

Phylogeny, Distribution, and Biology of Pygmy Whitefish (Prosopium coulterii) in the Beringia Region (Chukotka)
 , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
3. Results
3.1. Distribution Patterns
3.2. Basic Biological Features
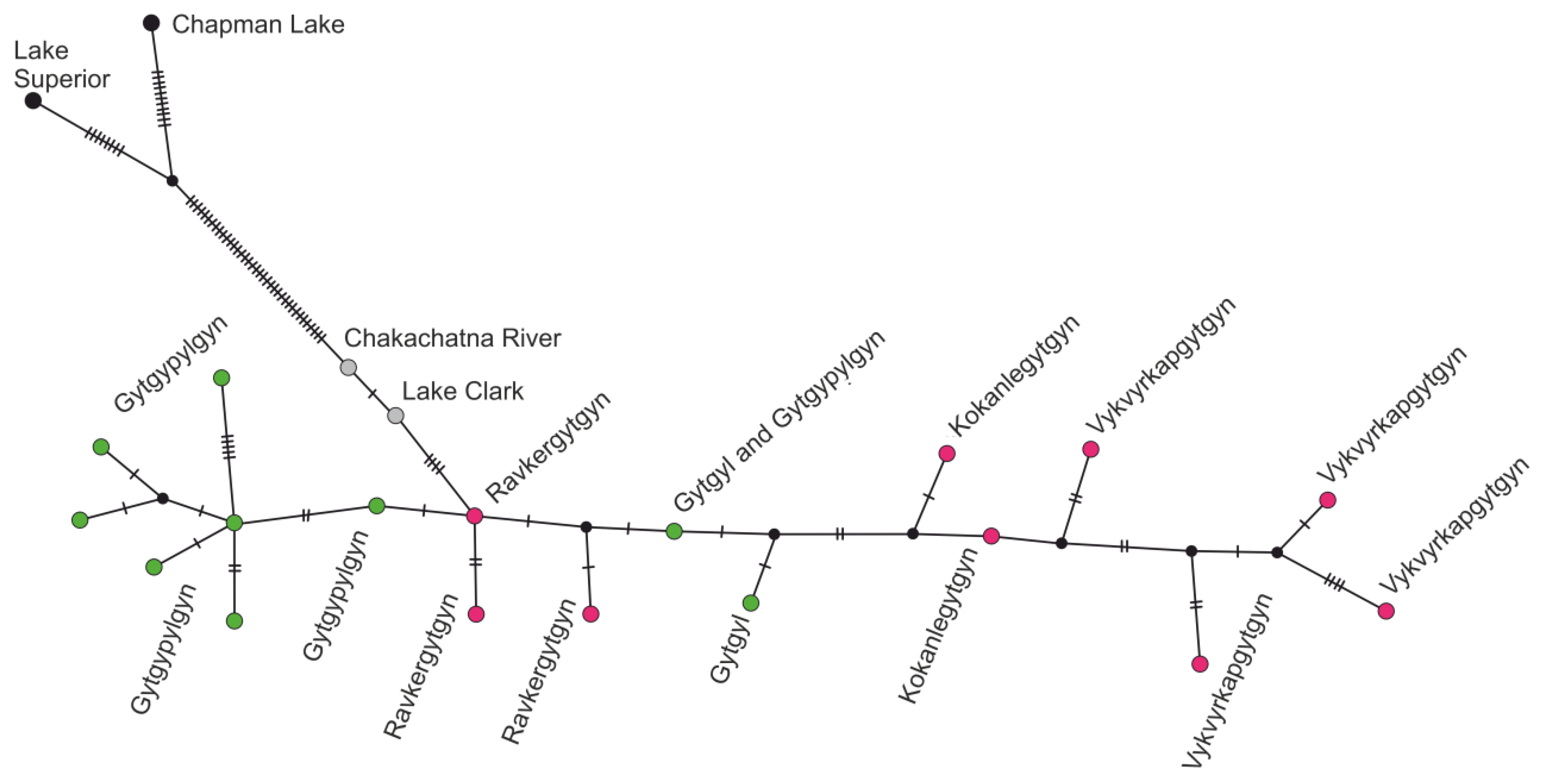
3.3. mtDNA Haplotype Diversity
4. Discussion
4.1. Distribution in Chukotka
4.2. Basic Biological Characteristics
4.3. Phylogeny within the Chukotkan Range
4.4. Global Phylogenetic History and Colonization Pathways
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Hewitt, G.M. Quaternary phylogeography: The roots of hybrid zones. Genetica 2011, 139, 617–638. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.R.; Lister, A.M. Cryptic northern refugia and the origins of the modern biota. Trends Ecol. Evol. 2001, 16, 608–613. [Google Scholar] [CrossRef]
- Shafer, A.B.; Cullingham, C.I.; Cote, S.D.; Coltman, D.W. Of glaciers and refugia: A decade of study sheds new light on the phylogeography of northwestern North America. Mol. Ecol. 2010, 19, 4589–4621. [Google Scholar] [CrossRef] [PubMed]
- Stamford, M.D.; Taylor, E.B. Phylogeographical lineages of Arctic grayling (Thymallus arcticus) in North America: Divergence, origins and affinities with Eurasian Thymallus. Mol. Ecol. 2004, 13, 1533–1549. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.N.; Taylor, E.B. Pleistocene glaciations and contemporary genetic diversity in a Beringian fish, the broad whitefish, Coregonus nasus (Pallas): Inferences from microsatellite DNA variation. J. Evol. Biol. 2010, 23, 72–86. [Google Scholar] [CrossRef]
- Bachevskaya, L.T.; Pereverzeva, V.V.; Ivanova, G.D.; Agapova, G.A.; Primak, A.A. Genetic structure of the Siberian Sucker (Catostomus catostomus rostratus) according to data on sequence variation of the mtDNA cytochrome b gene. Biol. Bull. 2014, 41, 306–311. [Google Scholar] [CrossRef]
- Bachevskaya, L.T.; Pereverzeva, V.V.; Primak, A.A.; Agapova, G.A. Genetic Variability of the Siberian Sucker Catostomus catostomus rostratus (Teleostei: Catastomidae) from Water Bodies of the Northeast of Russia. Russ. J. Genet. 2022, 58, 428–435. [Google Scholar] [CrossRef]
- Campbell, M.A.; Lopéz, J.A. Mitochondrial phylogeography of a Beringian relict: The endemic freshwater genus of blackfish Dallia (Esociformes). J. Fish Biol. 2014, 84, 523–538. [Google Scholar] [CrossRef]
- Campbell, M.A.; Takebayashi, N.; López, J.A. Beringian sub-refugia revealed in blackfish (Dallia): Implications for understanding the effects of Pleistocene glaciations on Beringian taxa and other Arctic aquatic fauna. BMC Evol. Biol. 2015, 15, 144. [Google Scholar] [CrossRef] [Green Version]
- McCart, P.J. Growth and Morphometry of the Pygmy Whitefish (Prosopium coulteri) in British Columbia. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 1963; pp. 1–97. [Google Scholar]
- Scott, W.B.; Crossman, E.J. Freshwater Fishes of Canada. Fish. Res. Board Can. Bull. 1973, 184, 966. [Google Scholar]
- Vecsei, P.; Panayi, D. Range extension for Pygmy Whitefish (Prosopium coulterii) in the Northwest Territories, Canada. Can. Field-Nat. 2015, 129, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Chereshnev, I.A.; Skopets, M.B. A new record of the pygmy whitefish, Prosopium coulteri, from the Amguem River Basin, (Chukotski Peninsula). J. Ichthyol. 1992, 32, 46–55. [Google Scholar]
- Eschmeyer, P.H.; Bailey, R.M. The pygmy whitefish, Coregonus coulteri, in Lake Superior. Trans. Am. Fish. Soc. 1955, 84, 161–199. [Google Scholar] [CrossRef]
- Lindsey, C.C.; Franzin, W.G. New complexities in zoogeography and taxonomy of the pygmy whitefish (Prosopium coulteri). J. Fish. Board Can. 1972, 29, 1772–1775. [Google Scholar] [CrossRef]
- Witt, J.D.; Zemlak, R.J.; Taylor, E.B. Phylogeography and the origins of range disjunctions in a north temperate fish, the pygmy whitefish (Prosopium coulterii), inferred from mitochondrial and nuclear DNA sequence analysis. J. Biogeogr. 2011, 38, 1557–1569. [Google Scholar] [CrossRef]
- Wiedmer, M.; Montgomery, D.R.; Gillespie, A.R.; Greenberg, H. Late Quaternary megafloods from Glacial Lake Atna, Southcentral Alaska, USA. Quat. Res. 2010, 73, 413–424. [Google Scholar] [CrossRef]
- Blanchfield, P.J.; Taylor, E.B.; Watkinson, D.A. Morphological and genetic analyses identify a new record of a glacial relict: Pygmy Whitefish (Prosopium coulterii) from Northwestern Ontario. Can. J. Zool. 2014, 92, 267–271. [Google Scholar] [CrossRef]
- Chereshnev, I.A.; Volobuev, V.V.; Shestakov, A.V.; Frolov, S.V. Salmonoid Fishes in Russian North-East. [Lososevidnye ryby Severo-Vostoka Rossii]; Dal’nauka: Vladivostok, Russia, 2002; p. 496. (In Russian) [Google Scholar]
- Chereshnev, I.A. Pygmy Whitefish. [Karlikovyj valek]. In Red Data Book of the Chukchi Autonomous District Vol. 1 Animals; Dikii Sever Publ: Magadan, Russia, 2008; p. 235. (In Russian) [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 1, 2–3. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Chereshnev, I.A. Biological Diversity of Freshwater Fish Fauna in the Russian North-East [Biologicheskoe Raznoobrazie Presnovodnoj Ihtiofauny Severo-Vostoka Rossii]; Dal’nauka: Vladivostok, Russia, 1996; p. 196. (In Russian) [Google Scholar]
- Knoll, L.B.; Williamson, C.E.; Pilla, R.M.; Leach, T.H.; Brentrup, J.A.; Fisher, T.J. Browning-related oxygen depletion in an oligotrophic lake. Inland Waters 2018, 8, 255–263. [Google Scholar] [CrossRef]
- Knudsen, R.; Amundsen, P.A.; Klemetsen, A. Arctic charr in sympatry with burbot: Ecological and evolutionary consequences. Hydrobiologia 2010, 650, 43–54. [Google Scholar] [CrossRef]
- Gowell, C.P.; Quinn, T.P.; Taylor, E.B. Coexistence and origin of trophic ecotypes of pygmy whitefish, Prosopium coulterii, in a south-western Alaskan lake. J. Evol. Biol. 2012, 25, 2432–2448. [Google Scholar] [CrossRef]
- Pavlidis, Y.A.; Ionin, A.S.; Medvedev, V.S. Paleogeography of the Late Wurm of the Beringian Shelf [Paleogeografiya pozdnego vyurma shel’fa Beringii]. In Geology and Geomorphology of Shelves and Continental Slopes; [Geologiya i Geomorfologiya Shel’fov i Materikovyh Sklonov]; Nauka: Moscow, Russia, 1985; pp. 65–76. (In Russian) [Google Scholar]
- Zardoya, R.; Doadrio, I. Molecular evidence on the evolutionary and biogeographical patterns of European cyprinids. J. Mol. Evol. 1999, 49, 227–237. [Google Scholar] [CrossRef]
- Oleinik, A.G. On the mutation rates of the mitochondrial and nuclear genomes of salmonid fishes. Russ. J. Mar. Biol. 2000, 26, 432–438. [Google Scholar] [CrossRef]
- Machordom, A.; Doadrio, I. Evidence of a Cenozoic Betic-Kabilian connection based on freshwater fish phylogeography (Luciobarbus, Cyprinidae). Mol. Phylogenet. Evol. 2001, 18, 252–263. [Google Scholar] [CrossRef]
- Glushkova, O.Y. Late Pleistocene glaciations in north-east Asia. Dev. Quat. Sci. 2011, 15, 865–875. [Google Scholar] [CrossRef]
- Oleinik, A.G.; Skurikhina, L.A.; Brykov, V.A. Phylogeny of charrs of the genus Salvelinus based on mitochondrial DNA data. Russ. J. Genet. 2015, 51, 55–68. [Google Scholar] [CrossRef]
- Lecaudey, L.A.; Schliewen, U.K.; Osinov, A.G.; Taylor, E.B.; Bernatchez, L.; Weiss, S.J. Inferring phylogenetic structure, hybridization and divergence times within Salmoninae (Teleostei: Salmonidae) using RAD-sequencing. Mol. Phylogenet. Evol. 2018, 124, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Duk-Rodkin, A.; Barendregt, R.W. Stratigraphical record of glacials/interglacials in northwest Canada. Dev. Quat. Sci. 2011, 15, 661–698. [Google Scholar] [CrossRef]
- COSEWIC. COSEWIC Assessment and Status Report on the Pygmy Whitefish Prosopium Coulterii, Southwestern Yukon Beringian Populations, Yukon River Populations, Pacific Populations, Western Arctic Populations, Great Lakes—Upper St. Lawrence populations, Water; Committee on the Status of Endangered Wildlife in Canada: Ottawa, ON, Canada, 2016. [Google Scholar]
- Esin, E.V.; Markevich, G.N. Evolution of the charrs, genus Salvelinus (Salmonidae). 1. origins and expansion of the species. J. Ichthyol. 2018, 58, 187–203. [Google Scholar] [CrossRef]
- Budy, P.; Rogers, K.B.; Kanno, Y.; Penaluna, B.E.; Hitt, N.; Thiede, G.P.; Dunham, J.; Mellison, C.; Somer, W.L.; DeRito, J. Distribution and status of trout and char in North America. In Trout and Char of the World; Kershner, J.L., Williams, J.E., Gresswell, R.E., Lobón-Cerviá, J., Eds.; American Fisheries Society: Vancouver, BC, Canada, 2019; Chapter 7; pp. 1–58. [Google Scholar] [CrossRef]
- Bernatchez, L.; Dodson, J.J. Phylogeographic structure in mitochondrial DNA of the lake whitefish (Coregonus clupeaformis) and its relation to Pleistocene glaciations. Evolution 1991, 45, 1016–1035. [Google Scholar]
- Geibert, W.; Matthiessen, J.; Stimac, I.; Wollenburg, J.; Stein, R. Glacial episodes of a freshwater Arctic Ocean covered by a thick ice shelf. Nature 2021, 590, 97–102. [Google Scholar] [CrossRef]
- Kaufman, D.S.; Young, N.E.; Briner, J.P.; Manley, W.F. Alaska palaeo-glacier atlas (version 2). Dev. Quat. Sci. 2011, 15, 427–445. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake (Basin, Region) Sample Size | COI Haplotypes | CytB Haplotypes | ATP6 Haplotypes |
|---|---|---|---|
| Kokanlegytgyn (Koluchinskaya, EC) 3 | 1COI/2; 3COI/1 | 1C/1; 3C/1 | 4A/1 |
| Vykvyrkapgytgyn (Koluchinskaya, EC) 8 | 1COI/4 | 1C/1; 5C/2; 6C/2; 7C/1; 8C/2 | 1A/3; 2A/5 |
| Ravkergytgyn (Amguema, EC) 4 | 1COI/3 | 1C/2; 2C/1; 4C/1 | 3A/4 |
| Gytgypylgyn (Pegtymel, WC) 8 | 1COI/3; 2COI/3; 4CO/1 | 1C/3; 3C/1; 9C/1; 10C/1; 11C/1; 12C/1 | 1A/1; 3A/2; 4A/2; 5A/1, 6A/1 |
| Gytgyl (Palavaam, WC) 2 | 2COI/2 | 9C/1; 13C/1 | 3A/1; 7A/1 |
| In total: 25 | 19 | 24 | 22 |
| Region | North America | Western Chukotka | Eastern Chukotka |
|---|---|---|---|
| Southern Alaska | 0.028; 0.036; 0.032 | 0.012; 0.004; 0.008 | 0.010; 0.004; 0.008 |
| North America | 0.026; 0.037; 0.031 | 0.022; 0.038; 0.031 | |
| Western Chukotka | 0.006; 0.002; 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markevich, G.N.; Solovyev, M.M.; Vlasenko, P.G.; Izotova, G.V.; Kashinskaya, E.N.; Bochkarev, N.A.; Politov, D.V.; Melnik, N.O.; Esin, E.V. Phylogeny, Distribution, and Biology of Pygmy Whitefish (Prosopium coulterii) in the Beringia Region (Chukotka). Diversity 2023, 15, 547. https://doi.org/10.3390/d15040547
Markevich GN, Solovyev MM, Vlasenko PG, Izotova GV, Kashinskaya EN, Bochkarev NA, Politov DV, Melnik NO, Esin EV. Phylogeny, Distribution, and Biology of Pygmy Whitefish (Prosopium coulterii) in the Beringia Region (Chukotka). Diversity. 2023; 15(4):547. https://doi.org/10.3390/d15040547
Chicago/Turabian StyleMarkevich, Grigorii N., Mikhail M. Solovyev, Pavel G. Vlasenko, Gelena V. Izotova, Elena N. Kashinskaya, Nikolai A. Bochkarev, Dmitry V. Politov, Nikolaii O. Melnik, and Evgeny V. Esin. 2023. "Phylogeny, Distribution, and Biology of Pygmy Whitefish (Prosopium coulterii) in the Beringia Region (Chukotka)" Diversity 15, no. 4: 547. https://doi.org/10.3390/d15040547