
Effects of an Invasive Mud Crab on a Macroalgae-Dominated Habitat of the Baltic Sea under Different Temperature Regimes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Acclimation of Mud Crabs
2.2. Collection of Fucus vesiculosus and Associated Invertebrates
2.3. Experimental Design and Setup
2.4. Data Collection and Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson JB, C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef]
- Sumaila, U.R.; Tai, T.C. End overfishing and increase the resilience of the ocean to climate change. Front. Mar. Sci. 2020, 7, 523. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Bailey, S.A.; Brown, L.; Campbell, M.L.; Canning-Clode, J.; Carlton, J.T.; Castro, N.; Chainho, P.; Chan, F.T.; Creed, J.C.; Curd, A.; et al. Trends in the detection of aquatic non-indigenous species across global marine, estuarine and freshwater ecosystems: A 50-year perspective. Divers. Distrib. 2020, 26, 1780–1797. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; van Kleunen, M.; Winter, M.; et al. Global rise in emerging alien species results from increased accessibility of new source pools. Proc. Natl. Acad. Sci. USA 2018, 115, E2264–E2273. [Google Scholar] [CrossRef]
- Boudreau, S.A.; Worm, B. Ecological role of large benthic decapods in marine ecosystems: A review. Mar. Ecol. Prog. Ser. 2012, 469, 195–213. [Google Scholar] [CrossRef]
- Reaka-Kudla, M.L. Crustaceans. In Encyclopedia of Biodiversity; Levin, S.A., Ed.; Academic Press: New York, NY, USA, 2001; Volume 1, pp. 915–943. [Google Scholar]
- Silliman, B.R.; Bertness, M.D. A trophic cascade regulates salt marsh primary production. Proc. Natl. Acad. Sci. USA 2002, 99, 10500–10505. [Google Scholar] [CrossRef]
- Gül, M.R.; Griffen, B.D. Burrowing behavior and burrowing energetics of a bioindicator under human disturbance. Ecol. Evol. 2019, 9, 14205–14216. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Wang, A.; Li, S.; Cui, B.; Bai, J.; Shao, D. Crab contributions as an ecosystem engineer to sediment turnover in the Yellow River Delta. Front. Mar. Sci. 2022, 9, 1019176. [Google Scholar] [CrossRef]
- Botto, F.; Valiela, I.; Iribarne, O.; Martinetto, P.; Alberti, J. Impact of burrowing crabs on C and N sources, control, and transformations in sediments and food webs of SW Atlantic estuaries. Mar. Ecol. Prog. Ser. 2005, 293, 155–164. [Google Scholar] [CrossRef]
- Wang, J.Q.; Zhang, X.D.; Jiang, L.F.; Bertness, M.D.; Fang, C.M.; Chen, J.K.; Hara, T.; Li, B. Bioturbation of burrowing crabs promotes sediment turnover and carbon and nitrogen movements in an estuarine salt marsh. Ecosystems 2010, 13, 586–599. [Google Scholar] [CrossRef]
- Fanjul, E.; Escapa, M.; Montemayor, D.; Addino, M.; Alvarez, M.F.; Grela, M.A.; Iribarne, O. Effect of crab bioturbation on organic matter processing in South West Atlantic intertidal sediments. J. Sea Res. 2015, 95, 206–216. [Google Scholar] [CrossRef]
- Aschenbroich, A.; Michaud, E.; Stieglitz, T.; Fromard, F.; Gardel, A.; Tavares, M.; Thouzeau, G. Brachyuran crab community structure and associated sediment reworking activities in pioneer and young mangroves of French Guiana, South America. Estuar. Coast. Shelf Sci. 2016, 182, 60–71. [Google Scholar] [CrossRef]
- van der Zee, E.M.; Angelini, C.; Govers, L.L.; Christianen MJ, A.; Altieri, A.H.; van der Reijden, K.J.; Silliman, B.R.; van de Koppel, J.; van der Geest, M.; van Gils, J.A.; et al. How habitat-modifying organisms structure the food web of two coastal ecosystems. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152326. [Google Scholar] [CrossRef]
- Falk-Petersen, J.; Renaud, P.; Anisimova, N. Establishment and ecosystem effects of the alien invasive red king crab (Paralithodes camtschaticus) in the Barents Sea—A review. ICES J. Mar. Sci. 2011, 68, 479–488. [Google Scholar] [CrossRef]
- Kotta, J.; Wernberg, T.; Jänes, H.; Kotta, I.; Nurkse, K.; Pärnoja, M.; Orav-Kotta, H. Novel crab predator causes marine ecosystem regime shift. Sci. Rep. 2018, 8, 4956. [Google Scholar] [CrossRef]
- Howard, B.R.; Francis, F.T.; Côté, I.M.; Therriault, T.W. Habitat alteration by invasive European green crab (Carcinus maenas) causes eelgrass loss in British Columbia, Canada. Biol. Invasions 2019, 21, 3607–3618. [Google Scholar] [CrossRef]
- Cornelius, A.; Wagner, K.; Buschbaum, C. Prey preferences, consumption rates and predation effects of Asian shore crabs (Hemigrapsus takanoi) in comparison to native shore crabs (Carcinus maenas) in northwestern Europe. Mar. Biodivers. 2021, 51, 75. [Google Scholar] [CrossRef]
- Clavero, M.; Franch, N.; Bernardo-Madrid, R.; López, V.; Abelló, P.; Queral, J.M.; Mancinelli, G. Severe, rapid and widespread impacts of an Atlantic blue crab invasion. Mar. Pollut. Bull. 2022, 176, 113479. [Google Scholar] [CrossRef] [PubMed]
- Hänfling, B.; Edwards, F.; Gherardi, F. Invasive alien Crustacea: Dispersal, establishment, impact and control. BioControl 2011, 56, 573–595. [Google Scholar] [CrossRef]
- Rato, L.D.; Crespo, D.; Lemos, M.F. Mechanisms of bioinvasions by coastal crabs using integrative approaches—A conceptual review. Ecol. Indic. 2021, 125, 107578. [Google Scholar] [CrossRef]
- Kotta, J.; Ojaveer, H. Rapid establishment of the alien crab Rhithropanopeus harrisii (Gould) in the Gulf of Riga. Est. J. Ecol. 2012, 61, 293. [Google Scholar] [CrossRef]
- Makino, W.; Miura, O.; Kaiser, F.; Geffray, M.; Katsube, T.; Urabe, J. Evidence of multiple introductions and genetic admixture of the Asian brush-clawed shore crab Hemigrapsus takanoi (Decapoda: Brachyura: Varunidae) along the Northern European coast. Biol. Invasions 2018, 20, 825–842. [Google Scholar] [CrossRef]
- Spiridonov, V.A.; Zalota, A.K. Understanding and forecasting dispersal of non-indigenous marine decapods (Crustacea: Decapoda) in East European and North Asian waters. J. Mar. Biol. Assoc. U. K. 2017, 97, 591–611. [Google Scholar] [CrossRef]
- González-Ortegón, E.; Jenkins, S.; Galil, B.S.; Drake, P.; Cuesta, J.A. Accelerated invasion of decapod crustaceans in the southernmost point of the Atlantic coast of Europe: A non-natives’ hot spot? Biol. Invasions 2020, 22, 3487–3492. [Google Scholar] [CrossRef]
- Fowler, A.E.; Forsström, T.; von Numers, M.; Vesakoski, O. The North American mud crab Rhithropanopeus harrisii (Gould, 1841) in newly colonized Northern Baltic Sea: Distribution and ecology. Aquat. Invasions 2013, 8, 89–96. [Google Scholar] [CrossRef]
- Turoboyski, K. Biology and ecology of the crab Rhithropanopeus harrisii ssp. tridentatus. Mar. Biol. 1973, 23, 303–313. [Google Scholar] [CrossRef]
- Normant, M.; Gibowicz, M. Salinity induced changes in haemolymph osmolality and total metabolic rate of the mud crab Rhithropanopeus harrisii Gould, 1841 from Baltic coastal waters. J. Exp. Mar. Biol. Ecol. 2008, 355, 145–152. [Google Scholar] [CrossRef]
- Aarnio, K.; Törnroos, A.; Björklund, C.; Bonsdorff, E. Food web positioning of a recent coloniser: The North American Harris mud crab Rhithropanopeus harrisii (Gould, 1841) in the northern Baltic Sea. Aquat. Invasions 2015, 10, 399–413. [Google Scholar] [CrossRef]
- Gagnon, K.; Boström, C. Habitat expansion of the Harris mud crab Rhithropanopeus harrisii (Gould, 1841) in the northern Baltic Sea: Potential consequences for the eelgrass food web. BioInvasions Rec. 2016, 5, 101–106. [Google Scholar] [CrossRef]
- Rybczyk, A.; Czerniejewski, P. Body weight, morphometry, and diet of the mud crab, Rhithropanopeus harrisii tridentatus (Maitland, 1874) in the Odra estuary, Poland. Crustaceana 2008, 81, 1289–1299. [Google Scholar]
- Hegele-Drywa, J.; Normant, M. Feeding ecology of the American crab Rhithropanopeus harrisii (Crustacea, Decapoda) in the coastal waters of the Baltic Sea. Oceanologia 2009, 51, 361–375. [Google Scholar] [CrossRef]
- Forsström, T.; Fowler, A.E.; Manninen, I.; Vesakoski, O. An introduced species meets the local fauna: Predatory behavior of the crab Rhithropanopeus harrisii in the Northern Baltic Sea. Biol. Invasions 2015, 17, 2729–2741. [Google Scholar] [CrossRef]
- Nurkse, K.; Kotta, J.; Orav-Kotta, H.; Pärnoja, M.; Kuprijanov, I. Laboratory analysis of the habitat occupancy of the crab Rhithropanopeus harrisii (Gould) in an invaded ecosystem: The north-eastern Baltic Sea. Estuar. Coast. Shelf Sci. 2015, 154, 152–157. [Google Scholar] [CrossRef]
- Jormalainen, V.; Gagnon, K.; Sjöroos, J.; Rothäusler, E. The invasive mud crab enforces a major shift in a rocky littoral invertebrate community of the Baltic Sea. Biol. Invasions 2016, 18, 1409–1419. [Google Scholar] [CrossRef]
- Liversage, K.; Kotta, J.; Kuprijanov, I.; Rätsep, M.; Nõomaa, K. A trophic cascade facilitates native habitat providers within assemblages of multiple invasive marine species. Ecosphere 2021, 12, e03621. [Google Scholar] [CrossRef]
- Lokko, K.; Kotta, J.; Orav-Kotta, H.; Nurkse, K.; Pärnoja, M. Introduction of a functionally novel consumer to a low diversity system: Effects of the mud crab Rhithropanopeus harrisii on meiobenthos. Estuar. Coast. Shelf Sci. 2018, 201, 132–139. [Google Scholar] [CrossRef]
- Nurkse, K.; Kotta, J.; Rätsep, M.; Kotta, I.; Kreitsberg, R. Experimental evaluation of the effects of the novel predators, round goby and mud crab on benthic invertebrates in the Gulf of Riga, Baltic Sea. J. Mar. Biol. Assoc. U. K. 2018, 98, 25–31. [Google Scholar] [CrossRef]
- Wikström, S.A.; Kautsky, L. Structure and diversity of invertebrate communities in the presence and absence of canopy-forming Fucus vesiculosus in the Baltic Sea. Estuar. Coast. Shelf Sci. 2007, 72, 168–176. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 29 April 2023).
- Lenth, R.L. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.8.5. 2023. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 29 April 2023).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.6. 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 29 April 2023).
- Hegele-Drywa, J.; Normant, M. Effect of temperature on physiology and bioenergetics of adult Harris mud crab Rhithropanopeus harrisii (Gould, 1841) from the southern Baltic Sea. Oceanol. Hydrobiol. Stud. 2014, 43, 219–227. [Google Scholar] [CrossRef]
- Hollebone, A.L.; Hay, M.E. An invasive crab alters interaction webs in a marine community. Biol. Invasions 2008, 10, 347–358. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Geburzi, J.C.; Brandis, D.; Buschbaum, C. Recruitment patterns, low cannibalism and reduced interspecific predation contribute to high invasion success of two Pacific crabs in northwestern Europe. Estuar. Coast. Shelf Sci. 2018, 200, 460–472. [Google Scholar] [CrossRef]
- Tomczak, M.T.; Müller-Karulis, B.; Järv, L.; Kotta, J.; Martin, G.; Minde, A.; Põllumäe, A.; Razinkovas, A.; Strake, S.; Bucas, M.; et al. Analysis of trophic networks and carbon flows in south-eastern Baltic coastal ecosystems. Prog. Oceanogr. 2009, 81, 111–131. [Google Scholar] [CrossRef]
- Griffiths, J.R.; Kadin, M.; Nascimento, F.J.; Tamelander, T.; Törnroos, A.; Bonaglia, S.; Bonsdorff, E.; Brüchert, V.; Gårdmark, A.; Järnström, M.; et al. The importance of benthic–pelagic coupling for marine ecosystem functioning in a changing world. Glob. Change Biol. 2017, 23, 2179–2196. [Google Scholar] [CrossRef]
- Reynolds, P.L.; Sotka, E.E. Non-consumptive predator effects indirectly influence marine plant biomass and palatability. J. Ecol. 2011, 99, 1272–1281. [Google Scholar] [CrossRef]
- Wahl, M.; Buchholz, B.; Winde, V.; Golomb, D.; Guy-Haim, T.; Müller, J.; Rilov, G.; Scotti, M.; Böttcher, M.E. A mesocosm concept for the simulation of near-natural shallow underwater climates: The Kiel Outdoor Benthocosms (KOB). Limnol. Oceanogr. Methods 2015, 13, 651–663. [Google Scholar] [CrossRef]
- Pansch, C.; Hiebenthal, C. A new mesocosm system to study the effects of environmental variability on marine species and communities. Limnol. Oceanogr. Methods 2019, 17, 145–162. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
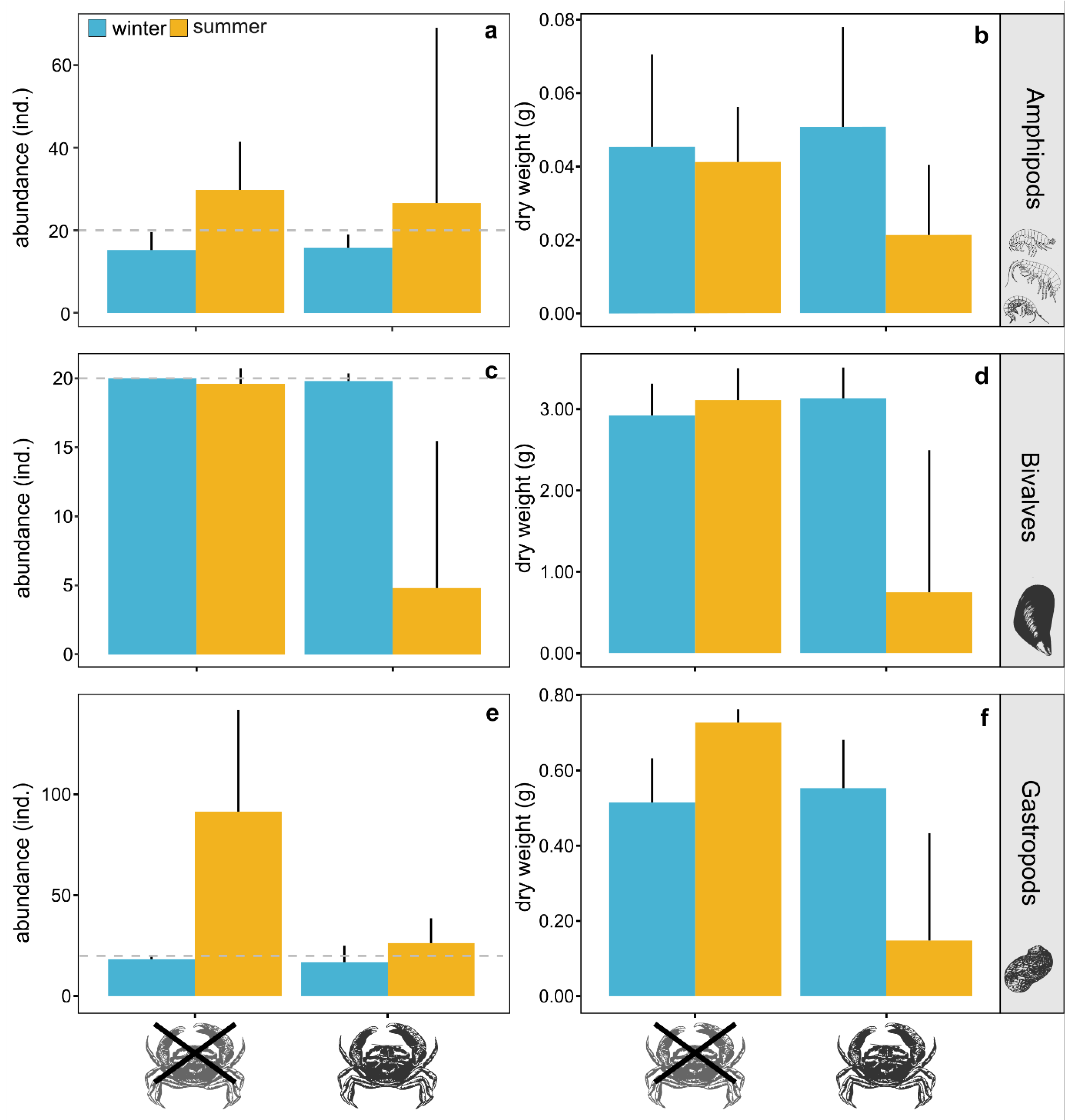
| Response Variable | Model Component | DF | SS | MSS | F-Value | p-Value |
|---|---|---|---|---|---|---|
| log(abundance of amphipods) | temperature | 1 | 0.01 | 0.01 | 0.008 | 0.9300 |
| Rh | 1 | 2.04 | 2.04 | 1.953 | 0.1810 | |
| temperature:Rh | 1 | 2.41 | 2.40 | 2.307 | 0.1480 | |
| residuals | 16 | 16.68 | 1.04 | |||
| log(abundance of bivalves) | temperature | 1 | 11.91 | 11.91 | 9.128 | 0.0081 |
| Rh | 1 | 11.75 | 11.75 | 9.001 | 0.0085 | |
| temperature:Rh | 1 | 11.59 | 11.59 | 8.882 | 0.0088 | |
| residuals | 16 | 20.88 | 1.31 | |||
| log(abundance of gastropods) | temperature | 1 | 5.00 | 5.00 | 22.64 | 0.0002 |
| Rh | 1 | 2.38 | 2.38 | 10.75 | 0.0047 | |
| temperature:Rh | 1 | 1.24 | 1.24 | 5.63 | 0.0305 | |
| residuals | 16 | 3.54 | 0.22 | |||
| log(dry weight of amphipods) | temperature | 1 | 1.36 | 1.36 | 4.063 | 0.0610 |
| Rh | 1 | 0.78 | 0.78 | 2.333 | 0.1462 | |
| temperature:Rh | 1 | 1.14 | 1.14 | 3.399 | 0.0838 | |
| residuals | 16 | 5.35 | 0.33 | |||
| log(dry weight of bivalve) | temperature | 1 | 6.45 | 6.45 | 11.57 | 0.0036 |
| Rh | 1 | 6.37 | 6.37 | 11.43 | 0.0038 | |
| temperature:Rh | 1 | 7.17 | 7.17 | 12.87 | 0.0025 | |
| residuals | 16 | 8.91 | 0.56 | |||
| dry weight of gastropods | temperature | 1 | 0.05 | 0.05 | 2.53 | 0.1315 |
| Rh | 1 | 0.37 | 0.37 | 19.93 | 0.0004 | |
| temperature:Rh | 1 | 0.48 | 0.48 | 25.97 | 0.0001 | |
| residuals | 16 | 0.29 | 0.02 | |||
| decrease in dry weight of Fucus vesiculosus | temperature | 1 | 1558.30 | 1558.30 | 23.04 | 0.0002 |
| Rh | 1 | 122.20 | 122.20 | 1.81 | 0.1977 | |
| temperature:Rh | 1 | 409.90 | 409.90 | 6.06 | 0.0256 | |
| residuals | 16 | 1082.30 | 67.60 |
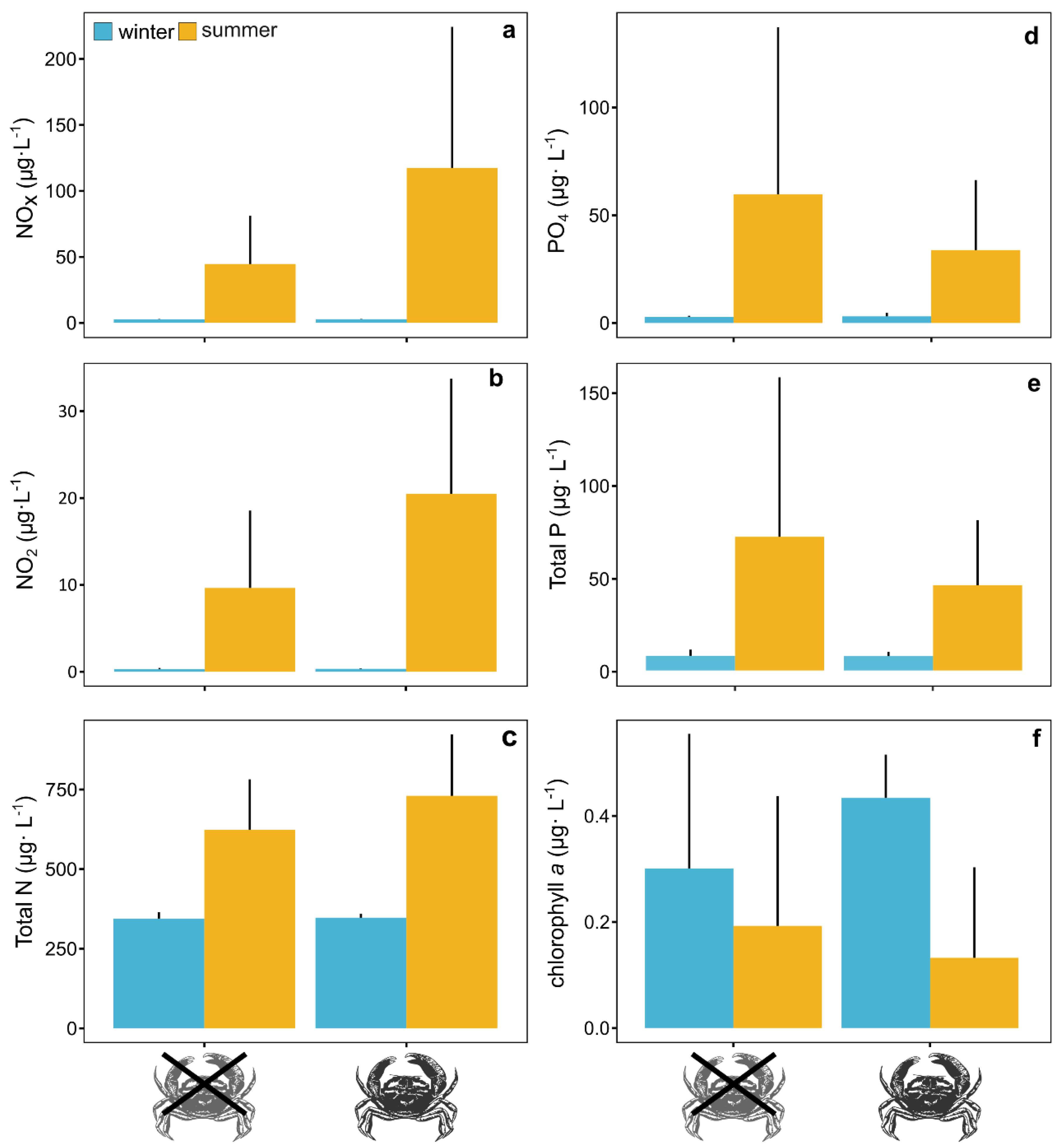
| Response Variable | Model Component | DF | SS | MSS | F-Value | p-Value |
|---|---|---|---|---|---|---|
| log(Total N) | temperature | 1 | 2.12 | 2.12 | 92.16 | <0.0001 |
| Rh | 1 | 0.03 | 0.03 | 1.44 | 0.2470 | |
| temperature:Rh | 1 | 0.03 | 0.03 | 1.12 | 0.3070 | |
| residuals | 16 | 0.37 | 0.02 | |||
| log(NOX) | temperature | 1 | 45.27 | 45.27 | 115.42 | <0.0001 |
| Rh | 1 | 1.28 | 1.28 | 3.27 | 0.0895 | |
| temperature:Rh | 1 | 1.02 | 1.02 | 2.60 | 0.1261 | |
| residuals | 16 | 6.27 | 0.39 | |||
| log(NO2) | temperature | 1 | 66.07 | 66.07 | 218.42 | <0.0001 |
| Rh | 1 | 1.39 | 1.39 | 4.61 | 0.0475 | |
| temperature:Rh | 1 | 0.74 | 0.74 | 2.46 | 0.1363 | |
| residuals | 16 | 4.84 | 0.30 | |||
| log(Total P) | temperature | 1 | 16.00 | 16.00 | 48.90 | <0.0001 |
| Rh | 1 | 0.11 | 0.11 | 0.34 | 0.5690 | |
| temperature:Rh | 1 | 0.11 | 0.11 | 0.33 | 0.5740 | |
| residuals | 16 | 5.24 | 0.33 | |||
| log(PO4) | temperature | 1 | 29.10 | 29.10 | 61.97 | <0.0001 |
| Rh | 1 | 0.23 | 0.23 | 0.49 | 0.4940 | |
| temperature:Rh | 1 | 0.27 | 0.27 | 0.57 | 0.4630 | |
| residuals | 16 | 7.51 | 0.47 | |||
| chlorophyll a | temperature | 1 | 0.21 | 0.21 | 8.08 | 0.0177 |
| Rh | 1 | 0.01 | 0.01 | 0.25 | 0.6227 | |
| temperature:Rh | 1 | 0.05 | 0.05 | 1.79 | 0.1996 | |
| residuals | 16 | 0.42 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beleem, I.B.; Kotta, J.; Barboza, F.R. Effects of an Invasive Mud Crab on a Macroalgae-Dominated Habitat of the Baltic Sea under Different Temperature Regimes. Diversity 2023, 15, 644. https://doi.org/10.3390/d15050644
Beleem IB, Kotta J, Barboza FR. Effects of an Invasive Mud Crab on a Macroalgae-Dominated Habitat of the Baltic Sea under Different Temperature Regimes. Diversity. 2023; 15(5):644. https://doi.org/10.3390/d15050644
Chicago/Turabian StyleBeleem, Imtiyaz B., Jonne Kotta, and Francisco R. Barboza. 2023. "Effects of an Invasive Mud Crab on a Macroalgae-Dominated Habitat of the Baltic Sea under Different Temperature Regimes" Diversity 15, no. 5: 644. https://doi.org/10.3390/d15050644