
Large-Scale Reduction in the Extent of Agriculture around Stopover Sites of Migratory Geese in European Russia between 1990 and 2015


Abstract
:1. Introduction
2. Materials and Methods
3. Results

{kind=link}
{kind=link}
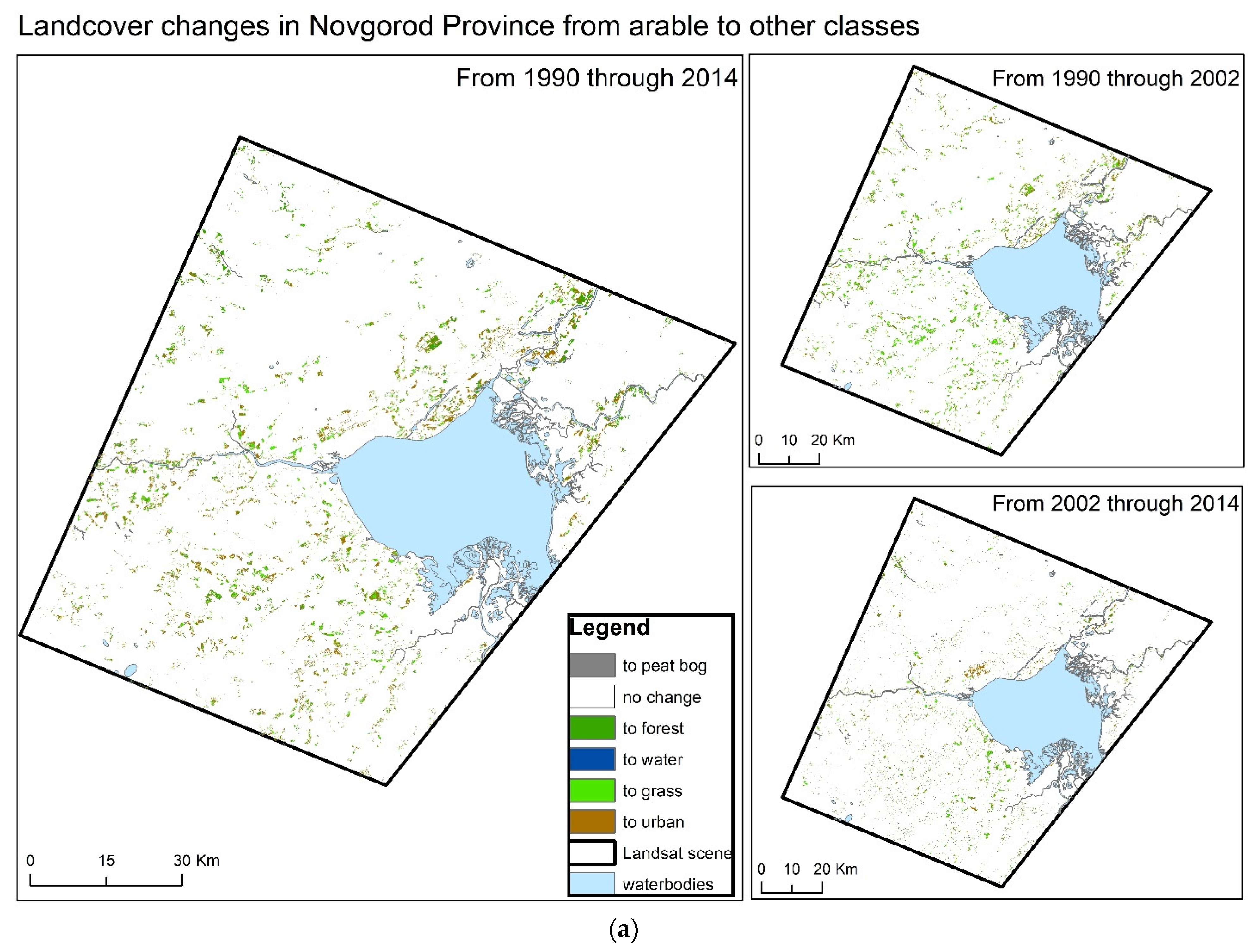{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rates | |||||||
|---|---|---|---|---|---|---|---|
| SCENE | LCC | 1990 | 2002 | 2014 | 1990–2002 | 2002–2014 | 1990–2014 |
| Yaroslav/Tver | urban | 15.05 | 18.12 | 15.92 | 0.016 | −0.011 | 0.002 |
| grass | 23.08 | 21.49 | 28.82 | −0.006 | 0.024 | 0.009 | |
| water | 48.13 | 37.68 | 63.7 | −0.020 | 0.044 | 0.012 | |
| forest | 40.65 | 41.25 | 41.36 | 0.001 | 0.000 | 0.001 | |
| arable | 13.83 | 11.72 | 5.71 | −0.014 | −0.060 | −0.037 | |
| bog | 10.24 | 11.3 | 9.91 | 0.008 | −0.015 | −0.003 | |
| Novgorod | urban | 1.71 | 2.39 | 2.88 | 0.028 | 0.016 | 0.022 |
| grass | 3.92 | 2.99 | 1.96 | −0.023 | −0.035 | −0.029 | |
| water | 2.56 | 2.63 | 2.33 | 0.002 | −0.010 | −0.004 | |
| forest | 8.83 | 9.8 | 10.68 | 0.009 | 0.007 | 0.008 | |
| arable | 7.85 | 4.91 | 6.59 | −0.039 | 0.025 | −0.007 | |
| bog | 1.95 | 1.46 | 1.25 | −0.024 | −0.013 | −0.019 | |
| combined | urban | 16.76 | 20.51 | 18.8 | 0.017 | −0.007 | 0.005 |
| grass | 27.00 | 24.48 | 30.78 | −0.008 | 0.019 | 0.005 | |
| water | 50.69 | 40.31 | 66.03 | −0.019 | 0.041 | 0.011 | |
| forest | 49.48 | 51.05 | 52.04 | 0.003 | 0.002 | 0.002 | |
| arable | 21.68 | 16.63 | 12.3 | −0.022 | −0.025 | −0.024 | |
| bog | 12.19 | 12.76 | 11.16 | 0.004 | −0.011 | −0.004 |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Runge, C.A.; Martin, T.G.; Possingham, H.P.; Willis, S.G.; Fuller, R.A. Conserving mobile species. Front. Ecol. Environ. 2014, 12, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Alerstam, T. Bird Migration; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Sutherland, W.J. Evidence for flexibility and constraint in migration systems. J. Avian Biol. 1988, 29, 441–446. [Google Scholar] [CrossRef]
- Xu, Y.; Si, Y.; Wang, Y.; Zhang, Y.; Prins, H.H.T.; Cao, L.; de Boer, W.F. Loss of functional connectivity in migration networks induces population decline in migratory birds. Ecol. Appl. 2019, 29, e01960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Si, Y.; Takekawa, J.; Liu, Q.; Prins, H.H.T.; Yin, S.; Prosser, D.J.; Gong, P.; De Boer, W.F. A network approach to prioritize conservation efforts for migratory birds. Conserv. Biol. 2020, 34, 416–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, J.S.; Stattersfield, A.J.; Butchart, S.H.; Evans, M.I.; Grimmett, R.F.; Jones, V.R.; O’Sullivan, J.; Tucker, G.M.; Newton, I. Key conservation issues for migratory land-and waterbird species on the world’s major flyways. Bird Conserv. Int. 2008, 18, S49–S73. [Google Scholar] [CrossRef] [Green Version]
- Bairlein, F. Migratory birds under threat. Science 2016, 354, 547–548. [Google Scholar] [CrossRef] [PubMed]
- Mooij, J.H. Was the (Lesser) Snow Goose (Anser c. caerulescens) once widespread in Eurasia? Goose Bull. 2022, 28, 27–42. [Google Scholar]
- Harrington, B.; Perry, E. Important Shorebird Staging Sites Meeting Western Hemisphere Shorebird Reserve Network Criteria in the United States; US Fish & Wildlife Service, Department of the Interior: Washington, DC, USA, 1995. [Google Scholar]
- McKellar, A.E.; Aubry, Y.; Drever, M.C.; Friis, C.A.; Gratto-Trevor, C.L.; Paquet, J.; Pekarik, C.; Smith, P.A. Potential Western Hemisphere Shorebird Reserve Network sites in Canada: 2020 update. Wader Study 2020, 127, 102–112. [Google Scholar] [CrossRef]
- Zhang, S.; Na, X.; Kong, B.; Wang, Z.; Jiang, H.; Yu, H.; Zhao, Z.; Li, X.; Liu, C.; Dale, P. Identifying wetland change in China’s Sanjiang Plain using remote sensing. Wetlands 2009, 29, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Lei, J.; Jia, Y.; Zuo, A.; Zeng, Q.; Shi, L.; Zhou, Y.; Zhang, H.; Lu, C.; Lei, G.; Wen, L. Bird satellite tracking revealed critical protection gaps in East Asian–Australasian Flyway. Int. J. Environ. Res. Public Health 2019, 16, 1147. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Si, Y.; Ji, L.; Gong, P. Dynamic response of East Asian Greater White-fronted Geese to changes of environment during migration: Use of multi-temporal species distribution model. Ecol. Model. 2017, 360, 70–79. [Google Scholar] [CrossRef]
- Prins, H.H.T.; Namgail, T. (Eds.) Bird Migration across the Himalayas: Wetland Functioning amidst Mountains and Glaciers; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Arzel, C.; Elmberg, J.; Guillemain, M. Ecology of spring-migrating Anatidae: A review. J. Ornithol. 2006, 147, 167–184. [Google Scholar] [CrossRef]
- Drent, R.H.; Eichhorn, G.; Flagstad, A.; Van der Graaf, A.J.; Litvin, K.E.; Stahl, J. Migratory connectivity in Arctic geese: Spring stopovers are the weak links in meeting targets for breeding. J. Ornithol. 2007, 148, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Fox, A.D.; Ebbinge, B.S.; Mitchell, C.; Heinicke, T.; Aarvak, T.; Colhoun, K.; Clausen, P.; Dereliev, S.; Faragó, S.; Koffijberg, K.; et al. Current estimates of goose population sizes in western Europe, a gap analysis and an assessment of trends. Ornis Svec. 2010, 20, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Koffijberg, K.; van Winden, E. Naar een Populatiemodel voor de Kolgans; Radboud Universiteit Nijmegen, NIOO & Sovon: Nijmegen, The Netherlands; Wageningen, The Netherlands, 2014. [Google Scholar]
- Madsen, J.; Pihl, S.; Clausen, P. Establishing a reserve network for waterfowl in Denmark: A biological evaluation of needs and consequences. Biol. Conserv. 1998, 85, 241–255. [Google Scholar] [CrossRef]
- Si, Y.; Skidmore, A.K.; Wang, T.; De Boer, W.F.; Toxopeus, A.G.; Schlerf, M.; Oudshoorn, M.; Zwerver, S.; Van Der Jeugd, H.; Exo, K.M.; et al. Distribution of Barnacle Geese Branta leucopsis in relation to food resources, distance to roosts, and the location of refuges. Ardea 2011, 99, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Kleijn, D.; van der Hout, J.; Voslamber, B.; van Randen, Y.; Melman, T.C.P. Broedende Grauwe ganzen in Nederland: Ontwikkelingen in Landbouwkundige Schade en Factoren die Hun Ruimtegebruik Beïnvloeden (Aterra Report 2343); Alterra Wageningen UR: Wageningen, The Netherlands, 2012. [Google Scholar]
- Baveco, J.M.; Kleijn, D.; de Lange, H.J.; Lammertsma, D.R.; Voslamber, B.; Melman, T.C. Populatiemodel voor de Grauwe Gans: Enkele Scenarioberekeningen voor Aantalsregulatie (No. 2445); Alterra: Wageningen, The Netherlands, 2013. [Google Scholar]
- Chudzińska, M.E.; van Beest, F.M.; Madsen, J.; Nabe-Nielsen, J. Using habitat selection theories to predict the spatiotemporal distribution of migratory birds during stopover–a case study of pink-footed geese Anser brachyrhynchus. Oikos 2015, 124, 851–860. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L. Factors Affecting Field Use of Large Grazing Birds: A Review. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2017. Available online: https://pub.epsilon.slu.se/14006/11/nilsson_l_170207.pdf (accessed on 31 January 2023).
- Grishchenko, M.; Prins, H.H.T.; Ydenberg, R.C.; Schaepman, M.E.; de Boer, W.F.; de Knegt, H.J. Land use change and the migration geography of Greater White-fronted geese in European Russia. Ecosphere 2019, 10, e02754. [Google Scholar] [CrossRef] [Green Version]
- McShea, W.J.; Underwood, H.B.; Rappole, J.H. (Eds.) The Science of Overabundance: Deer Ecology and Population Management; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2003. [Google Scholar]
- Abraham, K.F.; Jefferies, R.L.; Rockwell, R.F. Goose-induced changes in vegetation and land cover between 1976 and 1997 in an Arctic coastal marsh. Arct. Antarct. Alp. Res. 2005, 37, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Czech, H.A.; Parsons, K.C. Agricultural wetlands & waterbirds: A review. Waterbirds 2002, 25, 56–65. [Google Scholar]
- Van Eerden, M.R.; Drent, R.H.; Stahl, J.; Bakker, J.P. Connecting seas: Western Palaearctic continental flyway for water birds in the perspective of changing land use and climate. Glob. Change Biol. 2005, 11, 894–908. [Google Scholar] [CrossRef]
- Davis, J.B.; Guillemain, M.; Kaminski, R.M.; Arzel, C.; Eadie, J.M.; Rees, E.C. Habitat and resource use by waterfowl in the northern hemisphere in autumn and winter. Wildfowl 2014, 17–69. [Google Scholar]
- Kim, M.K.; Sang, I.L.; Lee, S.D. Habitat use and its implications for the conservation of the overwintering populations of Bean Goose Anser fabalis and Greater White-fronted Goose A. albifrons in South Korea. Ornithol. Sci. 2016, 15, 141–149. [Google Scholar] [CrossRef]
- Jefferies, R.L.; Rockwell, R.F.; Abraham, K.F. The embarrassment of riches: Agricultural food subsidies, high goose numbers, and loss of Arctic wetlands a continuing saga. Environ. Rev. 2004, 11, 193–232. [Google Scholar] [CrossRef] [Green Version]
- Calvert, A.M.; Gauthier, G.; Reed, A. Spatiotemporal heterogeneity of greater snow goose harvest and implications for hunting regulations. J. Wildl. Manag. 2005, 69, 561–573. [Google Scholar] [CrossRef]
- Ankney, C.D. An embarrassment of riches: Too many geese. J. Wildl. Manag. 1996, 60, 217–223. [Google Scholar] [CrossRef]
- Jefferies, R.L.; Rockwell, R.F.; Abraham, K.F. Agricultural food subsidies, migratory connectivity and large-scale disturbance in arctic coastal systems: A case study. Integr. Comp. Biol. 2004, 44, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Owen, M.; Black, J.M. Geese and their future fortune. Ibis 1991, 133, 28–35. [Google Scholar] [CrossRef]
- Buitendijk, N.H.; de Jager, M.; Hornman, M.; Kruckenberg, H.; Kölzsch, A.; Moonen, S.; Nolet, B.A. More grazing, more damage? Assessed yield loss on agricultural grassland relates nonlinearly to goose grazing pressure. J. Appl. Ecol. 2022, 59, 2878–2889. [Google Scholar] [CrossRef]
- Kerbes, R.H.; Kotanen, P.M.; Jefferies, R.L. Destruction of wetland habitats by lesser snow geese: A keystone species on the west coast of Hudson Bay. J. Appl. Ecol. 1990, 27, 242–258. [Google Scholar] [CrossRef]
- Gauthier, G.; Bêty, J.; Giroux, J.F.; Rochefort, L. Trophic interactions in a high arctic snow goose colony. Integr. Comp. Biol. 2004, 44, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegren, S.K. Rural migration and agrarian reform in Russia: A research note. Eur.-Asia Stud. 1995, 47, 877–888. [Google Scholar] [CrossRef]
- Ioffe, G.; Nefedova, T.; Zaslavsky, I. From spatial continuity to fragmentation: The case of Russian farming. Ann. Assoc. Am. Geogr. 2004, 94, 913–943. [Google Scholar]
- Grishchenko, M. Land Use Changes in Russia and Their Impact on Migrating Geese. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2018. Available online: https://edepot.wur.nl/446108 (accessed on 31 January 2023).
- Grishchenko, M.; Prins, H.H.T. Abandoned field succession in Russia and its potential effect on Corncrake Crex crex habitats. Vogelwelt 2016, 136, 175–184. [Google Scholar]
- Queiroz, C.; Beilin, R.; Folke, C.; Lindborg, R. Farmland abandonment: Threat or opportunity for biodiversity conservation? A global review. Front. Ecol. Environ. 2014, 12, 288–296. [Google Scholar] [CrossRef]
- Poschlod, P.; Bakker, J.P.; Kahmen, S. Changing land use and its impact on biodiversity. Basic Appl. Ecol. 2005, 6, 93–98. [Google Scholar] [CrossRef]
- Pal, M. Random forest classifier for remote sensing classification. Int. J. Remote Sens. 2005, 26, 217–222. [Google Scholar] [CrossRef]
- Rodriguez-Galiano, V.F.; Ghimire, B.; Rogan, J.; Chica-Olmo, M.; Rigol-Sanchez, J.P. An assessment of the effectiveness of a random forest classifier for land-cover classification. ISPRS J. Photogramm. Remote Sens. 2012, 67, 93–104. [Google Scholar] [CrossRef]
- Madsen, J. Impact of disturbance on field utilization of pink-footed geese in West Jutland, Denmark. Biol. Conserv. 1985, 33, 53–63. [Google Scholar] [CrossRef]
- Fox, A.D.; Leafloor, J.O. (Eds.) A Global Audit of the Status and Trends of Arctic and Northern Hemisphere Goose Populations; Conservation of Arctic Flora and Fauna International Secretariat: Akureyri, Iceland, 2018. [Google Scholar]
- Smith, P.A.; McKinnon, L.; Meltofte, H.; Lanctot, R.B.; Fox, A.D.; Leafloor, J.O.; Soloviev, M.; Franke, A.; Falk, K.; Golovatin, M.; et al. Status and trends of tundra birds across the circumpolar Arctic. Ambio 2020, 49, 732–748. [Google Scholar] [CrossRef] [Green Version]
- Prishchepov, A.V.; Müller, D.; Dubinin, M.; Baumann, M.; Radeloff, V.C. Determinants of agricultural land abandonment in post-Soviet European Russia. Land Use Policy 2013, 30, 873–884. [Google Scholar] [CrossRef] [Green Version]
- Kuemmerle, T.; Kaplan, J.O.; Prishchepov, A.V.; Rylsky, I.; Chaskovskyy, O.; Tikunov, V.S.; Müller, D. Forest transitions in Eastern Europe and their effects on carbon budgets. Glob. Change Biol. 2015, 21, 3049–3061. [Google Scholar] [CrossRef] [PubMed]
- Kölzsch, A.; Müskens, G.J.; Kruckenberg, H.; Glazov, P.; Weinzierl, R.; Nolet, B.A.; Wikelski, M. Towards a new understanding of migration timing: Slower spring than autumn migration in geese reflects different decision rules for stopover use and departure. Oikos 2016, 125, 1496–1507. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L.; Persson, H. Changes in field choice among staging and wintering geese in southwestern Scania, south Sweden. Ornis Svec. 2000, 10, 161–169. [Google Scholar] [CrossRef]
- Rosin, Z.M.; Skórka, P.; Wylegała, P.; Krąkowski, B.; Tobolka, M.; Myczko, Ł.; Sparks, T.H.; Tryjanowski, P. Landscape structure, human disturbance and crop management affect foraging ground selection by migrating geese. J. Ornithol. 2012, 153, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Ely, C.R.; Raveling, D.G. Seasonal variation in nutritional characteristics of the diet of greater white-fronted geese. J. Wildl. Manag. 2011, 75, 78–91. [Google Scholar] [CrossRef]
- Drent, R.H.; Fox, A.D.; Stahl, J. Travelling to breed. J. Ornithol. 2006, 147, 122–134. [Google Scholar] [CrossRef]
- Heuermann, N.; van Langevelde, F.; van Wieren, S.E.; Prins, H.H.T. Increased searching and handling effort in tall swards lead to a Type IV functional response in small grazing herbivores. Oecologia 2011, 166, 659–669. [Google Scholar] [CrossRef] [Green Version]
- Kokko, H.; Pöysä, H.; Lindström, J.; Ranta, E. Assessing the impact of spring hunting on waterfowl populations. Ann. Zool. Fenn. 1998, 35, 195–204. [Google Scholar]
- Kruckenberg, H.; Bellebaum, J.; Wille, V. Escape distances of staging Arctic geese along the flyway. Vogelwelt 2008, 129, 169–173. [Google Scholar]
- Panova, I.N.; Litvina, K.E.; Ebbinge, B.S.; Rosenfeld, S.B. Reasons for the reduction in the population of the western subspecies of the bean goose (Anser fabalis fabalis and Anser fabalis rossicus): What do the ringing data say? Biol. Bull. 2022, 49, 839–850. [Google Scholar] [CrossRef]
- Eichhorn, G.; Drent, R.H.; Stahl, J.; Leito, A.; Alerstam, T. Skipping the Baltic: The emergence of a dichotomy of alternative spring migration strategies in Russian barnacle geese. J. Anim. Ecol. 2009, 78, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Layton-Matthews, K.; Hansen, B.B.; Grøtan, V.; Fuglei, E.; Loonen, M.J. Contrasting consequences of climate change for migratory geese: Predation, density dependence and carryover effects offset benefits of high-arctic warming. Glob. Change Biol. 2020, 26, 642–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurvers, R.H.; Straates, K.; Ydenberg, R.C.; van Wieren, S.E.; Swierstra, P.S.; Prins, H.H. Social Information use by Barnacle Geese Branta leucopsis: An experiment revisited. Ardea 2014, 102, 173–180. [Google Scholar] [CrossRef] [Green Version]






| Coverage | ||||
|---|---|---|---|---|
| Years Abandoned | Grass | Herbs | Shrubs | Trees |
| up to 2 years | ~95% | ------ | 5% or less | 0% |
| 3–5 years | up to 50% | remainder | 5% | |
| up to 10 years | 10–30% | 70–90% | ||
| 11–15 years | 10% | up to 90% | 10% | |
| 16–20 years | none | up to 70% | up to 30% | |
| % of Total Area | % Change in Area | Annual Rate | ||||
|---|---|---|---|---|---|---|
| SCENE | STOPOVER | SCENE | STOPOVER | SCENE | STOPOVER | |
| urban | 9.43% | 22.33% | 12.20% | 49.70% | 0.005 | 0.017 |
| grass | 15.19% | 18.17% | 14.00% | 12.50% | 0.005 | 0.005 |
| water | 28.51% | 7.45% | 30.30% | 0.20% | 0.011 | 0 |
| forest | 27.83% | 27.55% | 5.20% | 11.70% | 0.002 | 0.005 |
| arable | 12.19% | 15.74% | −43.30% | −47.20% | −0.024 | −0.027 |
| bog | 6.86% | 8.74% | −8.40% | −19.60% | −0.004 | −0.009 |
| Rates | |||||||
|---|---|---|---|---|---|---|---|
| SCENE | LCC | 1990 | 2002 | 2014 | 1990–2002 | 2002–2014 | 1990–2014 |
| Yaroslav/Tver | urban | 4.03 | 5.4 | 4.26 | 0.024 | −0.020 | 0.002 |
| grass | 6.33 | 5.59 | 7.86 | −0.010 | 0.028 | 0.009 | |
| water | 2.47 | 2.03 | 2.55 | −0.016 | 0.019 | 0.001 | |
| forest | 9.76 | 10.34 | 10.58 | 0.005 | 0.002 | 0.003 | |
| arable | 3.77 | 2.88 | 1.61 | −0.022 | −0.048 | −0.035 | |
| bog | 3.16 | 3.26 | 2.65 | 0.003 | −0.017 | −0.007 | |
| Novgorod | urban | 5.35 | 6.91 | 9.78 | 0.021 | 0.029 | 0.025 |
| grass | 1.30 | 1.04 | 0.72 | −0.019 | −0.031 | −0.025 | |
| water | 0.66 | 0.69 | 0.587 | 0.004 | −0.013 | −0.005 | |
| forest | 1.81 | 2.07 | 2.34 | 0.011 | 0.010 | 0.011 | |
| arable | 2.84 | 1.72 | 1.88 | −0.042 | 0.007 | −0.017 | |
| bog | 0.52 | 0.45 | 0.31 | −0.012 | −0.031 | −0.022 | |
| combined | urban | 9.38 | 12.31 | 14.04 | 0.023 | 0.011 | 0.017 |
| grass | 7.63 | 6.63 | 8.58 | −0.012 | 0.021 | 0.005 | |
| water | 3.13 | 2.72 | 3.137 | −0.012 | 0.012 | 0.000 | |
| forest | 11.57 | 12.41 | 12.92 | 0.006 | 0.003 | 0.005 | |
| arable | 6.61 | 4.60 | 3.49 | −0.030 | −0.023 | −0.027 | |
| bog | 3.68 | 3.71 | 2.96 | 0.001 | −0.019 | −0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grishchenko, M.; Ydenberg, R.C.; Prins, H.H.T. Large-Scale Reduction in the Extent of Agriculture around Stopover Sites of Migratory Geese in European Russia between 1990 and 2015. Diversity 2023, 15, 447. https://doi.org/10.3390/d15030447
Grishchenko M, Ydenberg RC, Prins HHT. Large-Scale Reduction in the Extent of Agriculture around Stopover Sites of Migratory Geese in European Russia between 1990 and 2015. Diversity. 2023; 15(3):447. https://doi.org/10.3390/d15030447
Chicago/Turabian StyleGrishchenko, Mikhail, Ronald C. Ydenberg, and Herbert H. T. Prins. 2023. "Large-Scale Reduction in the Extent of Agriculture around Stopover Sites of Migratory Geese in European Russia between 1990 and 2015" Diversity 15, no. 3: 447. https://doi.org/10.3390/d15030447