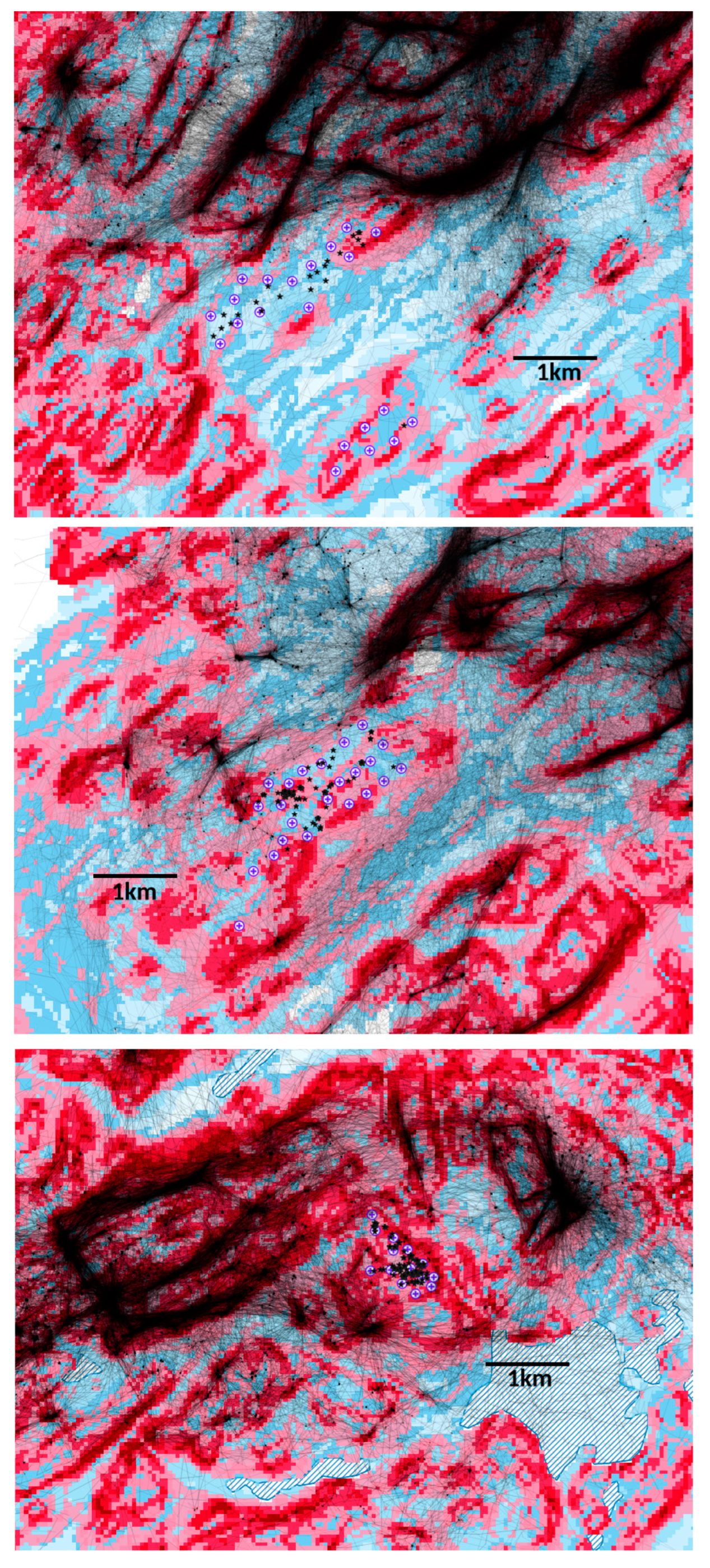
Our finding that territorial golden eagles in Scotland typically avoid wind turbines begs the question of whether this may affect their choice of nest sites or their occupation or re-occupation of formerly vacant territories which now include wind farms. Subjectively, there appears to be no discernible effect. For example, within the present study, nest sites were used or newly established which were 1.9 km and 1.5 km away when there were no closer alternatives nearer wind farms (A’Chruach and Cruach Mhor wind farms, respectively:
Table 1,
Figure 2). Re-occupation of vacant territories by tagged birds has occurred despite the proximity of wind farms (e.g., Beinn Ghlas:
Table 1,
Figure 2).
Anecdotally, despite a presumptive planning guideline for Scottish wind farms to be discouraged within the ‘core’ of a territory (i.e., within 3 km of nest sites: [
73]), to our knowledge, nine pairs with at least one tagged partner have established nest sites or new territories involving nests located within 2 km of an operational wind farm. In the larger non-tagged population, there are further examples, though several will not have been documented, with the closest nest 125 m from the nearest turbine (R. Reid personal communication). Intriguingly, it appears that while Scottish territorial golden eagles are wary of most turbines and so avoid them, this does not necessarily transfer to their choice of nest sites as regards ‘disturbance distance’ or apparently ‘accepting’ them as (typically) largely unsuitable locations within their home range.
The present study re-affirms and expands on an apparent difference between the response to wind turbines by golden eagles in Scotland [
39,
40,
56,
57,
58] and in the USA [
7,
14,
15,
18,
19,
28,
49]. Previous speculation on this difference suggested that eagles could be more wary of turbines in Scotland because of greater historical and contemporary persecution by humans, rendering it beneficial to survival if any novel anthropogenic feature of the environment (such as wind turbines) is avoided [
39,
40]. There may also be planning differences if Scottish turbines are less often consented to in highly preferred eagle habitats, but this seems highly unlikely—at least in recent years (post-initial wind farm installations at Altamont in California) with increasing knowledge—given the enhanced protected status of the golden eagle in the USA (e.g., [
14]).
Elsewhere in mainland Europe, there have been several studies highlighting the overlap between golden eagle distribution and potential wind farm development e.g., [
54,
55,
104]. These studies typically emphasised the potential adverse impact of collision mortality, following from earlier USA findings. To our knowledge, however, we are unaware of any subsequent European study which has subsequently confirmed the collision mortality rates documented in the USA. The apparent absence of any empirical study of golden eagles at post-operational wind farms in mainland Europe, or any reporting of fatality rates (other than incidental records) in the peer-reviewed literature, is curious and may have several causes. It deserves further attention and outputs, especially when (for example), in some regions of Spain, monitoring of collision fatalities is mandatory [
48].
In Scotland, golden eagle and wind farm development overlap in distribution was similarly prefaced [
53] and has subsequently been reported on ([
39,
40], present study). In Scotland, the response to the coincidence of wind farm development and golden eagle distribution is primarily avoidance but does not exclude the possibility of collision in some circumstances. Records of collision fatalities are correspondingly rare (Introduction) despite a marked overlap between eagle distribution and substantial movements with wind farm distribution. However, it is important to caveat conclusions from studies such as [
53] because the measure of what constituted suitable eagle habitat was a broad measure, but satellite tracking data show that quite fine-scale characteristics of the habitat are important in determining the movements of eagles; for example, see the flight lines in
Figure 2.
In conclusion, while our results show that territorial eagles were like non-territorial eagles in substantively avoiding wind turbines, there were clearly common factors that may lead to greater prospects of collision, supporting those outlined previously [
39,
40]. The primary factor was if turbines were in highly preferred habitats surrounded by similarly highly preferred habitats.
In wind farm design planning, attempts to avoid such habitat would not only be beneficial by way of reducing prospective collision risk—even if fundamentally unlikely in Scotland—but would also reduce the functional habitat loss through avoidance. Such considerations based on habitat are not new at wider landscape scales (e.g., [
31,
32,
33]) or at local installation scales [
105], or specific turbine scales [
101,
102].
Our study emphasises the importance of wind farm design to include a focus on specific turbine locations and to identify prospective potential effects appropriately. In this respect, we recommend the involvement of the GET model for golden eagles and other facultative soaring large raptors (when it can be readily transferred: [
72])—given a persistent finding of its predictions on habitat preference influence in informing proximity to turbine locations. For several large raptors, similar models to GET could provide an empirically based tool by which potential wind farm sites can be assessed and by which designs on specific turbine locations can be adjusted to avoid the potentially most problematic areas.
Whether via GET [
72], other vulnerability modelling [
19,
101,
102], or robust empirical assessment utilisation data, proposed specific turbine locations should be placed under special scrutiny in the planning process for large raptors, as they can differ in potential effect and impact (see also [
50]). It is particularly important that proposed turbine locations with high habitat preference (and with similar surroundings), and the outer turbine locations, are evaluated as avoidance of these turbines on, for example, a ridge which could result in additional habitat losses outside of the wind farm.
A key and generic international takeaway from the present and earlier studies of Scottish golden eagles’ responses to turbines is that, if proposed turbines are in a highly preferred habitat, then regardless of the base response to turbines (on the avoidance—collision continuum), such locations should be problematic in wind farm planning. This is because, if consented, they will either disproportionately increase the risk of greater functional habitat loss or increase the risk of collision fatalities. The two outcomes differ in their effects, but each has detrimental impacts.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}