

Abiotic and Biotic Factors from the Past as Predictors of Alien Bird Richness and Temporal Beta-Diversity
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data
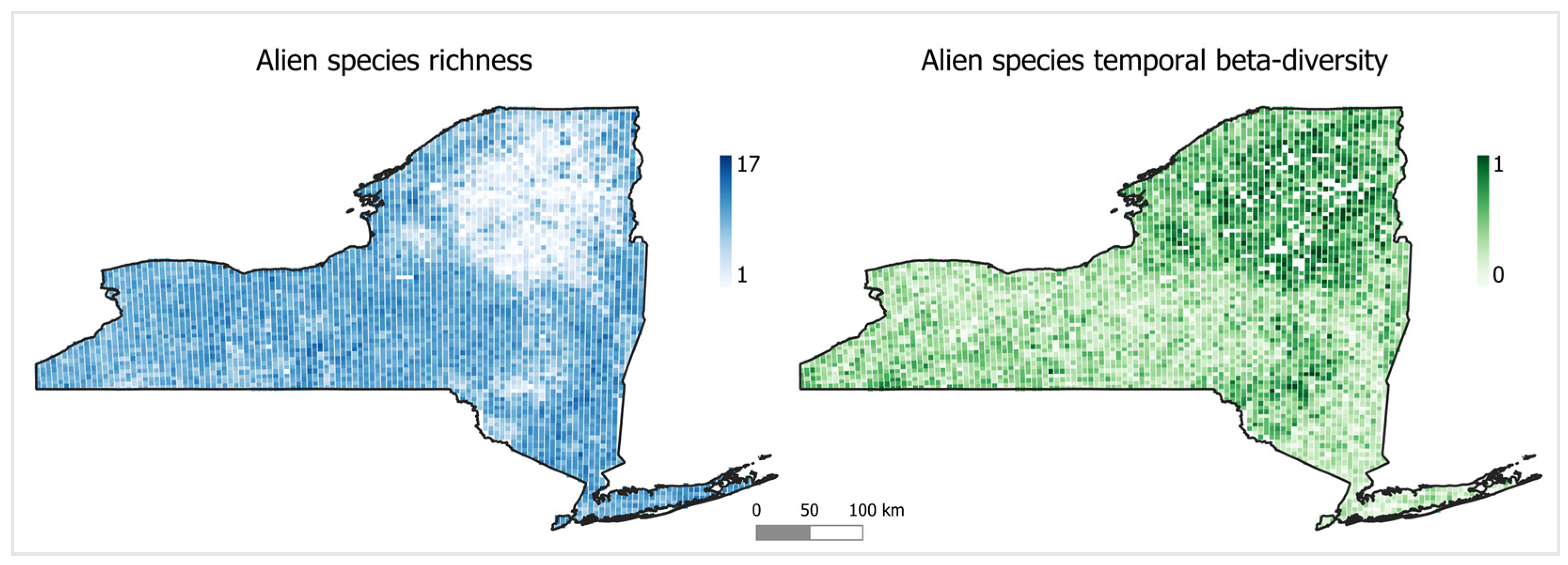
2.1.1. Alien Species Diversity
2.1.2. Biotic and Abiotic Predictors
Biotic Factors: Native Taxonomic, Functional and Phylogenetic Diversity, and Human Population Density
Abiotic Factors: Climate and Land-Use
2.2. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Hoffmann, M.; Wirsing, A.J.; McCauley, D.J. Extinction risk is most acute for the world’s largest and smallest vertebrates. Proc. Natl. Acad. Sci. USA 2017, 114, 10678–10683. [Google Scholar] [PubMed] [Green Version]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [PubMed]
- Vilà, M.; Hulme, P.E. Impact of Biological Invasions on Ecosystem Services; Springer: Berlin/Heidelberg, Germany, 2017; Volume 12. [Google Scholar]
- Ricciardi, A.; Hoopes, M.F.; Marchetti, M.P.; Lockwood, J.L. Progress toward understanding the ecological impacts of nonnative species. Ecol. Monogr. 2013, 83, 263–282. [Google Scholar]
- Powell, K.I.; Chase, J.M.; Knight, T.M. A synthesis of plant invasion effects on biodiversity across spatial scales. Am. J. Bot. 2011, 98, 539–548. [Google Scholar]
- Strayer, D.L.; Eviner, V.T.; Jeschke, J.M.; Pace, M.L. Understanding the long-term effects of species invasions. Trends Ecol. Evol. 2006, 21, 645–651. [Google Scholar]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [PubMed] [Green Version]
- Kuebbing, S.E.; Nuñez, M.A. Negative, neutral, and positive interactions among nonnative plants: Patterns, processes, and management implications. Glob. Chang. Biol. 2015, 21, 926–934. [Google Scholar]
- Schmiedel, D.; Wilhelm, E.-G.; Roth, M.; Scheibner, C.; Nehring, S.; Winter, S. Evaluation system for management measures of invasive alien species. Biodivers. Conserv. 2016, 25, 357–374. [Google Scholar]
- Stohlgren, T.J.; Barnett, D.; Flather, C.; Fuller, P.; Peterjohn, B.; Kartesz, J.; Master, L.L. Species richness and patterns of invasion in plants, birds, and fishes in the United States. Biol. Invasions 2006, 8, 427–447. [Google Scholar]
- Lembrechts, J.J.; Alexander, J.M.; Cavieres, L.A.; Haider, S.; Lenoir, J.; Kueffer, C.; McDougall, K.; Naylor, B.J.; Nuñez, M.A.; Pauchard, A. Mountain roads shift native and non-native plant species’ ranges. Ecography 2017, 40, 353–364. [Google Scholar]
- Abellán, P.; Tella, J.L.; Carrete, M.; Cardador, L.; Anadón, J.D. Climate matching drives spread rate but not establishment success in recent unintentional bird introductions. Proc. Natl. Acad. Sci. USA 2017, 114, 9385–9390. [Google Scholar] [PubMed] [Green Version]
- Redding, D.W.; Pigot, A.L.; Dyer, E.E.; Şekercioğlu, Ç.H.; Kark, S.; Blackburn, T.M. Location-level processes drive the establishment of alien bird populations worldwide. Nature 2019, 571, 103–106. [Google Scholar]
- Dukes, J.S. Biodiversity and invasibility in grassland microcosms. Oecologia 2001, 126, 563–568. [Google Scholar] [PubMed]
- Gerhold, P.; Pärtel, M.; Tackenberg, O.; Hennekens, S.M.; Bartish, I.; Schaminée, J.H.; Fergus, A.J.; Ozinga, W.A.; Prinzing, A. Phylogenetically poor plant communities receive more alien species, which more easily coexist with natives. Am. Nat. 2011, 177, 668–680. [Google Scholar]
- Iannone, B.V.; Potter, K.M.; Hamil, K.-A.D.; Huang, W.; Zhang, H.; Guo, Q.; Oswalt, C.M.; Woodall, C.W.; Fei, S. Evidence of biotic resistance to invasions in forests of the Eastern USA. Landscape Ecol. 2016, 31, 85–99. [Google Scholar]
- Ng, J.; Weaver, W.N.; Laport, R.G. Testing Darwin’s Naturalization Conundrum using phylogenetic relationships: Generalizable patterns across disparate communities? Divers. Distrib. 2019, 25, 361–373. [Google Scholar]
- Whitfeld, T.J.; Lodge, A.G.; Roth, A.M.; Reich, P.B. Community phylogenetic diversity and abiotic site characteristics influence abundance of the invasive plant Rhamnus cathartica L. J. Plant Ecol. 2013, 7, 202–209. [Google Scholar]
- Andrikou-Charitidou, A.; Boutsis, G.; Karadimou, E.; Kallimanis, A. Untangling the positive association of phylogenetic, functional, and taxonomic diversity with alien bird species richness. Ecosphere 2020, 11, e03007. [Google Scholar]
- El-Barougy, R.; MacIvor, J.S.; Arnillas, C.A.; Nada, R.M.; Khedr, A.H.A.; Cadotte, M.W. Richness, phylogenetic diversity, and abundance all have positive effects on invader performance in an arid ecosystem. Ecosphere 2020, 11, e03045. [Google Scholar]
- Loiola, P.P.; de Bello, F.; Chytrý, M.; Götzenberger, L.; Carmona, C.P.; Pyšek, P.; Lososová, Z. Invaders among locals: Alien species decrease phylogenetic and functional diversity while increasing dissimilarity among native community members. J. Ecol. 2018, 106, 2230–2241. [Google Scholar]
- de Bello, F.; Šmilauer, P.; Diniz-Filho, J.A.F.; Carmona, C.P.; Lososová, Z.; Herben, T.; Götzenberger, L. Decoupling phylogenetic and functional diversity to reveal hidden signals in community assembly. Methods Ecol. Evol. 2017, 8, 1200–1211. [Google Scholar]
- Galland, T.; Adeux, G.; Dvořáková, H.; E-Vojtkó, A.; Orbán, I.; Lussu, M.; Puy, J.; Blažek, P.; Lanta, V.; Lepš, J. Colonization resistance and establishment success along gradients of functional and phylogenetic diversity in experimental plant communities. J. Ecol. 2019, 107, 2090–2104. [Google Scholar]
- Qin, T.J.; Zhou, J.; Sun, Y.; Müller-Schärer, H.; Luo, F.L.; Dong, B.C.; Li, H.L.; Yu, F.H. Phylogenetic diversity is a better predictor of wetland community resistance to Alternanthera philoxeroides invasion than species richness. Plant Biol. 2020, 22, 591–599. [Google Scholar] [PubMed]
- Levine, J.M.; Adler, P.B.; Yelenik, S.G. A meta-analysis of biotic resistance to exotic plant invasions. Ecol. Lett. 2004, 7, 975–989. [Google Scholar]
- Helsen, K.; Van Cleemput, E.; Bassi, L.; Graae, B.J.; Somers, B.; Blonder, B.; Honnay, O. Inter-and intraspecific trait variation shape multidimensional trait overlap between two plant invaders and the invaded communities. Oikos 2020, 129, 677–688. [Google Scholar]
- Fridley, J.D.; Stachowicz, J.; Naeem, S.; Sax, D.; Seabloom, E.; Smith, M.; Stohlgren, T.; Tilman, D.; Holle, B.V. The invasion paradox: Reconciling pattern and process in species invasions. Ecology 2007, 88, 3–17. [Google Scholar]
- Bellard, C.; Leroy, B.; Thuiller, W.; Rysman, J.F.; Courchamp, F. Major drivers of invasion risks throughout the world. Ecosphere 2016, 7, e01241. [Google Scholar]
- Gallardo, B.; Aldridge, D.C. The ‘dirty dozen’: Socio-economic factors amplify the invasion potential of 12 high-risk aquatic invasive species in Great Britain and Ireland. J. Appl. Ecol. 2013, 50, 757–766. [Google Scholar]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Bacher, S.; Chiron, F.; Didžiulis, V.; Essl, F. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar] [PubMed] [Green Version]
- McKinney, M.; Kark, S. Factors shaping avian alien species richness in Australia vs Europe. Divers. Distrib. 2017, 23, 1334–1342. [Google Scholar]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [PubMed] [Green Version]
- Richardson, D.M.; Pyšek, P.; Carlton, J.T. A compendium of essential concepts and terminology in invasion ecology. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; John Wiley & Sons Ltd.: Chichester, UK, 2011; pp. 409–420. [Google Scholar]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar]
- Gallardo, B. Europe’s top 10 invasive species: Relative importance of climatic, habitat and socio-economic factors. Ethol. Ecol. Evol. 2014, 26, 130–151. [Google Scholar]
- Qian, H.; Ricklefs, R.E. The role of exotic species in homogenizing the North American flora. Ecol. Lett. 2006, 9, 1293–1298. [Google Scholar]
- Spear, D.; Foxcroft, L.C.; Bezuidenhout, H.; McGeoch, M.A. Human population density explains alien species richness in protected areas. Biol. Conserv. 2013, 159, 137–147. [Google Scholar]
- Lazarina, M.; Tsianou, M.A.; Boutsis, G.; Andrikou–Charitidou, A.; Karadimou, E.; Kallimanis, A.S. Urbanization and Human Population Favor Species Richness of Alien Birds. Diversity 2020, 12, 72. [Google Scholar]
- Chiron, F.; Shirley, S.; Kark, S. Human-related processes drive the richness of exotic birds in Europe. Proc. R. Soc. B Biol. Sci. 2009, 276, 47–53. [Google Scholar]
- Marini, L.; Bertolli, A.; Bona, E.; Federici, G.; Martini, F.; Prosser, F.; Bommarco, R. Beta-diversity patterns elucidate mechanisms of alien plant invasion in mountains. Global Ecol. Biogeogr. 2013, 22, 450–460. [Google Scholar]
- Latombe, G.; Pyšek, P.; Jeschke, J.M.; Blackburn, T.M.; Bacher, S.; Capinha, C.; Costello, M.J.; Fernández, M.; Gregory, R.D.; Hobern, D. A vision for global monitoring of biological invasions. Biol. Conserv. 2017, 213, 295–308. [Google Scholar] [CrossRef]
- Gavioli, A.; Milardi, M.; Castaldelli, G.; Fano, E.A.; Soininen, J. Diversity patterns of native and exotic fish species suggest homogenization processes, but partly fail to highlight extinction threats. Divers. Distrib. 2019, 25, 983–994. [Google Scholar] [CrossRef] [Green Version]
- Leprieur, F.; Olden, J.D.; Lek, S.; Brosse, S. Contrasting patterns and mechanisms of spatial turnover for native and exotic freshwater fish in Europe. J. Biogeogr. 2009, 36, 1899–1912. [Google Scholar] [CrossRef]
- Olden, J.D.; Rooney, T.P. On defining and quantifying biotic homogenization. Glob. Ecol. Biogeogr. 2006, 15, 113–120. [Google Scholar] [CrossRef]
- Olden, J.D.; Poff, N.L. Toward a mechanistic understanding and prediction of biotic homogenization. Am. Nat. 2003, 162, 442–460. [Google Scholar] [CrossRef] [Green Version]
- Kueffer, C.; Pyšek, P.; Richardson, D.M. Integrative invasion science: Model systems, multi-site studies, focused meta-analysis and invasion syndromes. New Phytol. 2013, 200, 615–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebens, H.; Bacher, S.; Blackburn, T.M.; Capinha, C.; Dawson, W.; Dullinger, S.; Genovesi, P.; Hulme, P.E.; van Kleunen, M.; Kühn, I. Projecting the continental accumulation of alien species through to 2050. Glob. Chang. Biol. 2021, 27, 970–982. [Google Scholar] [CrossRef]
- Cohen, T.M.; McKinney, M.; Kark, S.; Dor, R. Global invasion in progress: Modeling the past, current and potential global distribution of the common myna. Biol. Invasions 2019, 21, 1295–1309. [Google Scholar] [CrossRef]
- Andrle, R.F.; Carroll, J.R. The Atlas of Breeding Birds in New York State; Cornell University Press: New York, NY, USA, 1988. [Google Scholar]
- McGowan, K.J.; Corwin, K. The Second Atlas of Breeding Birds in New York State; Cornell University Press: New York, NY, USA, 2008. [Google Scholar]
- Chesser, R.; Burns, K.; Cicero, C.; Dunn, J.; Kratter, A.; Lovette, I.; Rasmussen, P.; Remsen, J., Jr.; Stotz, D.; Winger, B. Checklist of North American Birds (Online). American Ornithological Society. 2018. Available online: https://checklist.americanornithology.org/taxa (accessed on 15 July 2018).
- Dyer, E.E.; Redding, D.W.; Blackburn, T.M. The global avian invasions atlas, a database of alien bird distributions worldwide. Sci. Data 2017, 4, 170041. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E.; Dornelas, M.; Moyes, F.; Henderson, P.A. Temporal β diversity—A macroecological perspective. Glob. Ecol. Biogeogr. 2019, 28, 1949–1960. [Google Scholar] [CrossRef]
- Baselga, A.; Orme, D.; Villeger, S.; De Bortoli, J.; Leprieur, F.; Baselga, M.A. Package ‘betapart’. Partitioning Beta Diversity Into Turnover Nestedness Components 2018. Available online: http://cran.nexr.com/web/packages/betapart/betapart.pdf (accessed on 17 September 2020).
- R Core Team. R: A Language and Environment for Statistical Computing. 2013. Available online: https://www.scirp.org/(S(i43dyn45teexjx455qlt3d2q))/reference/ReferencesPapers.aspx?ReferenceID=1787696 (accessed on 17 September 2020).
- Legendre, P. A temporal beta-diversity index to identify sites that have changed in exceptional ways in space–time surveys. Ecol. Evol. 2019, 9, 3500–3514. [Google Scholar] [CrossRef] [Green Version]
- Barnagaud, J.-Y.; Gaüzère, P.; Zuckerberg, B.; Princé, K.; Svenning, J.-C. Temporal changes in bird functional diversity across the United States. Oecologia 2017, 185, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–871. [Google Scholar] [CrossRef]
- Gower, J.C. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Mason, N.W.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B.; Laliberté, M.E. Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. R Package FD 2014. Available online: https://www.scirp.org/(S(i43dyn45teexjx455qlt3d2q))/reference/ReferencesPapers.aspx?ReferenceID=1820451 (accessed on 17 September 2020).
- Jetz, W.; Thomas, G.; Joy, J.; Hartmann, K.; Mooers, A. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef] [Green Version]
- Price, C.V.; Nakagaki, N.; Hitt, K.J.; Clawges, R. Enhanced Historical Land-Use and Land-Cover Data Sets of the US Geological Survey; US Department of the Interior, US Geological Survey: Washington, DC, USA, 2007.
- Center for International Earth Science Information Network—CIESIN—Columbia University. Global Population Density Grid Time Series Estimates; NASA Socioeconomic Data and Applications Center (SEDAC): Palisades, NY, USA, 2017. [CrossRef]
- Yatish, S.; Jiarpakdee, J.; Thongtanunam, P.; Tantithamthavorn, C. Mining Software Defects: Should We Consider Affected Releases? In Proceedings of the International Conference on Software Engineering (ICSE), Montreal, QC, Canada, 25–31 May 2019. [Google Scholar]
- Queen, J.P.; Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Pinheiro, J.; Bates, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-162. 2023. Available online: https://CRAN.R-project.org/package=nlme (accessed on 17 September 2020).
- Groemping, U.; Matthias, L. Package ‘relaimpo’. Relative Importance of Regressors in Linear Models (R Package Version). 2018. Available online: https://prof.bht-berlin.de/groemping/software/relaimpo/ (accessed on 17 September 2021).
- Essl, F.; Dawson, W.; Kreft, H.; Pergl, J.; Pyšek, P.; Van Kleunen, M.; Weigelt, P.; Mang, T.; Dullinger, S.; Lenzner, B. Drivers of the relative richness of naturalized and invasive plant species on Earth. AoB Plants 2019, 11, plz051. [Google Scholar] [CrossRef] [Green Version]
- Del Grosso, S.; Parton, W.; Stohlgren, T.; Zheng, D.; Bachelet, D.; Prince, S.; Hibbard, K.; Olson, R. Global potential net primary production predicted from vegetation class, precipitation, and temperature. Ecology 2008, 89, 2117–2126. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Mooney, H.A. Invasive alien species in an era of globalization. Front. Ecol. Environ. 2007, 5, 199–208. [Google Scholar] [CrossRef]
- Essl, F.; Dullinger, S.; Rabitsch, W.; Hulme, P.E.; Hülber, K.; Jarošík, V.; Kleinbauer, I.; Krausmann, F.; Kühn, I.; Nentwig, W. Socioeconomic legacy yields an invasion debt. Proc. Natl. Acad. Sci. USA 2011, 108, 203–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coetzee, B.W.; Chown, S.L. Land-use change promotes avian diversity at the expense of species with unique traits. Ecol. Evol. 2016, 6, 7610–7622. [Google Scholar] [CrossRef] [PubMed]
- Sol, D.; Bartomeus, I.; Griffin, A.S. The paradox of invasion in birds: Competitive superiority or ecological opportunism? Oecologia 2012, 169, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Dyer, E.E.; Cassey, P.; Redding, D.W.; Collen, B.; Franks, V.; Gaston, K.J.; Jones, K.E.; Kark, S.; Orme, C.D.L.; Blackburn, T.M. The global distribution and drivers of alien bird species richness. PLoS Biol. 2017, 15, e2000942. [Google Scholar] [CrossRef] [Green Version]
- Lovell, R.S.; Blackburn, T.M.; Dyer, E.E.; Pigot, A.L. Environmental resistance predicts the spread of alien species. Nat. Ecol. Evol. 2021, 5, 322–329. [Google Scholar] [CrossRef]
- Lazarina, M.; Sgardelis, S.P.; Michailidou, D.E.; Tsianou, M.; Andrikou-Charitidou, A.; Touloumis, K.; Kallimanis, A.S. Replacement drives native β-diversity of British avifauna, while richness differences shape alien β-diversity. Divers. Distrib. 2023, 29, 61–74. [Google Scholar] [CrossRef]
- Borges, P.A.; Rigal, F.; Ros-Prieto, A.; Cardoso, P. Increase of insular exotic arthropod diversity is a fundamental dimension of the current biodiversity crisis. Insect Conserv. Divers. 2020, 13, 508–518. [Google Scholar] [CrossRef]
- Blonder, B. Do hypervolumes have holes? Am. Nat. 2016, 187, E93–E105. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Delean, S.; Pyšek, P.; Cassey, P. On the island biogeography of aliens: A global analysis of the richness of plant and bird species on oceanic islands. Glob. Ecol. Biogeogr. 2016, 25, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Encyclopedia of Life. Available online: http://eol.org (accessed on 15 July 2018).
- Myers, P.; Espinosa, R.; Parr, C.S.; Jones, T.; Hammond, G.S.; Dewey, T.A. The Animal Diversity Web. 2019. Available online: https://animaldiversity.org (accessed on 15 July 2018).
- Sibley, D.A. The Sibley Guide to Birds, 2nd ed.; Knopf: New York, NY, USA, 2014. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
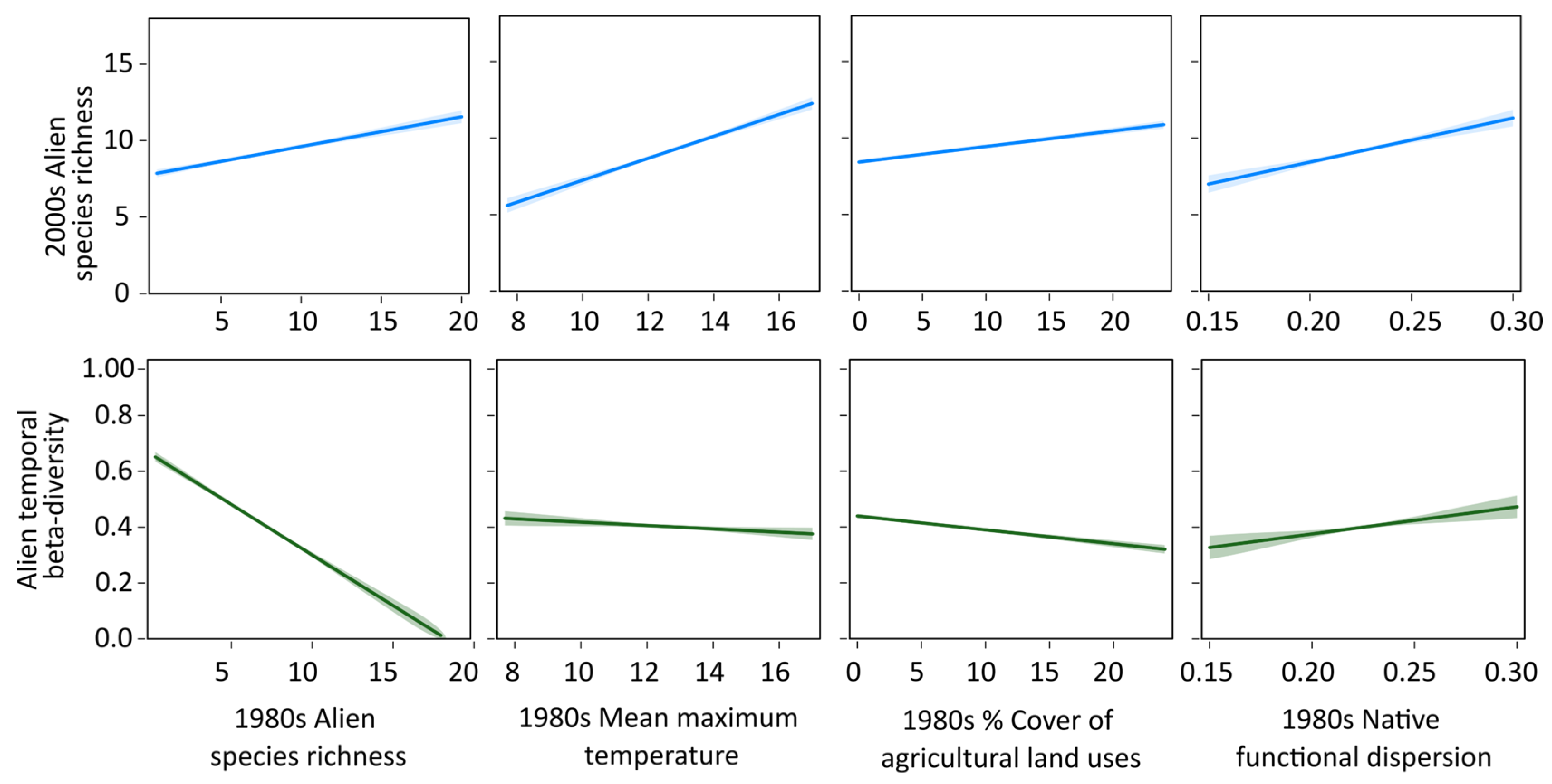
| Predictors (1980s) | Alien Species Richness (2000s) | Alien Temporal Beta-Diversity | ||
|---|---|---|---|---|
| AIC | R2 | AIC | R2 | |
| Tmax | 21,304.59 | 0.343 *** | −4199.23 | 0.258 *** |
| PPT | 21,466.01 | 0.088 *** | −3840.59 | 0.0391 *** |
| LandDiv | 21,410.64 | 0.065 *** | −3847.55 | 0.0192 *** |
| Developed | 21,526.44 | 0.036 *** | −3861.10 | 0.0599 *** |
| Agricultural | 21,448.51 | 0.232 *** | −3932.30 | 0.134 *** |
| PopDen | 21,540.96 | 0.013 *** | −3814.484 | 0.0246 *** |
| Alien SR | 21,120.53 | 0.451 *** | −5387.39 | 0.457 *** |
| Native SR | 21,347.59 | 0.032 ** | −4025.42 | 0.040 *** |
| Native FDis | 21,411.59 | 0.1494 *** | −3936.52 | 0.114 *** |
| Native MPD | 21,500.44 | 0.048 *** | −3968.500 | 0.086 *** |
| Multivariate model | 20,554.29 | 0.569 *** | −5526.491 | 0.480 *** |
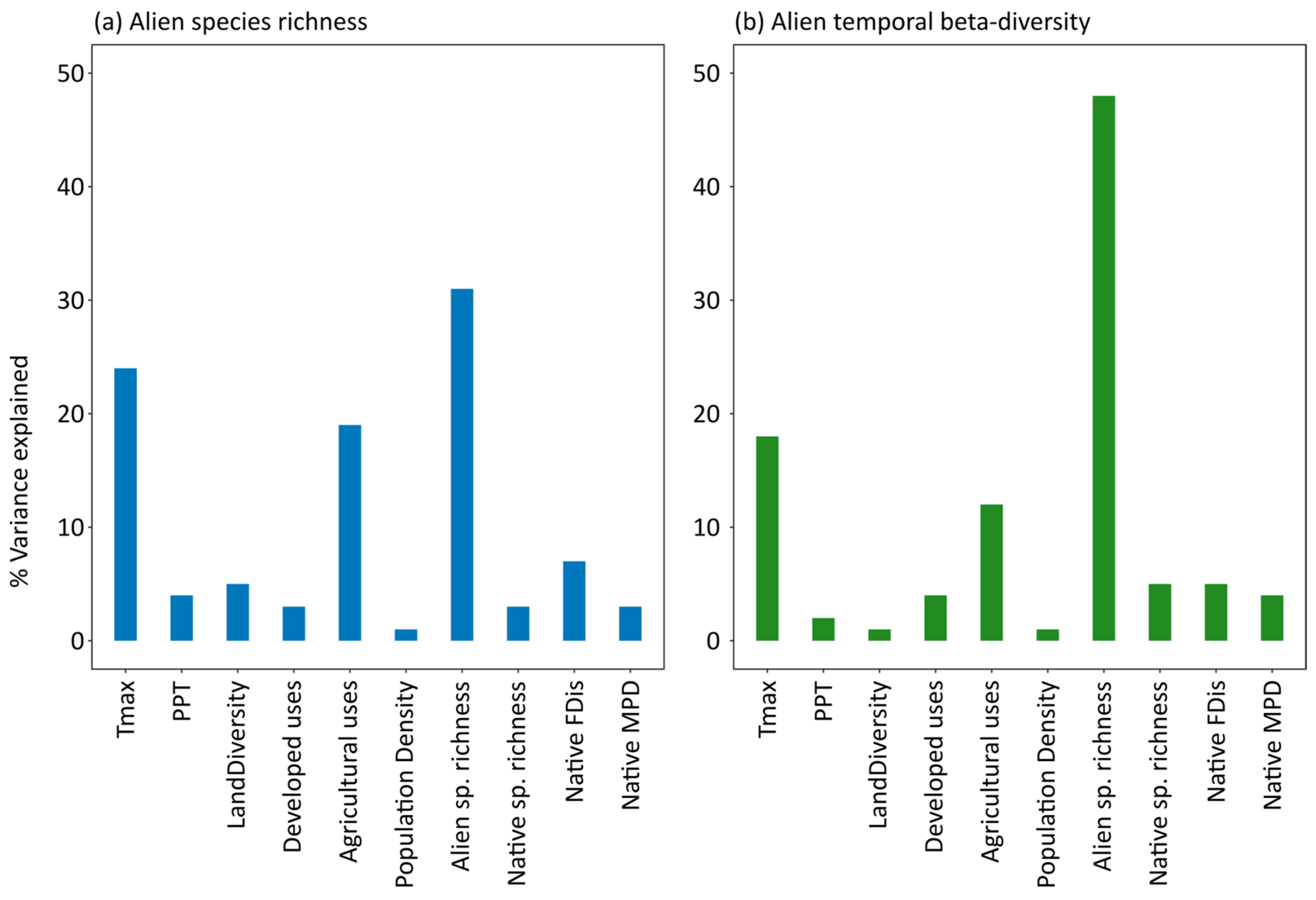
| Alien Species Richness (2000s) | Alien Temporal Beta-Diversity | |||||
|---|---|---|---|---|---|---|
| Coef. | SE | p | Coef. | SE | p | |
| (Intercept) | −3.806 | 1.004 | 0.000 | 0.727 | 0.060 | 0.000 |
| Tmax | 0.717 | 0.046 | 0.000 | −0.006 | 0.003 | 0.020 |
| PPT | −0.017 | 0.006 | 0.005 | −0.001 | 0.000 | 0.010 |
| LandDiv | 0.997 | 0.108 | 0.000 | −0.011 | 0.007 | 0.122 |
| Developed | 0.027 | 0.012 | 0.027 | −0.004 | 0.001 | 0.000 |
| Agricultural | 0.102 | 0.007 | 0.000 | −0.005 | 0.000 | 0.000 |
| PopDen | 0.000 | 0.000 | 0.562 | 0.000 | 0.000 | 0.562 |
| Alien SR | 0.194 | 0.017 | 0.000 | −0.036 | 0.001 | 0.000 |
| Native SR | 0.021 | 0.003 | 0.000 | 0.000 | 0.000 | 0.120 |
| Native FDis | 28.749 | 3.707 | 0.000 | 0.972 | 0.279 | 0.001 |
| Native MPD | −0.049 | 0.005 | 0.000 | −0.001 | 0.000 | 0.064 |
| AIC: 20,554.29, R2 = 0.5687, p < 0.005 | AIC: −5526.491, R2 = 0.4803, p < 0.005 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrikou-Charitidou, A.; Boutsis, G.; Karadimou, E.; Tsianou, M.A.; Lazarina, M.; Kallimanis, A.S. Abiotic and Biotic Factors from the Past as Predictors of Alien Bird Richness and Temporal Beta-Diversity. Diversity 2023, 15, 417. https://doi.org/10.3390/d15030417
Andrikou-Charitidou A, Boutsis G, Karadimou E, Tsianou MA, Lazarina M, Kallimanis AS. Abiotic and Biotic Factors from the Past as Predictors of Alien Bird Richness and Temporal Beta-Diversity. Diversity. 2023; 15(3):417. https://doi.org/10.3390/d15030417
Chicago/Turabian StyleAndrikou-Charitidou, Aristi, Georgios Boutsis, Elpida Karadimou, Mariana A. Tsianou, Maria Lazarina, and Athanasios S. Kallimanis. 2023. "Abiotic and Biotic Factors from the Past as Predictors of Alien Bird Richness and Temporal Beta-Diversity" Diversity 15, no. 3: 417. https://doi.org/10.3390/d15030417