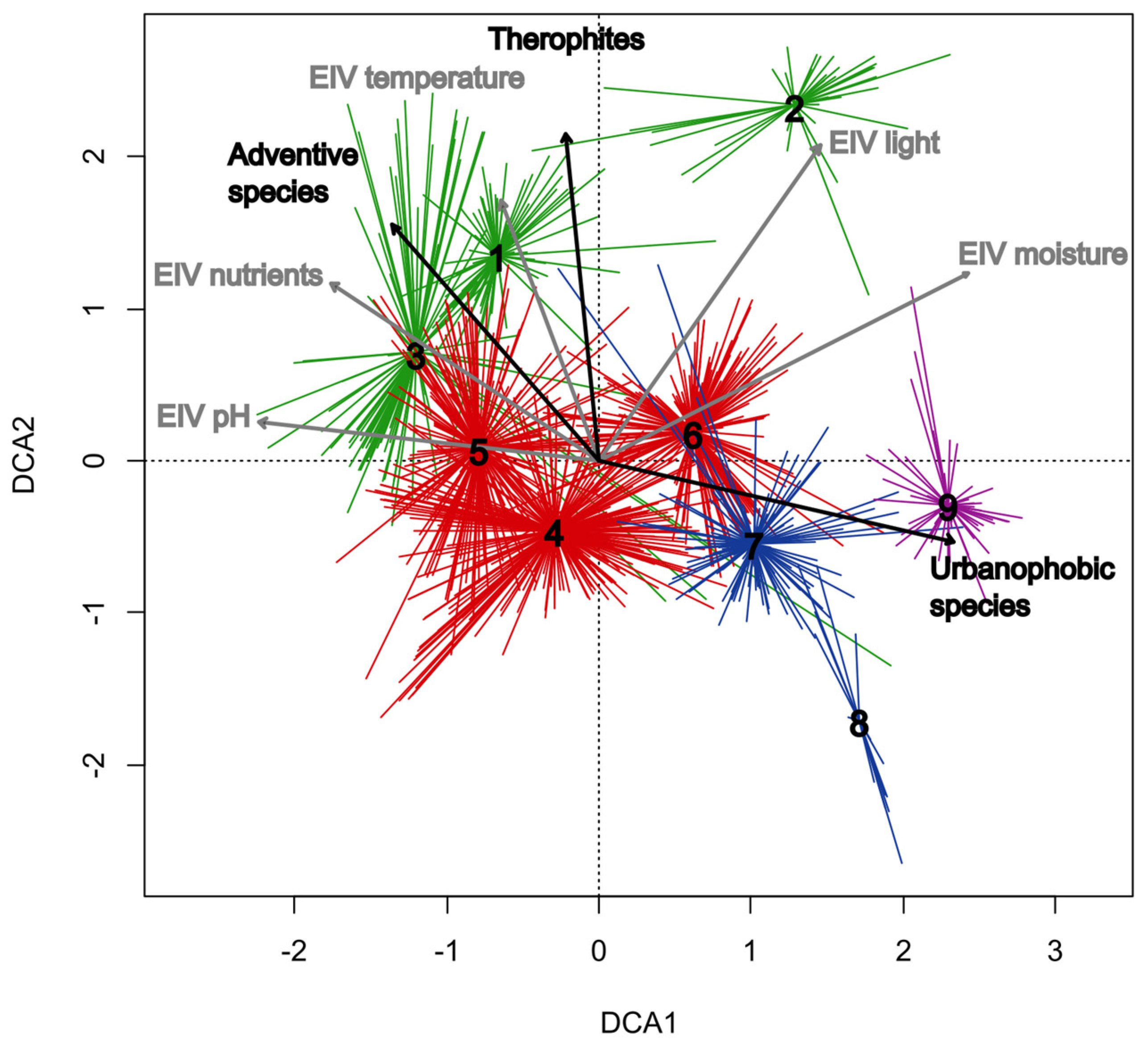
3.2. Overview of the Classified Communities
3.2.1. Salicetea purpureae Group of Clusters (Clusters 1–3; Table 1, Columns 1–3; Table A1)
This group of clusters consists of willow scrub and woodland communities that are found near stream banks or on regularly flooded floodplain sites. This class is represented by three alliances in the researched area, which was confirmed by the results of unsupervised classification of the first subdataset.
Syntaxonomy: Salicion albae
This cluster is mostly comprised of relevés of tall Salix alba-dominated communities. Salix euxina and Populus nigra are also sometimes present in the tree layer. Invasive species such as Acer negundo and Amorpha fruticose can often be important.
Diagnostic (bold) and constant species within the WFS: Salix alba, Salix euxina, Rubus caesius, Galium aparine, Phalaroides arundinacea, Urtica dioica.
Diagnostic (bold) and constant species within the Salicetea purpureae group of clusters: Acer negundo, Amorpha fruticosa, Euonymus europaeus, Rubus caesius, Salix alba, Salix euxina, Carex remota, Galium aparine, Glechoma hederacea, Iris pseudacorus, Rubus caesius, Phalaroides arundinacea, Urtica dioica.
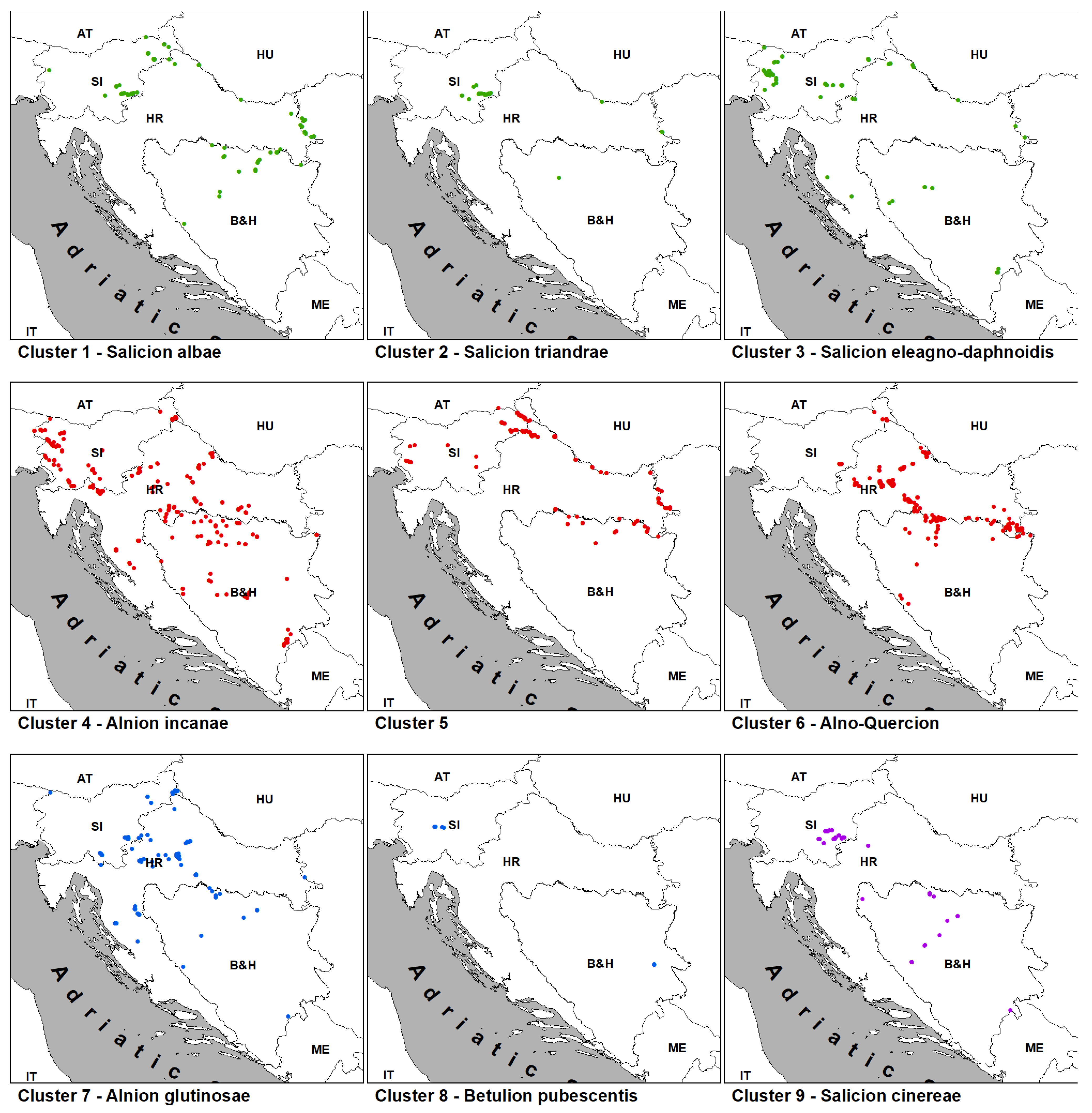
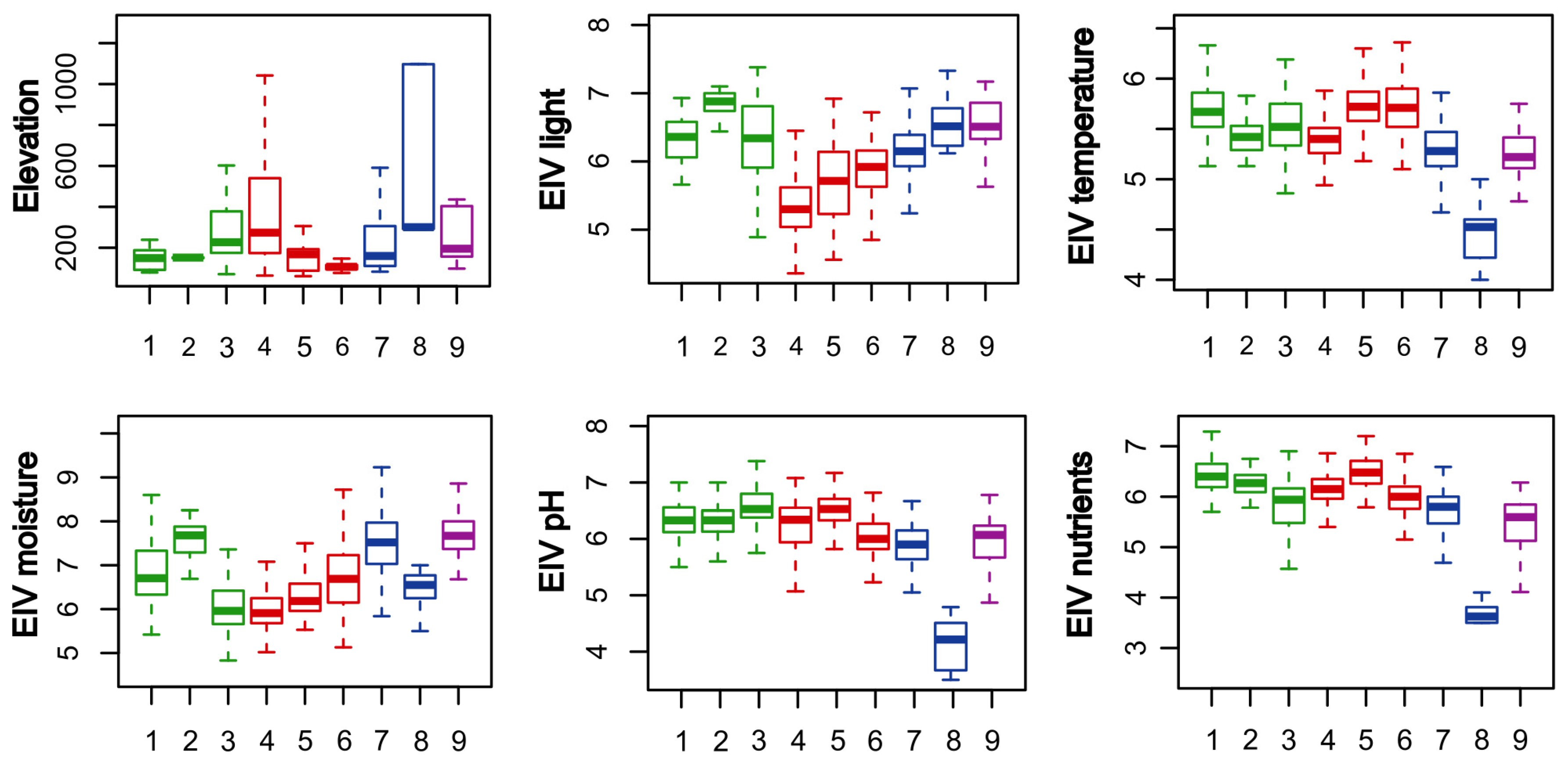
Ecology and distribution: These communities are usually located on the lower part of river terraces or in regularly flooded micro-depressions formed outside of the main riverbanks. In both cases, floodings with flowing water are regular and relatively long-lasting events. Soils are nutrient-rich fluvisols with a fine granulometric composition capable of retaining water for a long period of the year, although topsoil layers can dry out during summer. They are found in the floodplains of large lowland rivers throughout the whole area of research: Drava, Sava, Danube, Mura, Krka, Una, Vrbas, Bosna and Drina.
Published relevés from this cluster were mainly referred to as Salicetum albae, Galio-Salicetum albae and Salici-Populetum. Populus nigra dominated or co-dominated communities were not classified within this cluster or even this group of clusters. Additionally, 24 new and unpublished relevés from B&H were classified within this cluster.
Syntaxonomy: Salicion triandrae
This cluster consists of Salix triandra-dominated scrub, with Salix viminalis sometimes present. The tree layer is absent, while the height of stands is up to 5 m.
Diagnostic (bold) and constant species within the WFS: Salix triandra, Agrostis stolonifera agg., Bidens tripartitus, Calystegia sepium, Echinocystis lobata, Persicaria dubia, Phalaroides arundinacea, Rorippa amphibia, Rorippa sylvestris, Rumex crispus, Solanum dulcamara, Lythrum salicaria, Persicaria hydropiper, Urtica dioica.
Diagnostic (bold) and constant species within the Salicetea purpureae group of clusters: Salix triandra, Solanum dulcamara; Agrostis stolonifera agg., Alisma plantago-aquatica, Bidens tripartitus, Calystegia sepium, Echinocystis lobata, Galium palustre agg., Lythrum salicaria, Persicaria hydropiper, Phalaroides arundinacea, Rorippa amphibia, Rorippa sylvestris, Urtica dioica.
Ecology and distribution: These communities are usually located on the lowest part of river terraces along the slower downstream of large rivers. They form narrow vegetation strips along riverbanks and on sandbars, where they are under constant accumulation of new sandy and loamy sediment brought by the river current for as many as 100 days a year. With new material accumulating the ground gets higher, and the flood dynamics changes towards fewer days under flood. Hence, these short-lived pioneer communities, after no more than ten years, give away to the next stages in the succession. Although there is a lack of relevés from these communities, they are present in all three countries: Drava, Danube, Vrbas and Drina.
All published relevés from this cluster were originally assigned the name Salicetum triandrae. Only one new and unpublished relevé from B&H was classified within this cluster.
Syntaxonomy: Salicion eleagno-daphnoidis
This cluster comprises relevés of Salix eleagnos and/or Salix purpurea-dominated scrub. Diagnostic species are light-demanding species with moisture requirements varying from moisture-demanding to mesophilic species.
Diagnostic (bold) and constant species within the WFS: Clematis vitalba, Salix eleagnos, Salix purpurea, Centaurea nigrescens ssp. vochinensis, Chaerophyllum hirsutum, Galium mollugo, Helianthus tuberosus, Knautia drymeia s.lat., Lathyrus sylvestris, Melilotus albus, Mentha longifolia, Pastinaca sativa, Petasites hybridus, Petasites paradoxus, Peucedanum altissimum, Pimpinella major, Plantago lanceolata, Saponaria officinalis, Silene vulgaris, Taraxacum sect. Taraxacum, Tussilago farfara, Vicia cracca s.lat., Rubus caesius, Brachypodium sylvaticum.
Diagnostic (bold) and constant species within the Salicetea purpureae group of clusters: Acer pseudoplatanus, Alnus incana, Carpinus betulus, Clematis vitalba, Corylus avellana, Frangula alnus, Fraxinus excelsior, Hedera helix, Salix eleagnos, Salix purpurea, Salvia glutinosa, Ulmus glabra, Brachypodium sylvaticum, Chaerophyllum hirsutum, Cirsium oleraceum, Deschampsia cespitosa, Equisetum arvense, Erigeron annuus, Eupatorium cannabinum, Festuca gigantea, Galium mollugo, Geranium robertianum, Helianthus tuberosus, Heracleum sphondylium, Knautia drymeia s.lat., Lamium orvala, Lunaria rediviva, Melilotus albus, Mentha longifolia, Mycelis muralis, Pastinaca sativa, Petasites hybridus, Petasites paradoxus, Peucedanum altissimum, Pimpinella major, Ranunculus lanuginosus, Saponaria officinalis, Silene vulgaris, Stachys sylvatica, Taraxacum sect. Taraxacum, Tussilago farfara, Vicia cracca s.lat., Rubus caesius.
Ecology and distribution: These communities are usually developed on gravel or sandy beds of small and medium rivers with fast-flowing water and with regular and intense short floods. Fluctuations are intensified by pronounced drought periods that occur in summer caused by a significant drop in the water table, which is intensified by the inability of gravel and sand to retain water. Relevés are primarily concentrated in Slovenia, while scattered over only a couple of localities in Croatia and Bosnia and Herzegovina at different altitudes.
Published relevés from this cluster were originally assigned the following names: Salici-Myricarietum, Salicetum incano-purpureae, Lamio orvalae-Salicetum eleagni, Lamio orvalae-Salicetum purpureae, Carici-Salicetum myrsinifoliae, Salicetum purpureae, Salicetum cinereo-purpureae, Saponario-Salicetum. Additionally, nine new and unpublished relevés from B&H were classified within this cluster.
3.2.2. Alno glutinosae-Populetea albae Group of Clusters (Clusters 4–6; Table 1, Columns 4–6; Table A2)
The Alno glutinosae-Populetea albae group of clusters contains floodplain riparian alder–ash, elm–ash and oak forests on nutrient-rich soils and characterized by inter- and intra-annual fluctuations in the water level. This class is represented by three alliances in the researched area, which was confirmed by the results of unsupervised classification of the second subdataset.
Syntaxonomy: Alnion incanae s. str.
This cluster consists of forests dominated by Alnus incana and/or A. glutinosa, as well as Salix eleagnos, and sometimes also S. alba and/or S. euxina. Trees related to mesophilous and ravine forests, such as Acer pseudoplatanus, Fagus sylvatica, Fraxinus excelsior and Ulmus glabra, are also frequent. The understory is also a mixture of hygrophilous, mesophilous and nitrophilous species.
Diagnostic (bold) and constant species within the WFS: Acer campestre, Acer pseudoplatanus, Alnus glutinosa agg., Corylus avellana, Fraxinus excelsior, Sambucus nigra, Aegopodium podagraria, Brachypodium sylvaticum, Cardamine bulbifera, Carex pendula, Carex sylvatica, Geum urbanum, Lamium galeobdolon agg., Lamium orvala, Lunaria rediviva, Mercurialis perennis, Oxalis acetosella, Primula acaulis, Symphytum tuberosum agg., Cornus sanguinea, Euonymus europaeus, Rubus caesius, Urtica dioica.
Diagnostic (bold) and constant species within the Alno glutinosae-Populetea albae group of clusters: Acer pseudoplatanus, Alnus glutinosa agg., Corylus avellana, Fagus sylvatica, Fraxinus excelsior, Salvia glutinosa, Sambucus nigra, Ulmus glabra, Aegopodium podagraria, Angelica sylvestris, Brachypodium sylvaticum, Cardamine bulbifera, Chaerophyllum hirsutum, Cirsium oleraceum, Equisetum arvense, Knautia drymeia s.lat., Lamium galeobdolon agg., Lamium orvala, Lunaria rediviva, Mercurialis perennis, Petasites hybridus, Primula acaulis, Ranunculus lanuginosus, Acer campestre, Cornus sanguinea, Euonymus europaeus, Rubus caesius, Urtica dioica.
Ecology and distribution: Stands classified in this cluster occur on stream banks and at headwater seepages, which are usually flooded in spring for several days or weeks and usually dry out during the summer. Stands dominated by Alnus incana and/or A. glutinosa, and sometimes Salix eleagnos, S. alba and S. euxina, together with Acer pseudoplatanus, Fagus sylvatica, Fraxinus excelsior and Ulmus glabra, usually occupy banks of small to medium-sized streams of the colline to montane belt, on stony to sandy, nutrient rich colluvial soil. On the other hand, stands dominated by Alnus glutinosa are mainly confined to lower and mid-elevations along smaller streams or at headwater seepages with sandy to loamy, slightly acidic and moderately rich soil. They are common in suitable habitats throughout the study area.
Published relevés from this cluster were referred to as: Alnetum incanae, Lamio orvalae-Alnetum incanae, Carici acutiformis-Alnetum glutinosae, Carici brizoidis-Alnetum glutinosae p.p., Carici elongatae-Alnetum p.p., Frangulo-Alnetum glutinosae, Lamio orvalae-Alnetum glutinosae, Pruno padi-Fraxinetum, Stellario-Alnetum glutinosae, Lamio orvalae-Salicetum eleagni, Lamio orvalae-Salicetum albae ranunculetosum lanuginosae. Additionally, 87 new and unpublished relevés from B&H were classified within this cluster.
Syntaxonomy: not assigned
This cluster contains floodplain hardwood (Ulmus laevis, Fraxinus angustifolia and sometimes Quercus robur) and/or poplar (Populus alba and P. nigra) forests. The shrub layer is well developed, with Cornus sanguinea, Sambucus nigra, Prunus padus, Euonymus europaeus and Prunus padus, among others, while the herb layer is typically made of nemoral mesophilous and hygromesophilous species. Invasive alien species such as Acer negundo, Solidago gigantea, Impatiens glandulifera and Robinia pseudoacacia can be common.
Diagnostic (bold) and constant species within the WFS: Acer negundo, Populus alba, Populus nigra, Prunus padus, Ulmus laevis, Anemone ranunculoides, Arum maculatum, Galium aparine, Leucojum vernum, Pulmonaria officinalis agg., Solidago gigantea, Veronica hederifolia, Cornus sanguinea, Rubus caesius, Sambucus nigra, Aegopodium podagraria, Urtica dioica.
Diagnostic (bold) and constant species within the Alno glutinosae-Populetea albae group of clusters: Acer negundo, Populus alba, Populus nigra, Prunus padus, Robinia pseudoacacia, Salix alba, Ulmus laevis, Galium aparine, Impatiens glandulifera, Solidago gigantea, Veronica hederifolia, Cornus sanguinea, Rubus caesius, Sambucus nigra, Aegopodium podagraria, Urtica dioica.
Ecology and distribution: These forests are developed on floodplains of the middle and lower reaches of the largest rivers in the study area (Sava, Drava, Danube, Una, Vrbas, Bosna and Drina). They form on alluvial deposits on the highest terraces within the floodplain, which are only under water during the highest, mainly spring floods. The soil is mainly sandy and, due to the pronounced water regime dynamics, it can become very dry during the summer months.
Published relevés from this cluster were referred to as: Equiseto-Alnetum incanae, Fraxino-Ulmetum effusae, Salicetum albae p.p. (polidominant communities), Lamio orvalae-Salicetum albae caricetosum pendulae, Populetum nigro-albae, Salici-Populetum and Carduo crispi-Populetum nigrae. Additionally, 22 new and unpublished relevés from B&H were classified within this cluster.
Syntaxonomy: Alno-Quercion roboris
The cluster encompasses floodplain hardwood forests dominated by Quercus robur and/or Fraxinus angustifolia s.lat. with Ulmus minor, Alnus glutinosa and Acer campestre, frequently admixed. In some cases, Alnus glutinosa has the role of edifier (probably in secondary succession stages). The shrub layer is not as developed as in Cluster 5, with Crataegus sp., Frangula alnus and Cornus sanguinea being the most important, with a frequency of around 40%. The herb layer is represented mainly by hygrophilous and hygromesophilous forest species.
Diagnostic (bold) and constant species within the WFS: Acer tataricum, Crataegus laevigata, Fraxinus angustifolia s.lat., Quercus robur, Ulmus minor, Carex remota, Carex strigosa, Glechoma hederacea, Lysimachia nummularia, Rumex sanguineus, Stachys palustris, Rubus caesius, Galium palustre agg., Iris pseudacorus, Urtica dioica.
Diagnostic (bold) and constant species within the Alno glutinosae-Populetea albae group of clusters: Acer tataricum, Crataegus laevigata, Fraxinus angustifolia s.lat., Quercus robur, Ulmus minor, Bidens tripartitus, Carex elongata, Carex remota, Carex riparia, Galium palustre agg., Glechoma hederacea, Iris pseudacorus, Leucojum aestivum, Lycopus europaeus, Lysimachia nummularia, Lythrum salicaria, Myosotis palustris agg., Persicaria hydropiper, Rumex sanguineus, Stachys palustris, Rubus caesius, Urtica dioica.
Ecology and distribution: These communities are mostly distributed in lowlands but are not confined to floodplains, since they can be quite distant from a river. The commonality of these forests is the presence of stagnant water at the surface during a longer or shorter time during the year (mostly in spring and autumn), which is influenced by the flat relief and clayey soil. When within a floodplain, they develop in a transitional zone between the highest river terraces (Cluster 5) and depressions with stagnant water (Cluster 7). The fluctuation of water level in the soil can vary greatly and is often a key factor determining the type of community to develop. In the period between floods, the soil may be dry or wet, depending on the flood duration and groundwater table. In the research area, these communities are widespread within alluvia of large rivers, but also on flat, periodically waterlogged, terrains outside the alluvium, such as karst poljes (e.g., Livanjsko polje in B&H).
Published relevés from this cluster were referred to as:
Genisto elatae-Quercetum,
Leucojo-Fraxinetum,
Frangulo-Alnetum glutinosae,
Carici elongatae-Alnetum p.p. (less swampy relevés),
Carici brizoidis-Alnetum glutinosae p.p. (only two relevés from original description [
34]). Additionally, 53 new and unpublished relevés from B&H were classified within this cluster.
3.2.3. Alnetea glutinosae Group of Clusters (Clusters 7–8; Table 1, Columns 7–8; Table A3)
The Alnetea glutinosae group of clusters consists of swamp alder forests and birch wooded mires on gleic soils of permanently waterlogged sites. Swamp species tolerant of oxidative stress at permanently waterlogged sites dominate in the herb layer. This class is represented in the researched area by two alliances, which was confirmed by the results of unsupervised classification of the third subdataset.
Syntaxonomy: Alnion glutinosae
Cluster 7 encompasses relevés of mesotrophic regularly flooded alder carr dominated by Alnus glutinosa and sometimes accompanied by Quercus robur and/or Fraxinus angustifolia. The understory is mainly represented by tall sedges (Carex elongate, C. acutiformis and C. riparia) and other wetland plant species.
Diagnostic (bold) and constant species within the WFS: Alnus glutinosa agg., Carex elongata, Carex riparia, Carex vesicaria, Lycopus europaeus, Peucedanum palustre, Valeriana dioica s.lat., Frangula alnus, Solanum dulcamara, Viburnum opulus, Dryopteris carthusiana, Galium palustre agg., Iris pseudacorus, Lysimachia vulgaris.
Diagnostic (bold) and constant species within the Alnetea glutinosae group of clusters: Alnus glutinosa agg., Rubus caesius, Carex elongata, Filipendula ulmaria, Iris pseudacorus, Lycopus europaeus, Lythrum salicaria, Urtica dioica, Frangula alnus, Solanum dulcamara, Viburnum opulus, Dryopteris carthusiana, Galium palustre agg., Lysimachia vulgaris, Peucedanum palustre.
Ecology and distribution: In the study area, these forests develop in shallow waterlogged depressions usually inundated by groundwater for considerable parts of the growing season. Soils lack well-aerated horizons and are often characterized by a significant accumulation of undecomposed organic matter. Although this habitat often occurs on sites not related to rivers, they can also be found along oxbows of large rivers (Sava, Vrbas, Bosna, Drina).
Published relevés from this cluster were referred to as: Carici elongatae-Alnetum, Carici acutiformis-Alnetum glutinosae, Carici brizoidis-Alnetum glutinosae p.p., Leucojo-Fraxinetum p.p., Genisto elatae-Quercetum roboris p.p. and Pseudostellario-Quercetum roboris p.p. (the last four names are related only to several relevés with a pronounced swamp character and dominated by Alnus glutinosa (besides Q. robur and F. angustifolia)). Additionally, nine new and unpublished relevés from B&H were classified within this cluster.
Syntaxonomy: Betulion pubescentis
Cluster 8 contains acidophilous and poor in nutrients forests on bog, dominated by Betula pubescens and sometimes accompanied by Pinus sylvestris or Betula pendula. The herb layer is represented by acidophilous species and species of nutrient-poor soils. The moss layer is well developed and with a significant participation of various Sphagnum species.
Diagnostic (bold) and constant species within the WFS: Betula pendula, Betula pubescens, Frangula alnus, Picea abies, Pinus sylvestris, Populus tremula, Rubus hirtus s.lat., Salix caprea, Salix pentandra, Sorbus aucuparia, Vaccinium myrtillus, Salix aurita, Lonicera nigra, Vaccinium vitis-idaea, Agrostis canina, Aruncus dioicus, Calamagrostis villosa, Calluna vulgaris, Carex echinata, Carex pallescens, Carex paniculata, Carex rostrata, Carex spicata, Cirsium palustre, Danthonia decumbens, Dryopteris carthusiana, Eleocharis palustris, Epilobium palustre, Equisetum palustre, Equisetum sylvaticum, Gentiana pneumonanthe, Knautia sarajevensis, Molinia caerulea agg., Orthilia secunda, Parnassia palustris, Persicaria bistorta, Pyrola media, Viola canina.
Diagnostic (bold) and constant species within the Alnetea glutinosae group of clusters: Betula pendula, Betula pubescens, Pinus sylvestris, Populus tremula, Rubus hirtus s.lat., Salix pentandra, Sorbus aucuparia, Salix aurita, Lonicera nigra, Agrostis canina, Aruncus dioicus, Calamagrostis villosa, Calluna vulgaris, Carex echinata, Carex rostrata, Carex spicata, Cirsium palustre, Danthonia decumbens, Epilobium palustre, Equisetum palustre, Equisetum sylvaticum, Knautia sarajevensis, Molinia caerulea agg., Parnassia palustris, Pyrola media, Viola canina, Frangula alnus, Dryopteris carthusiana.
Ecology and distribution: These communities are far to the south of the center of their distribution, and there are only a few relict sites with this habitat type in the research area (Slovenia and Bosnia and Herzegovina). The stands occur on acidic and nutrient poor waterlogged habitats with Sphagnum peat.
Published relevés from this cluster were referred to as: Pineto-Betuletum pubescentis, Sphagno nemorei-Betuletum pubescentis and Betulo-Quercetum roboris.
3.2.4. Franguletea Group of Clusters (Cluster 9; Table 1, Column 9)
There is only one cluster in this group of willow swamp scrub.
Syntaxonomy: Salicion cinereae
Cluster 9 encompasses willow carr dominated by Salix cinerea, sometimes accompanied by S. pentandra. The herb layer is heterogeneous, represented by hygrophilous species of wet meadows and swamps.
Diagnostic (bold) and constant species within the WFS: Salix cinerea, Carex nigra, Carex panicea, Filipendula ulmaria, Rhinanthus rumelicus, Succisella inflexa, Solanum dulcamara, Galium palustre agg., Lycopus europaeus, Lythrum salicaria, Ranunculus repens.
Ecology and distribution: This scrub can be found in river alluviums, wet meadows, fens and lake shores throughout the research area. They are a stage in the natural succession of lakes and fens, as well as the secondary succession following the abandonment of wet meadows or the removal of alder carrs.
Published relevés from this cluster were referred to as Salicetum cinereae. Additionally, 23 new and unpublished relevés from B&H were classified within this cluster.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}



