

Palaeoeconomy and Palaeoenvironment of Halmyris—A Roman Settlement in Southeast Romania: Archaeozoological and Phytolith Evidences

 , , and
, , and
Abstract
:1. Introduction
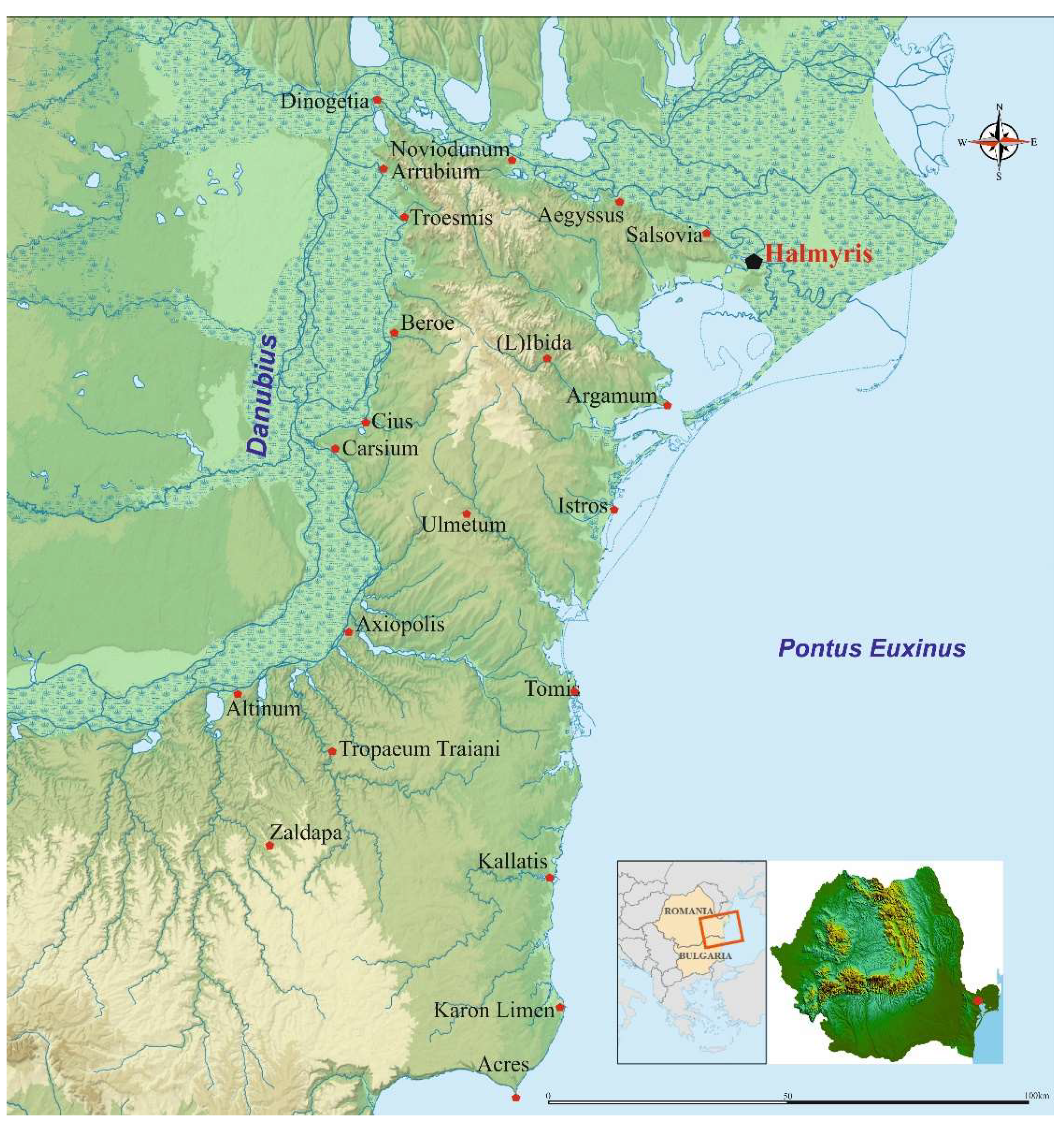
2. Study Area and the Archaeological Context
2.1. Study Area
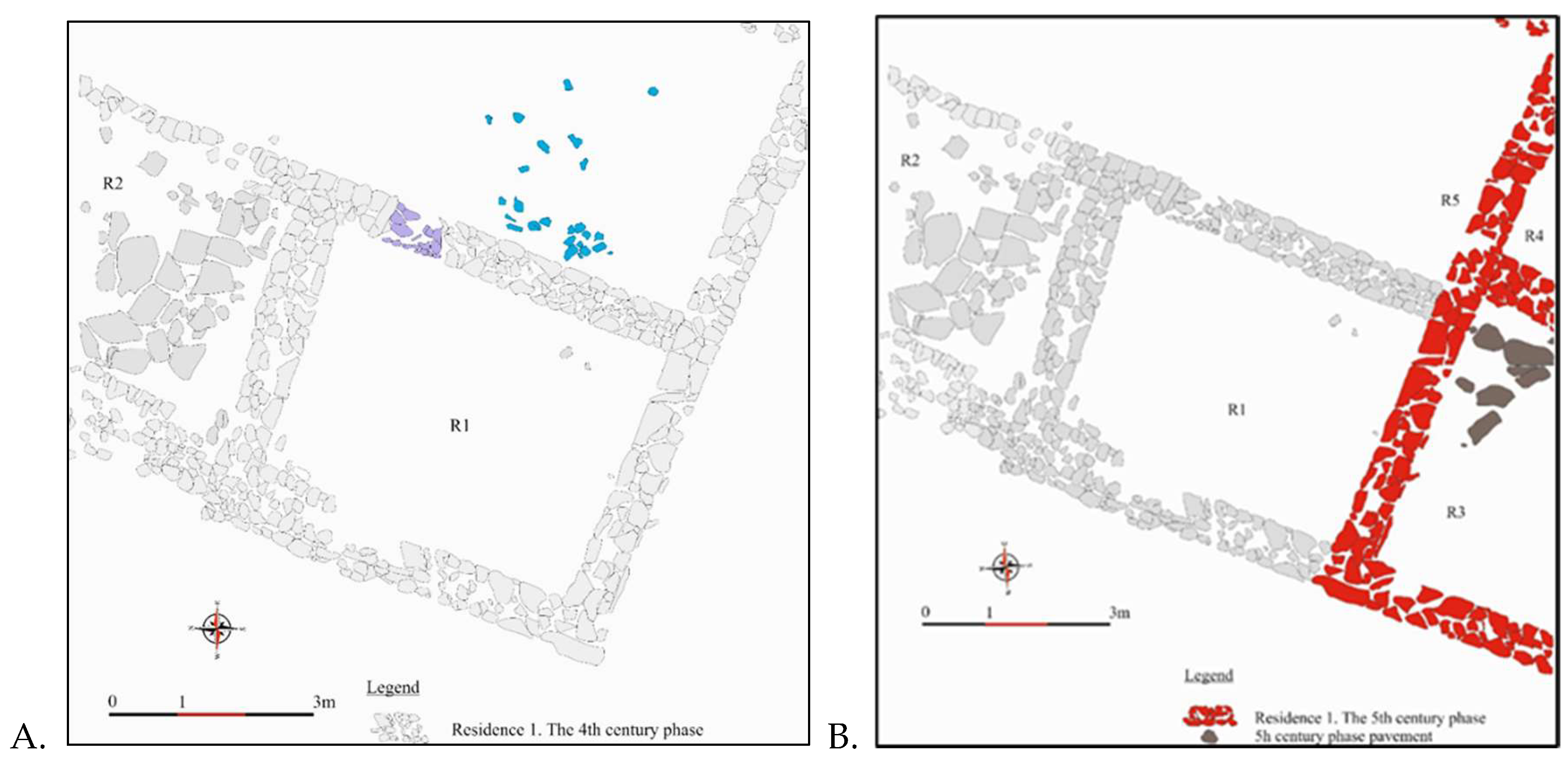
2.2. Archaeological Context
3. Materials and Methods
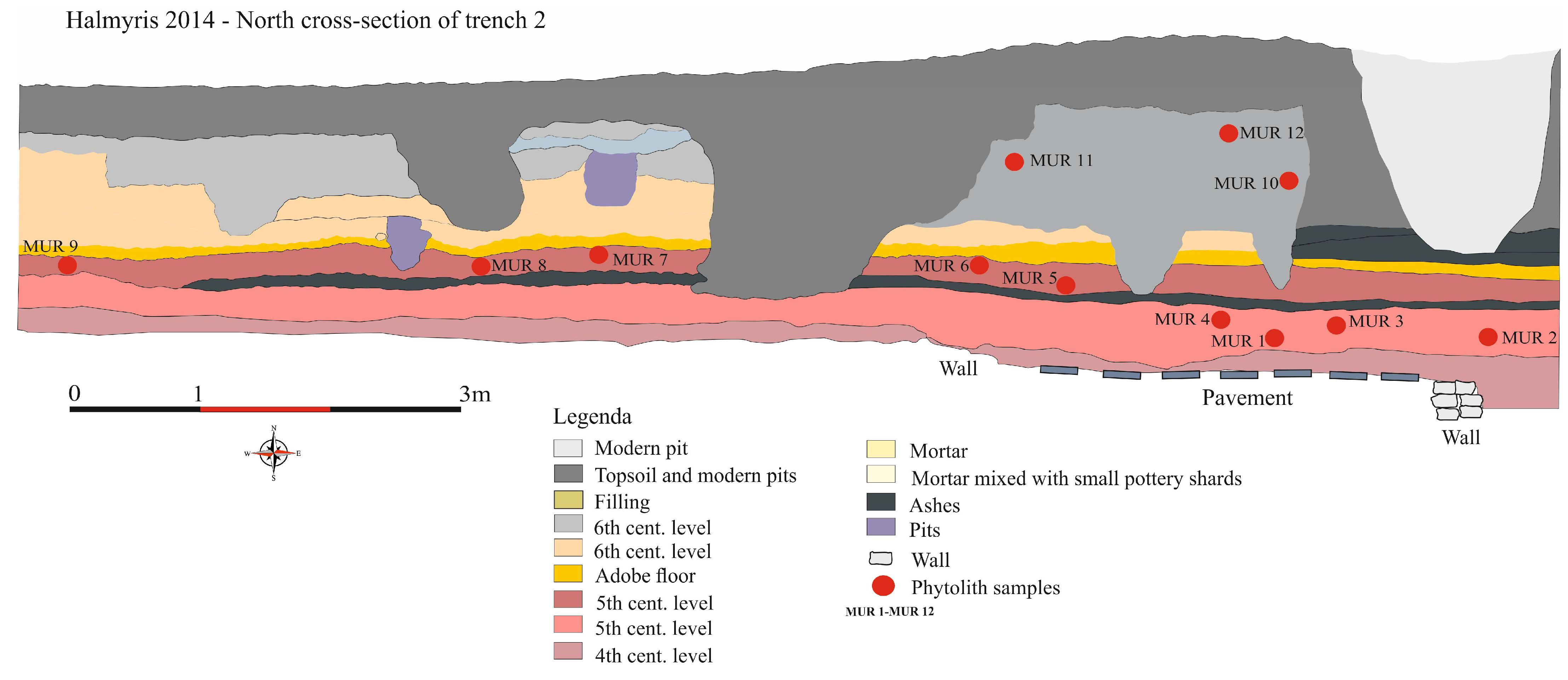
3.1. Phytolith analysis
3.2. Archaeozoology
4. Results and Discussion
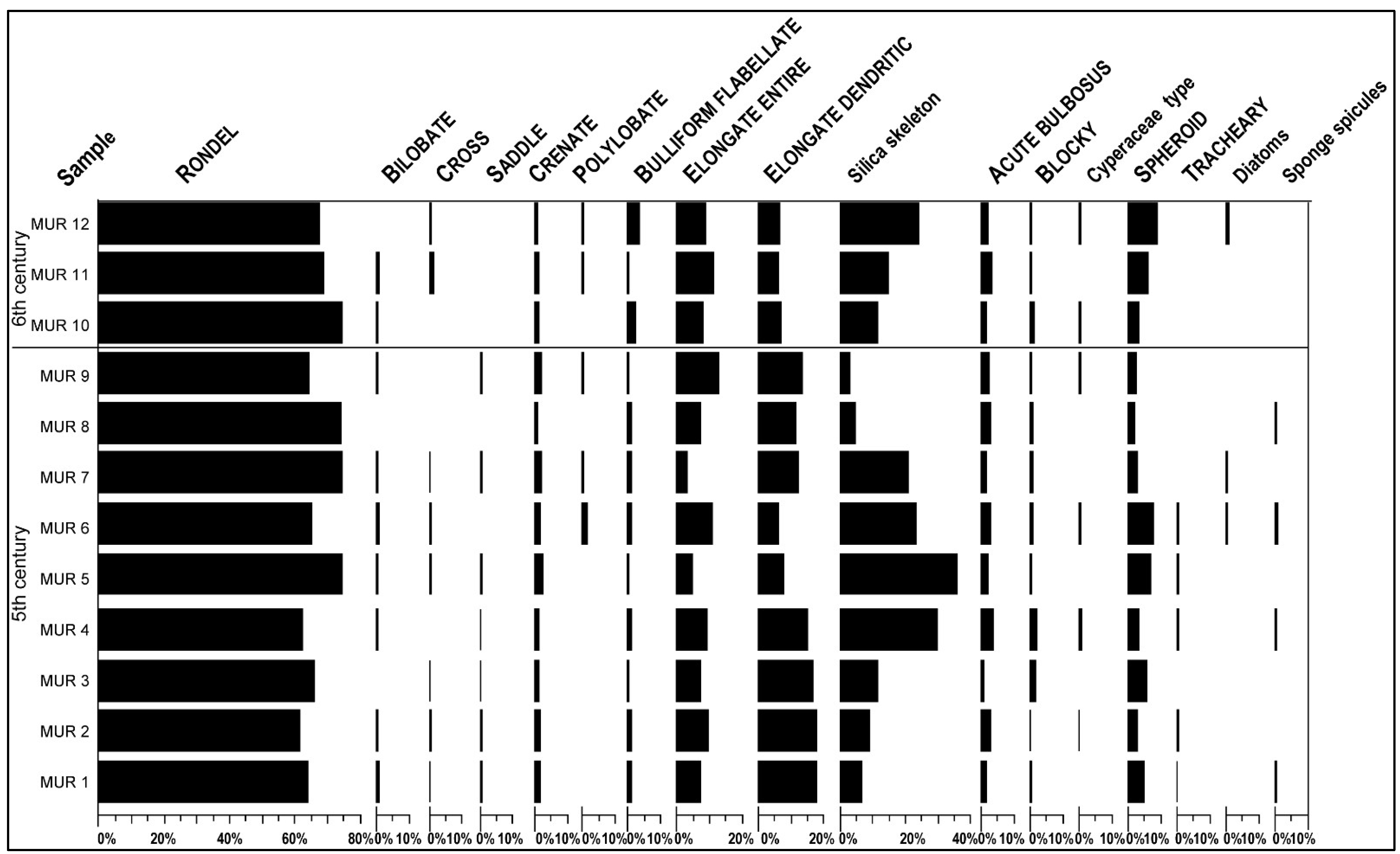
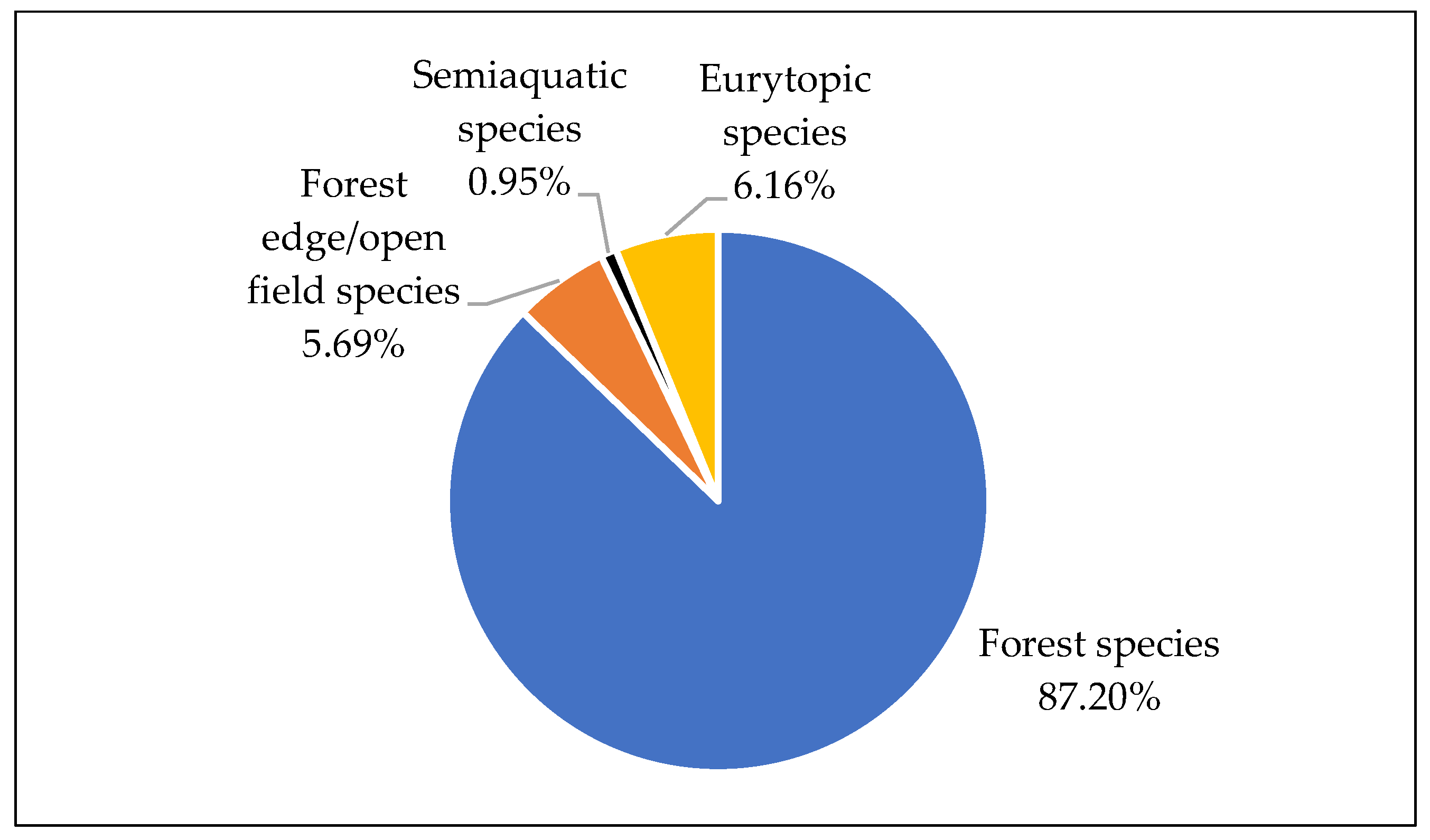
4.1. Phytolith Analysis
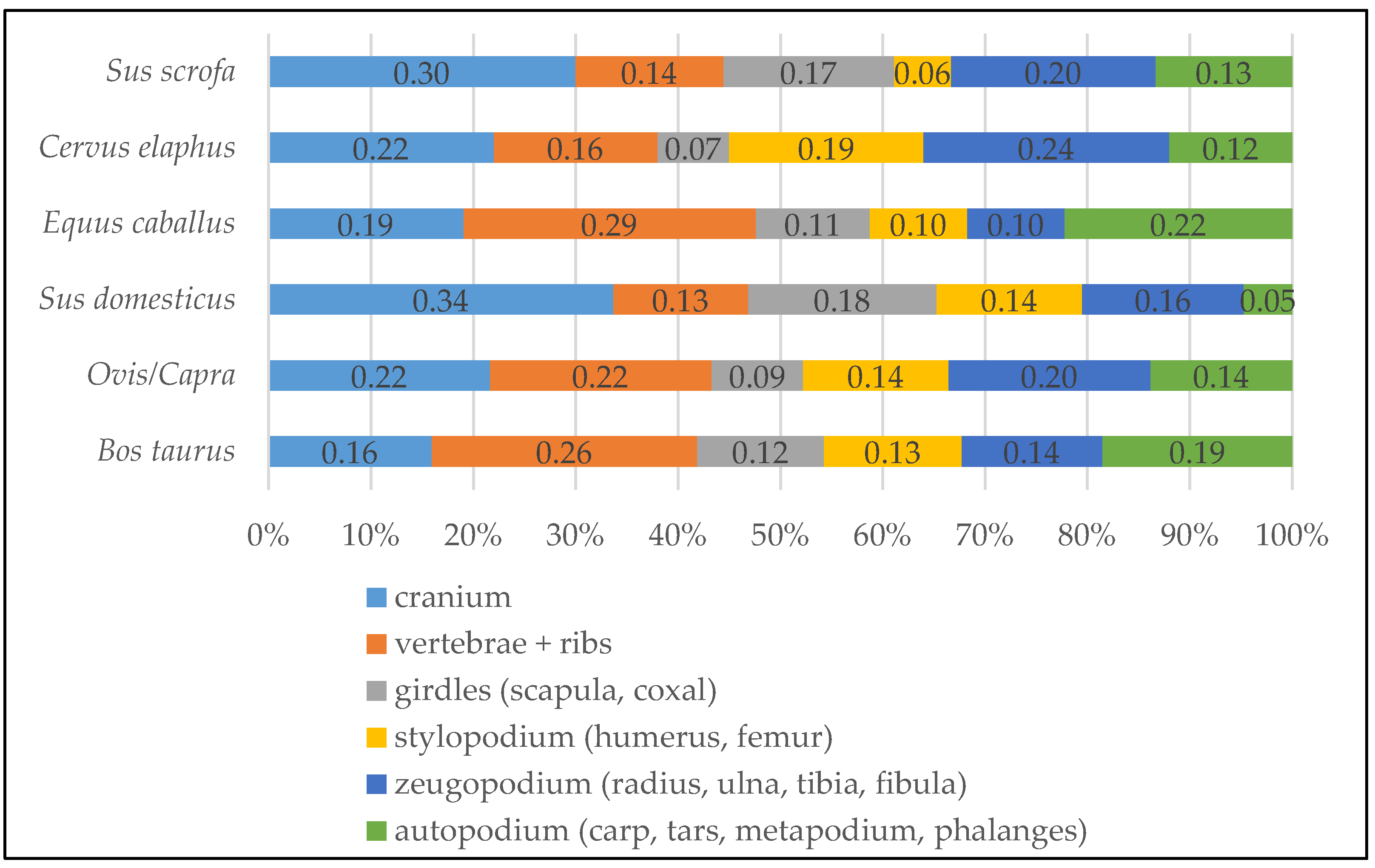
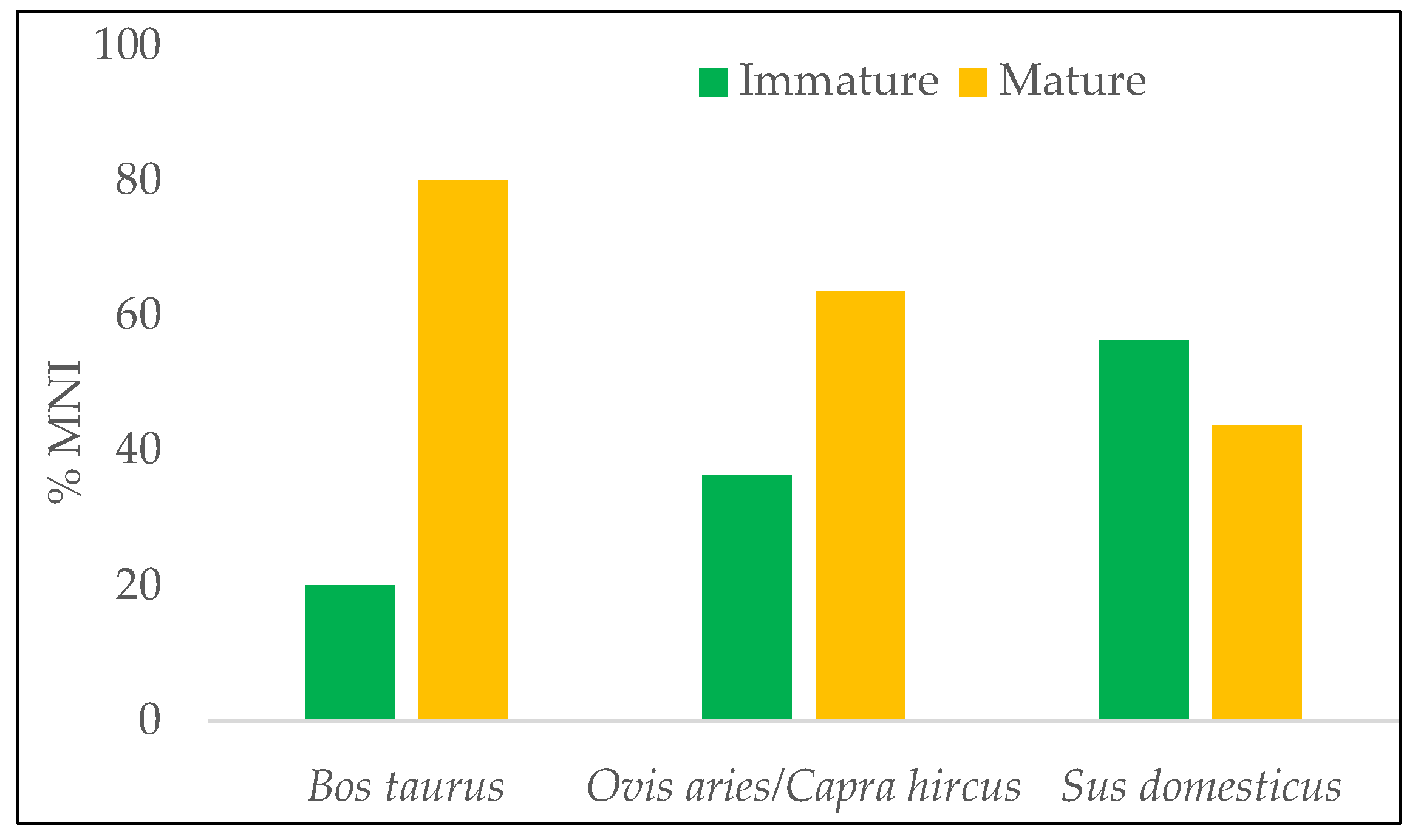
4.2. Archaeozoology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teodor, E.S. The Border area between Moesia Secunda and Scythia Minor in a topographical approach. In Identităţi Culturale, Locale Şi Regionale în Context European. Studii de Arheologie şi Antropologie Istorică. In Memoriam Alexandri V. Matei; Matei, A.V., Pop, H., Bejinariu, I., Băcueţ-Crişan, S., Băcueţ-Crişan, C., Eds.; Mega: Cluj-Napoca, Romania, 2010; pp. 421–438. [Google Scholar]
- Zahariade, M. Moesia Secunda, Scythia și Notitia Dignitatum; Biblioteca de arheologie, Editura Academiei: București, Romania, 1988. [Google Scholar]
- Zahariade, M. The Scythian Section of Notitia Dignitatum: A Structural and Chronological Analysis. Ad Fines Imperii Romani. Studia Thaddaeo Sarnowski Septuagenario ab Amicis, Collegis Discipulisque Dedicata; Institute of Archaeology, University of Warsaw: Warsaw, Poland, 2015; pp. 151–172. [Google Scholar]
- Țentea, O.; Opriș, I.C.; Popescu, F.-M.; Rațiu, A.; Băjenaru, C.; Călina, V. Frontiera romană din Dobrogea. O trecere în revistă și o actualizare. Cercet. Arheol. 2019, 26, 9–82. [Google Scholar] [CrossRef]
- Zahariade, M.; Lungu, V.; Covacef, Z. Scythia Minor. A History of a Later Roman Province (284-681); Pontic Provinces of the Later Roman Empire I; Adolf M. Hakkert: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Ellis, L.; Places, E. A Chorological Approach to Identity and Territory in Scythia Minor (Second-Seventh Centuries). In Romans, Barbarians, and the Transformation of the Roman World; Ralph, W.M., Danuta, S., Eds.; Ashgate: Surrey, UK, 2011; pp. 241–251. [Google Scholar]
- Ştefan Al., S. Cetatea romană târzie de la Murighiol. Studiu aerofotografic. Peuce 1984, 9, 663–674. [Google Scholar]
- Dintchev, V. Sur la caracteristique d‘Iatrus (deuxieme moitie du Ive—debut du Ve s.) in Vorträge der Internationalen Konferenz Svištov, Bulgarien (1–5 September 1998). In Der Limes an der Unteren Donau von Diokletian bis Heraklios; Herausgegeben von Gerda von Bülow und Alexandra Milčeva: Sofia, Bulgaria, 1999; pp. 165–174. [Google Scholar]
- Van Lanen, R.; de Kleijn, M.; Gouw-Bouman, M.; Pierik, H. Exploring Roman and early-medieval habitation of the Rhine–Meuse delta: Modelling large-scale demographic changes and corresponding land-use impact. Neth. J. Geosci. 2018, 97, 45–68. [Google Scholar] [CrossRef] [Green Version]
- King, A. The Romanization of diet in the western empire: Comparative archaeozoological studies. In Italy and the West, Comparative Issues in Romanization; Keay, S., Terrenato, N., Eds.; Oxbow Books: Oxford, UK, 2001; pp. 210–223. [Google Scholar]
- El Susi, G. Data about hunting practices by Halmiris (Murighiol, Tulcea County) inhabitants in 4th–7th centuries AD. Cult. și civ. la Dunărea de Jos 2008, 24, 201–210. [Google Scholar]
- El Susi, G. Animal breedind in late Roman settlements from Dobrudja, in the light of research at Murighiol (Halmyris, Tulcea county). Cult. și civ. la Dunărea de Jos 2011, 28, 170–189. [Google Scholar]
- Stanc, S.; Danu, M.; Paraschiv, D.; Bejenaru, L. Bioarcheological Indicators Related to Human–Environmental Interactions in a Roman–Byzantine Settlement in Southeast Romania: Ibida Fortress. SAGE Open 2020, 10, 2158244020969664. [Google Scholar] [CrossRef]
- Kibblewhite, M.; Tóth, G.; Hermann, T. Predicting the preservation of cultural artefacts and buried material in soil. Sci. Total Environ. 2015, 529, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Dal Corso, M.; Out, W.A.; Ohlrau, R.; Hofmann, R.; Dreibrodt, S.; Videiko, M.; Müller, J.; Kirleis, W. Where are the cereals? Contribution of phytolith analysis to the study of subsistence economy at the Trypillia site Maidanetske (ca. 3900-3650 BCE), central Ukraine. J. Arid Environ. 2018, 157, 137–148. [Google Scholar] [CrossRef]
- Danu, M.; Diaconu, V.; Bejenaru, L. Chalcolithic agropastoralism traces in the site of Răucești (Neamț county, Romania): Phytoliths and animal remains. Int. J. Conserv. Sci. 2016, 7, 1071–1080. [Google Scholar]
- Danu, M.; Messager, E.; Carozza, J.M.; Carozza, L.; Bouby, L.; Philibert, S.; Anderson, P.; Burens, A.; Micu, C. Phytolith evidence of cereal processing in the Danube Delta during the Chalcolithic period. Quat. Int. 2019, 504, 128–138. [Google Scholar] [CrossRef]
- Delhon, C.; Binder, D.; Verdin, P.; Mazuy, A. Phytoliths as a seasonality indicator? The example of the Neolithic site of Pendimoun, south-eastern France. Veget. Hist. Archaeobotany 2020, 29, 229–240. [Google Scholar]
- Malaxa, D.I.; Stanc, M.S.; Bărbat, I.A.; Gâza, O.; Păceşilă, D.; Bejenaru, L.; Danu, M. Farming Beginning in Southwestern Transylvania (Romania). Subsistence Strategies in Mureş Valley during the Early Neolithic. Diversity 2022, 14, 894. [Google Scholar] [CrossRef]
- Pető, Á.; Kenéz, Á.; Prunner, A.C.; Lisztes-Szabó, Z. Activity area analysis of a Roman period semi-subterranean building by means of integrated archaeobotanical and geoarchaeological data. Veg. Hist. Archaeobotany 2015, 24, 101–120. [Google Scholar] [CrossRef]
- Wouters, B.; Devos, Y.; Vrydaghs, L.; Ball, T.; De Winter, N.; Reygel, P. An integrated micromorphological and phytolith study of urban soils and sediments from the Gallo-Roman town Atuatuca Tungrorum, Belgium. Geoarchaeology 2019, 34, 448–466. [Google Scholar] [CrossRef]
- Druzhinina, O.; van den Berghe, K.; Golyeva, A. Application of the microbiomorphic (phytolith) analysis in the geoarchaeological study of the land-use at the Voorthuizen-Wikselaarseweg archaeological site (Netherlands). In Proceedings of the EGU General Assembly 2021, online, 19–30 April 2021. EGU21-9043. [Google Scholar] [CrossRef]
- Kirchner, A.; Herrmann, N.; Matras, P.; Müller, I.; Meister, J.; Schattner, T.G. A pedo-geomorphological view on land use and its potential in the surroundings of the ancient Hispano-Roman city Munigua (Seville, SW Spain). EG Quat. Sci. J. 2022, 71, 123–143. [Google Scholar] [CrossRef]
- Pîrnău, R.-G.; Stanc, S.M.; Roșca, B.; Bejenaru, L.; Danu, M. A Multidisciplinary Approach to Human–Environmental Interactions at the Roman-Byzantine Ibida Fortress (Dobrogea, South-Eastern Romania). Environ. Archaeol. 2022. [Google Scholar] [CrossRef]
- Pîrnău, R.G.; Patriche, C.-V.; Roşca, B.; Vasiliniuc, I.; Vornicu, N.; Stanc, S. Soil spatial patterns analysis at the ancient city of Ibida (Dobrogea, SE Romania), via portable X-ray fluorescence spectrometry and multivariate statistical methods. Catena 2020, 189, 104506. [Google Scholar] [CrossRef]
- Stanc, S.; Stănică, A.D.; Bejenaru, L.; Danu, M. Archaeological animal remains from Noviodunum fortress. Int. J. Conserv. Sci. 2021, 12, 195–204. [Google Scholar]
- Stanc, S. Arheozoologia Primului Mileniu d.Hr. Pentru Teritoriul Cuprins între Dunăre şi Marea Neagră; Editura Universităţii Alexandru Ioan Cuza Iaşi: Iaşi, Romania, 2009. [Google Scholar]
- Stanc, S.; Bejenaru, L. Animal resources exploited in settlements of the 2nd–7th centuries in the area between Danube and Black Sea: Archaeozoological data. Istros 2013, 19, 389–409. [Google Scholar]
- Neef, R. Archäobotanische Untersuchungen im spätantiken Iatrus/Krivina (Grabungskampagnen 1992–2000), in Gerda von Büllow, Burkhard Böttger, Sven Conrad, Bernhard Döhle, Gudrun Gomolka-Fuchs, Edith Schönert-Geiss, Dimităr Stančev und Klaus Wachtel. In Iatrus-Krivina. Spätantike Befestigung und frühmittelalterliche Siedlung an der Unteren Donau. Band VI: Ergebnisse der Ausgrabungen 1992–2000; Römisch-Germanische Kommission des Deutschen Archaölogischen Instituts, Frankfurt a.M., Verlag Philipp von Zabern, Mainz am Rhein: Frankfurt, Germany, 2007; pp. 415–446. [Google Scholar]
- Lazarova, M. Pollenanalytische Untersuchungen an Material aus den archäologischen Ausgrabungen im Kastell Iatrus, in Gerda von Büllow, Burkhard Böttger, Sven Conrad, Bernhard Döhle, Gudrun Gomolka-Fuchs, Edith Schönert-Geiss, Dimităr Stančev und Klaus Wachtel. In Iatrus-Krivina. Spätantike Befestigung und Frühmittelalterliche Siedlung an der Unteren Donau. Band VI: Ergebnisse der Ausgrabungen 1992–2000; Römisch-Germanische Kommission des Deutschen Archaölogischen Instituts, Frankfurt a.M., Verlag Philipp von Zabern, Mainz am Rhein: Frankfurt, Germany, 2007; pp. 447–456. [Google Scholar]
- Niemczak, K. Archaeozoological analysis of animal remains from the Roman fortress camp in Novae. Novensia 2016, 27, 55–82. [Google Scholar]
- Vuković-Bogdanović, S. Roman archaeozoology in Serbia: State of the discipline and preliminary results. In Archaeology and Science; Institute of Archaeology: Belgrade, Serbia, 2016; Volume 13, pp. 99–113. [Google Scholar]
- King, A. Diet in the Roman world: A regional inter-site comparison of the mammal bones. J. Rom. Archaeol. 1999, 12, 168–202. [Google Scholar] [CrossRef]
- Zahariade, M. The Aegyssus/Ad Stoma-Roman Frontier Sector in Extrema Scythiae Minoris: Understanding a Defensive System in a River Delta Environment. In Kontaktzone Balkan. Contributions to the International Colloquium “The Danube-Balkan Region as a Contact Zone between East-West and North-South” from 16–18 May 2012 in Frankfurt a. M.; von Bülow, G., Ed.; Rdolf Habelt Verlag: Bonn, Germany, 2015; pp. 219–235. [Google Scholar]
- Suceveanu, A.; Zahariade, M. Du nom antique de la cité romaine tardive d’lndependenta (dep. de Tulcea). Dacia N.S. 1987, 31, 87–96. [Google Scholar]
- Zahariade, M.; Suceveanu, A.; Opaiț, A.; Opaiț, C.; Topoleanu, F. Early and Late Roman Fortification at lndependența, Tulcea County. Dacia N.S. 1987, 31, 97–106. [Google Scholar]
- Zahariade, M. An Early and Late Roman Fort on the Lower Danube Limes: Halmyris (Independența, Tulcea County, Romania). In Roman Frontier Studies 1989, Proceedings of the XVth International Congress of Roman Frontier Studies, University of Canterbury, UK, 2-10 September 1989; Maxfield, V.A., Dobson, M.J., Eds.; University of Exeter Press: Exeter, UK, 1991; pp. 311–317. [Google Scholar]
- Suceveanu, A.; Zahariade, M.; Topoleanu, F.; Poenaru Bordea, G. Halmyris. In Halmyris I. Monografie Arheologică; Editura Neremia Napocae: Cluj-Napoca, Romania, 2003. [Google Scholar]
- Zahariade, M. From dava to civitas: Halmyris, a Getic, Early and Late Roman Settlement on the Lower Danube. In The Bosporus: Gateway between the Ancient West and East (1st Millennium BC-5th Century AD), Proceedings of the Fourth International Congress on Black Sea Antiquities Istanbul, 14th–18th September 2009; Tsetskhladze, G., Atasoy, S., Avram, A., Dönmez, S., Hargrave, J., Eds.; BAR International Series 2517: Oxford, UK, 2013; pp. 207–212. [Google Scholar]
- Romanescu, G.; Mihu-Pintilie, A.; Carboni, D. The City-Port of Halmyris: An Integrated Geoarchaeological and Environmental Approach to the Last Roman Bastion on the Eastern Flank of the Danubian Limes; PESD, Alexandru Ioan Cuza University Publishing House: Iași, Romania, 2018; Volume 2, pp. 25–46. [Google Scholar]
- Giaime, M.; Magne, G.; Bivolaru, A.; Grandouin, E.; Marriner, N.; Morhange, C. Halmyris: Geoarchaeology of a fluvial harbour on the Danube Delta (Dobrogea, Romania). Holocene 2019, 29, 313–327. [Google Scholar] [CrossRef] [Green Version]
- Suceveanu, A.; Zahariade, M. A New Vicus on the Territory of Roman Dobrudja. Dacia N.S. 1986, 30, 109–120. [Google Scholar]
- Zahariade, M. The Northern and Western Gates of the Halmyris Fort. In Limes XX. XXth International Congress of Roman Frontier Studies (LIMES XX Gladius, Anejos 13); Morillo, A., Hanel, N., Martin, E., Eds.; Consejo Superior de Investigaciones Cientificas: Madrid, Spain, 2009; pp. 77–88. [Google Scholar]
- Mărgineanu Cârstoiu, A.; Apostol, V. La fortification d’Halmyris: Étude architecturale des Portes Ouest et Nord. Caiete ARA 2015, 6, 37–78. [Google Scholar] [CrossRef]
- Zahariade, M. The Halmyris Episcopal Basilica and the Martyrs’ Crypt. Il Mar Nero 2001–2003, 5, 143–168. [Google Scholar]
- Zahariade, M. The Episcopal Basilica from Halmyris and the Crypt of Epictetus and Astion. Thraco-Dacica S.N. 2009, 1, 131–150. [Google Scholar]
- Mirițoiu, N.; Soficaru, A.D. Anthropological study of the human bones found in the crypt from Halmyris. Thraco-Dacica S.N. 2009, 1, 151–181. [Google Scholar]
- Nuțu, G. Le “Zwiebelknopffibeln” da Halmyris (Provincia Scythia). Quad. Friulani Di Archeol. 2010, 20, 99–108. [Google Scholar]
- Nuțu, G. Belt buckles, strap-ends and appliqués from Halmyris (Moesia Inferior/Scythia). Novensia 2011, 22, 171–199. [Google Scholar]
- Rafailă-Stan, S.; Nuţu, G. Worked bone and antler from Halmyris: An insight on everyday life of a frontier post of scythia. Quat. Int. 2018, 472, 142–148. [Google Scholar] [CrossRef]
- Zahariade, M. Personal Names at Halmyris. Thraco-Dacica S.N. 2012–2013, 4–5, 159–182. [Google Scholar]
- Zahariade, M.; Alexandrescu, C.G. Greek and Latin Inscriptions from Halmyris. Inscriptions on Stone, Signa, and Instruments Found between 1981 and 2010; BAR IS: Oxford, UK, 2011. [Google Scholar]
- Lentfer, C.J.; Boyd, W.E. A comparison of three methods for the extraction of phytoliths from sediments. J. Archaeol. Sci. 1998, 25, 1159–1183. [Google Scholar] [CrossRef]
- International Committee on Phytolith Taxonomy (ICPT). International code for phytolith nomenclature (ICPN) 2.0. Ann. Bot. 2019, 124, 189–199. [CrossRef]
- Udrescu, M.; Bejenaru, L.; Hriscu, C. Introducere în Arheozoologie; Editura Corson: Iași, Romania, 1999. [Google Scholar]
- Reitz, E.J.; Wing, E.S. Zooarchaeology (Cambridge Manuals in Archaeology), 2nd ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- von den Driesch, A. A guide to the measurement of animal bones from archaeological sites. In Peabody Museum Bulletin 1; Harvard University Press: Cambridge, MA, USA, 1976; pp. 1–137. [Google Scholar]
- Fock, J. Metrische Untersuchungen an Metapodien Einiger Europäischer Rinderrassen. Diss.Med. vet., LMU-München; Ludwig-Maximilians-Universität München: München, Germany, 1966. [Google Scholar]
- Teichert, M. Osteometrische Untersuchungen zur Berechnung der Widerristhohe bei Schafen. In Archaeozoological Studies; Clason, A.T., Ed.; North-Holland Publishing Company: Amsterdam, The Netherlands, 1975; pp. 51–69. [Google Scholar]
- Schramm, Z. Różnice morfologiczne niektórych kości kozy i owcy/Morphological differences of some goat and sheep bones. Roczniki Wyższej Szkoły Rolniczej w Poznaniu. Wydział Zootech. 1967, 10, 107–133. [Google Scholar]
- Teichert, M. Osteometrische untersuchungen zur berechnung der widerristhöhe bei vor- und frühgeschichtlichen schweinen. Kühn-Archiv 1969, 83, 237–292. [Google Scholar]
- Teichert, M. Withers height calculations for pigs—Remarks and experience. In Proceedings of the Handout distributed at the 6th ICAZ Conference, Washington, DC, USA, 21–25 May 1990. [Google Scholar]
- Harcourt, R.A. The dog in prehistoric and early historic Britain. J. Archaeol. Sci. 1974, 1, 151–175. [Google Scholar] [CrossRef]
- Piperno, D.R.; Pearsall, D.M. The Silica Bodies of Tropical American Grasses: Morphology, Taxonomy, and Implications for Grass Systematics and Fossil Phytolith Identification. Smithson. Contrib. Bot. 1998, 85, 1–22. [Google Scholar] [CrossRef]
- Ball, T.B.; Gardner, J.S.; Anderson, N. An approach to identifying inflorescence phytoliths from selected species of wheat and barley. In Phytoliths: Applications in Earth Sciences and Human History; Meunier, J.D., Colin, F., Eds.; A. A. Balkema: Abingdon, UK, 2001; pp. 289–302. [Google Scholar]
- Novello, A.; Barboni, D. Grass inflorescence phytoliths of useful species and wild cereals from sub-Saharan Africa. J. Archaeol. Sci. 2015, 59, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Honaine, M.; Zucol, A.; Osterrieth, M. Phytolith analysis of Cyperaceae from the Pampean region, Argentina. Aust. J. Bot. 2009, 57, 512–523. [Google Scholar] [CrossRef]
- Strömberg, C.A.E. The Origin and Spread of Grass-Dominated Ecosystems during the Tertiary of North America and How It Relates to the Evolution of Hypsodonty in Equids. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2003. [Google Scholar]
- Novello, A.; Barboni, D.; Berti-Equille, L.; Mazur, J.-C.; Poilecot, P.; Vignaud, P. Phytolith signal of aquatic plants and soils in Chad, Central Africa. Rev. Palaeobot. Palynol. 2012, 178, 43–58. [Google Scholar] [CrossRef]
- Esau, K. Plant Anatomy, 2nd ed.; Wiley: New York, USA, 1965. [Google Scholar]
- Albert, R.M.; Lavi, O.; Estroff, L.; Weiner, S.; Tsatskin, A.; Ronen, A.; Lev-Yadun, S. Mode of occupation of Tabun Cave, Mt Carmel, Israel during the Mousterian period: A study of the sediments and phytoliths. J. Archaeol. Sci. 1999, 26, 1249–1260. [Google Scholar] [CrossRef]
- Delhon, C.; Alexandre, A.; Berger, J.F.; Thiébault, S.; Brochier, J.L.; Meunier, J.D. Phytolith assemblages as a promising tool for reconstructing Mediterranean Holocene vegetation. Quat. Res. 2003, 59, 48–60. [Google Scholar] [CrossRef]
- Runge, F. The opal phytolith inventory of soils in Central Africa. Quantities, shapes, classification and spectra. Rev. Palaeobot. Palynol. 1999, 107, 23–53. [Google Scholar] [CrossRef]
- Danu, M.; Carozza, J.-M.; Messager, E. Les sédiments comme révélateurs des activités anthropiques dans le delta du Danube—phytolithes. In Au-Delà de la Nature : Le Bas Danube et Son Delta Durant les Huit Derniers Millénaires; Carozza, L., Micu, C., Eds.; Editura Mega: Cluj, România, 2022. [Google Scholar]
- Bolomey, A. Materiale paleofaunistice de la Histria. Stud. Si Cercet. Antropol. 1965, 2, 179–189. [Google Scholar]
- Haimovici, S. Ameliorarea rasiala a animalelor domestice evaluata prin specia taurine (Bos taurus), facuta de catre romani, dupa venirea lor in Antichitate, in actuala Dobroge. Pontica 2006, 39, 349–353. [Google Scholar]
- Duval, C.; Lepetz, S.; Horard-Herbin, M.-P. Diversité des cheptels et diversification des morphotypes bovins dans le tiers nord-ouest des Gaules entre la fin de l’âge du Fer et la période romaine; Gallia—Archéologie de la France Antique, CNRS Éditions: Paris, France, 2012; Volume 69, pp. 79–114. [Google Scholar]
- Audoin-Rouzeau, F. La taille du boeuf domestique en Europe de l‘antiquite aux tepms modernes. In Fiches D’osteologie Pour L’archeologie, Serie B: Mammiferes, 2, APDCA; Juan-les-Pines: Paris, France, 1991; pp. 3–40. [Google Scholar]
- Audoin-Rouzeau, F. Compter et mesurer les os animaux. Pour une histoire de l’élevage et de l’alimentation en Europe de l’Antiquité aux Temps Modernes. Hist. Mes. 1995, 10, 277–312. [Google Scholar] [CrossRef]
- Haimovici, S. Studiul arheozoologic al unor resturi faunistice descoperite în nivelul aparţinând sec. al VI-lea p.Chr. al cetăţii Histria. Pontica 2007, XL, 541–558. [Google Scholar]
- Haimovici, S. Studiul arheozoologic al resturilor de la Dinogetia (Garvăn), aparţinând epocii romane târzii. Peuce 1991, 10, 355–360. [Google Scholar]
- Stanc, S.; Radu, V.; Bejenaru, L. Analyse archeozoologique des restes de poissons provenant du site de Slava Rusă (Roumanie). In Archéologie du Poisson. 30 ans d‘Archeo-Ichtyologie au CNRS. XXVIIIe Rencontres Internationales D’archéologie et D’histoire D’antibes; Bearez, P., Grouard, S., Clavel, B., Eds.; Éditions APDCA: Antibes, France, 2008; pp. 257–279. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Rondel | Bulliform flabellate | Spheroid | Acute bulbosus | Elongate entire | Elongate dendritic | Crenate | Polylobate | Bilobate | Cross | Saddle | Blocky | Cyperaceae type | Tracheary | Diatoms | Sponge Spicules | Silica Skeleton | Phytolith Sum |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MUR 12 | 223 | 12 | 29 | 7 | 29 | 22 | 3 | 1 | 0 | 1 | 0 | 2 | 2 | 0 | 3 | 0 | 80 | 331 |
| MUR 11 | 218 | 1 | 19 | 11 | 36 | 20 | 4 | 1 | 3 | 4 | 0 | 1 | 0 | 0 | 0 | 0 | 47 | 318 |
| MUR 10 | 248 | 8 | 11 | 6 | 27 | 23 | 4 | 0 | 1 | 0 | 0 | 4 | 1 | 0 | 0 | 0 | 38 | 333 |
| MUR 9 | 248 | 2 | 10 | 10 | 50 | 51 | 8 | 2 | 1 | 0 | 2 | 1 | 1 | 0 | 0 | 0 | 12 | 386 |
| MUR 8 | 295 | 5 | 8 | 11 | 29 | 45 | 3 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 1 | 19 | 399 |
| MUR 7 | 281 | 4 | 11 | 7 | 12 | 46 | 7 | 1 | 1 | 1 | 2 | 4 | 0 | 0 | 2 | 0 | 78 | 377 |
| MUR 6 | 207 | 4 | 24 | 9 | 35 | 20 | 5 | 5 | 3 | 1 | 0 | 3 | 1 | 1 | 1 | 2 | 74 | 318 |
| MUR 5 | 225 | 1 | 21 | 6 | 14 | 23 | 7 | 0 | 1 | 1 | 1 | 2 | 0 | 1 | 0 | 0 | 109 | 303 |
| MUR 4 | 242 | 4 | 13 | 15 | 36 | 58 | 5 | 0 | 2 | 0 | 1 | 8 | 3 | 1 | 0 | 2 | 115 | 388 |
| MUR 3 | 264 | 2 | 22 | 4 | 28 | 66 | 5 | 0 | 0 | 1 | 1 | 7 | 0 | 0 | 0 | 0 | 46 | 400 |
| MUR 2 | 262 | 5 | 13 | 13 | 41 | 77 | 7 | 0 | 2 | 2 | 2 | 1 | 1 | 1 | 0 | 0 | 38 | 427 |
| MUR 1 | 325 | 6 | 25 | 8 | 36 | 90 | 7 | 0 | 4 | 1 | 2 | 3 | 0 | 1 | 0 | 1 | 34 | 508 |
| Taphonomy Evidence | NR | % |
|---|---|---|
| Remains with butchering traces | 574 | 26.00 |
| Remains with burn traces | 74 | 3.35 |
| Remains with animal teeth marks | 187 | 8.47 |
| Manufactured bones and antlers | 12 | 0.54 |
| Total sample | 2207 | 100 |
| Taxonomic Group | Species | Extramuros Sample | Intramuros Sample [11,12] | ||
|---|---|---|---|---|---|
| NR | % | NR | % | ||
| Molluscs | Unio sp. (freshwater mussel) | 11 | 0.50 | - | - |
| Mytilus sp. (mussel) | 1 | 0.04 | - | - | |
| Unidentified molluscs | - | - | 9 | 0.25 | |
| Total molluscs | 12 | 0.54 | 9 | 0.25 | |
| Fish | Cyprinus carpio (common carp) | 43 | 1.95 | - | - |
| Silurus glanis (wels catfish) | 77 | 3.49 | - | - | |
| Esox lucius (northern pike) | 4 | 0.18 | - | - | |
| Acipenser sp. (sturgeons) | 8 | 0.36 | - | - | |
| Unidentified Teleostei | 37 | 1.68 | - | - | |
| Total fish | 169 | 7.66 | * | - | |
| Reptiles | Testudo sp. (tortoise) | 3 | 0.14 | 0 | - |
| Birds | Gallus domesticus (chicken) | 4 | 0.18 | 50 | 1.41 |
| Anser domesticus (goose) | - | - | 5 | 0.14 | |
| Anas plathyrinchos (duck) | - | - | 4 | 0.11 | |
| Unidentified bird | 5 | 0.23 | 28 | 0.79 | |
| Total birds | 9 | 0.41 | 87 | 2.45 | |
| Mammals | Total mammals | 2014 | 91.26 | 3457 | 97.30 |
| Total sample | 2207 | 100 | 3553 | 100 | |
| Species | Extramuros Sample | Intramuros Sample [11,12] | ||||
|---|---|---|---|---|---|---|
| NR | % | NR | % | NR | % | |
| Bos taurus (cattle) | 929 | 54.11 | 25 | 30.49 | 685 | 24.04 |
| Ovis aries/Capra hircus (sheep/goat) | 268 | 15.61 | 11 | 13.41 | 642 | 22.53 |
| Sus domesticus (pig) | 190 | 11.07 | 16 | 19.51 | 708 | 24.85 |
| Equus caballus (horse) | 63 | 3.67 | 4 | 4.88 | 127 | 4.46 |
| Canis familiaris (dog) | 46 | 2.68 | 4 | 4.88 | 58 | 2.04 |
| Equus asinus (donkey) | 10 | 0.58 | 2 | 2.44 | 16 | 0.56 |
| Felis domesticus (cat) | - | - | - | - | 8 | 0.28 |
| Total domestic mammals | 1506 | 87.71 | 62 | 75.61 | 2244 | 78.76 |
| Cervus elaphus (red deer) | 100 | 5.82 | 6 | 7.32 | 209 | 7.33 |
| Sus scrofa (wild boar) | 84 | 4.89 | 5 | 6.10 | 330 | 11.58 |
| Capreolus capreolus (roe deer) | 9 | 0.52 | 2 | 2.44 | 16 | 0.56 |
| Lepus europaeus (hare) | 2 | 0.12 | 1 | 1.22 | 10 | 0.35 |
| Canis lupus (wolf) | 4 | 0.23 | 1 | 1.22 | - | - |
| Bos primigenius (auroch) | 1 | 0.06 | 1 | 1.22 | 1 | 0.04 |
| Castor fiber (beaver) | 2 | 0.12 | 1 | 1.22 | 5 | 0.18 |
| Meles meles (badger) | 8 | 0.47 | 2 | 2.44 | 5 | 0.18 |
| Lutra lutra (Eurasian otter) | - | - | - | - | 10 | 0.35 |
| Martes martes (pine marten) | - | - | - | - | 8 | 0.28 |
| Vulpes vulpes (fox) | 1 | 0.06 | 1 | 1.22 | 11 | 0.39 |
| Total wild mammals | 211 | 12.29 | 20 | 24.39 | 605 | 21.24 |
| Total identified mammals | 1717 | 100.00 | 82 | 100.00 | 2849 | 100 |
| Unidentified mammals | 297 | 608 | ||||
| Total mammals | 2014 | 3457 | ||||
| Species | Category Immature/Mature | MNI | Age |
|---|---|---|---|
| Bos taurus (cattle) | Immature (under 2.5 years old) | 1 | 4–6 months |
| 2 | 6–12 months | ||
| 1 | 12–18 months | ||
| 1 | 2–2.5 years | ||
| Total | 5 | ||
| Mature MNI (over 2.5 years old) | 1 | 2.5 years | |
| 6 | 2.5–4 years | ||
| 13 | Over 4 years | ||
| Total | 20 | ||
| Ovis aries/Capra hircus (sheep/goat) | Immature (under 2 years old) | 1 | 3–6 months |
| 1 | 1–1.5 years | ||
| 2 | 1.5–2 years | ||
| Total | 4 | ||
| Mature (over 2 years old) | 1 | 2 years | |
| 2 | 2–3 years | ||
| 4 | Over 3 years | ||
| Total | 7 | ||
| Sus domesticus (pig) | Immature (under 2 years old) | 3 | 4–6 months |
| 3 | 6–9 months | ||
| 3 | 13–18 months | ||
| Total | 9 | ||
| Mature MNI (over 2 years old) | 1 | 2 years | |
| 3 | 2–3 years | ||
| 3 | Over 3 years | ||
| Total | 7 |
| Species | Anatomical Element | Bone Length (mm) | Sex Estimation | Withers Height (cm) |
|---|---|---|---|---|
| Bos taurus | metacarpus | 218 | castrated | 133.41 |
| Bos taurus | metacarpus | 205 | castrated | 125.46 |
| Bos taurus | metacarpus | 204 | castrated | 124.84 |
| Bos taurus | metacarpus | 190 | castrated | 116.28 |
| Bos taurus | metacarpus | 192 | female | 115.20 |
| Bos taurus | metacarpus | 205 | female | 123.00 |
| Bos taurus | metacarpus | 186 | female | 111.60 |
| Bos taurus | metacarpus | 204 | female | 122.40 |
| Bos taurus | metacarpus | 201 | female | 120.60 |
| Bos taurus | metacarpus | 195 | female | 117.00 |
| Bos taurus | metacarpus | 195 | male | 121.87 |
| Bos taurus | metacarpus | 199 | castrated | 121.78 |
| Bos taurus | metacarpus | 202 | castrated | 123.62 |
| Bos taurus | metacarpus | 203 | castrated | 124.23 |
| Bos taurus | metacarpus | 183.5 | castrated | 112.30 |
| Bos taurus | metacarpus | 192 | castrated | 117.50 |
| Bos taurus | metacarpus | 181 | female | 108.60 |
| Bos taurus | metacarpus | 182 | female | 109.20 |
| Bos taurus | metacarpus | 215 | male | 134.37 |
| Bos taurus | metatarsus | 217 | castrated | 118.26 |
| Bos taurus | metatarsus | 248 | castrated | 135.16 |
| Bos taurus | metatarsus | 234 | castrated | 127.53 |
| Bos taurus | metatarsus | 244 | castrated | 132.98 |
| Bos taurus | metatarsus | 225 | female | 120.37 |
| Bos taurus | metatarsus | 218 | female | 116.63 |
| Bos taurus | metatarsus | 220 | female | 117.70 |
| Bos taurus | metatarsus | 232 | male | 128.76 |
| Bos taurus | metatarsus | 208 | male | 115.44 |
| Bos taurus | metatarsus | 227 | female | 121.44 |
| Bos taurus | metatarsus | 218 | female | 116.63 |
| Bos taurus | metatarsus | 230 | castrated | 125.35 |
| Bos taurus | metatarsus | 237 | castrated | 129.16 |
| Capra hircus | metacarpus | 137 | - | 78.77 |
| Capra hircus | metacarpus | 126.5 | - | 72.73 |
| Capra hircus | metatarsus | 126 | - | 67.28 |
| Ovis aries | metatarsus | 148.5 | - | 67.42 |
| Ovis aries | metatarsus | 141 | - | 64.01 |
| Ovis aries | metatarsus | 150 | - | 68.10 |
| Ovis aries | metatarsus | 147 | - | 66.73 |
| Sus domesticus | metacarpus 4 | 77.5 | - | 78.66 |
| Sus scrofa | calcaneus | 106 | - | 101.60 |
| Sus scrofa | metatarsus 3 | 108 | - | 101.43 |
| Sus scrofa | metatarsus 3 | 108.5 | - | 101.89 |
| Sus scrofa | radius | 210 | - | 109.16 |
| Sus scrofa | astragalus | 53 | - | 97.17 |
| Sus scrofa | astragalus | 56 | - | 102.5 |
| Canis familiaris | tibia | 193 | - | 57.29 |
| Canis familiaris | femur | 141 | - | 42.97 |
| Canis familiaris | humerus | 170 | - | 55.65 |
| Canis familiaris | femur | 217 | - | 66.84 |
| Canis familiaris | tibia | 196 | - | 58.17 |
| Canis familiaris | femur | 195.5 | - | 60.09 |
| Sample | Taxonomic Group | Mollusca (Molluscs) | Pisces (Fish) | Reptilia (Reptiles) | Aves (Birds) | Mammalia (Mammals) | Total |
|---|---|---|---|---|---|---|---|
| Halmyris (extramuros) | NR | 12 | 169 | 3 | 9 | 2013 | 2207 |
| % | 0.54 | 7.66 | 0.14 | 0.41 | 91.26 | 100 | |
| Halmyris (intramuros) [11] | NR | 9 | 0 | 0 | 87 | 3457 | 3553 |
| % | 0.25 | 0 | 0 | 2.45 | 97.3 | 100 | |
| Histria (6th century AD) [80] | NR | 0 | 0 | 0 | 0 | 570 | 570 |
| % | 0 | 0 | 0 | 0 | 100 | 100 | |
| Ibida [13] | NR | 0 | 33 | 0 | 0 | 923 | 956 |
| % | 0 | 3.45 | 0 | 0 | 96.55 | 100 | |
| Ibida [24] | NR | 31 | 9131 | 0 | 186 | 8045 | 17393 |
| % | 0.18 | 52.50 | 0 | 1.07 | 46.25 | 100 | |
| Dinogetia (4th–6th centuries AD) [81] | NR | 16 | 28 | 0 | 7 | 129 | 180 |
| % | 8.89 | 15.55 | 0 | 3.89 | 71.67 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanc, M.S.; Bejenaru, L.; Nuțu, G.; Mototolea, A.C.; Danu, M. Palaeoeconomy and Palaeoenvironment of Halmyris—A Roman Settlement in Southeast Romania: Archaeozoological and Phytolith Evidences. Diversity 2023, 15, 373. https://doi.org/10.3390/d15030373
Stanc MS, Bejenaru L, Nuțu G, Mototolea AC, Danu M. Palaeoeconomy and Palaeoenvironment of Halmyris—A Roman Settlement in Southeast Romania: Archaeozoological and Phytolith Evidences. Diversity. 2023; 15(3):373. https://doi.org/10.3390/d15030373
Chicago/Turabian StyleStanc, Margareta Simina, Luminița Bejenaru, George Nuțu, Aurel Constantin Mototolea, and Mihaela Danu. 2023. "Palaeoeconomy and Palaeoenvironment of Halmyris—A Roman Settlement in Southeast Romania: Archaeozoological and Phytolith Evidences" Diversity 15, no. 3: 373. https://doi.org/10.3390/d15030373