

The Addition of a High Concentration of Phosphorus Reduces the Diversity of Arbuscular Mycorrhizal Fungi in Temperate Agroecosystems

Abstract
:1. Introduction
2. Material and Methods Introduction
2.1. Study Site and Physical and Chemical Properties of Soil
2.2. Experimental Design, Sample Collection, and Measured Soil Properties
2.3. DNA Extraction and Illumina MiSeq Sequencing
2.4. Statistical Analysis
3. Results
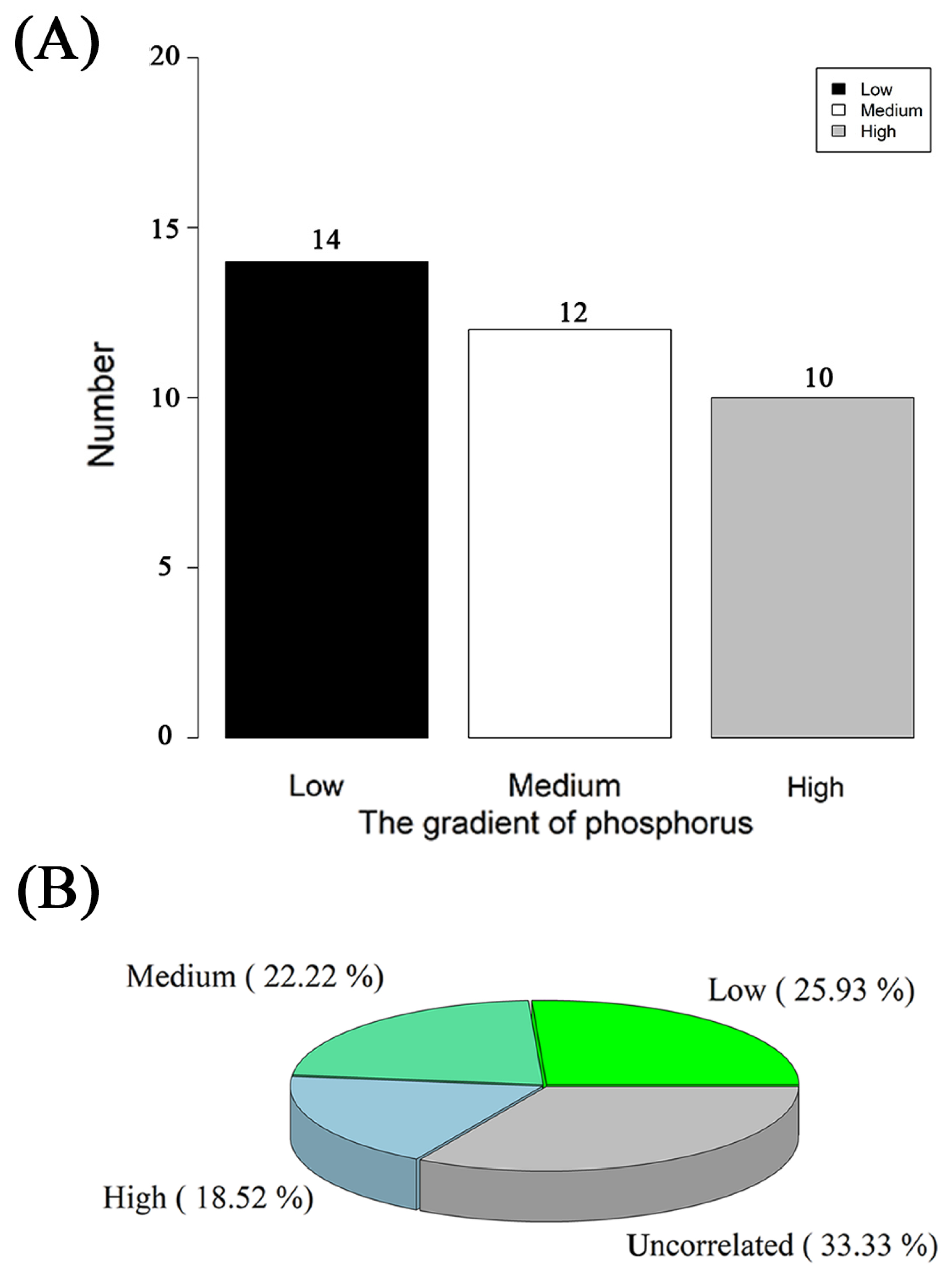
3.1. Species Diversity of AMF under Different P Gradients
3.2. Correlation of AMF Community under Three P Concentrations
3.3. Relative Importance of Deterministic and Stochastic Processes along the P Gradient
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tilman, D. Global environmental impacts of agricultural expansion: The need for sustainable and efficient practices. Proc. Natl. Acad. Sci. USA 1999, 96, 5995–6000. [Google Scholar] [CrossRef] [PubMed]
- Childers, D.; Corman, J.; Edwards, M.; Elser, J. Sustainability challenges of phosphorus and food: Solutions from closing the human phosphorus cycle. BioScience 2011, 61, 117–124. [Google Scholar] [CrossRef]
- Li, H.; Huang, G.; Meng, Q.; Ma, L.; Yuan, L.; Wang, F.; Zhang, F. Integrated soil and plant phosphorus management for crop and environment in China: A review. Plant Soil 2011, 349, 157–167. [Google Scholar] [CrossRef]
- Zhang, F.S.; Cui, Z.L.; Chen, X.P.; Ju, X.T.; Shen, J.B.; Chen, Q.; Liu, X.J.; Zhang, W.F.; Mi, G.H.; Fan, M.S.; et al. Integrated Nutrient Management for Food Security and Environmental Quality in China. Adv. Agron. 2012, 116, 1–40. [Google Scholar] [CrossRef]
- Drinkwater, L.E.; Snapp, S.S. Nutrients in agroecosystems: Rethinking the management paradigm. Adv. Agron. 2007, 92, 163–186. [Google Scholar] [CrossRef]
- Qin, B.; Gao, G.; Zhu, G.; Zhang, Y.; Song, Y.; Tang, X.; Xu, H.; Deng, J. Lake eutrophication and its ecosystem response. Chin. Sci. Bull. 2013, 58, 961–970. [Google Scholar] [CrossRef]
- Zhao, X.; Min, J.; Wang, S.; Shi, W.; Xing, G. Further understanding of nitrous oxide emission from paddy fields under rice/wheat rotation in south China. J. Geophys. Res. 2011, 116, G02016. [Google Scholar] [CrossRef]
- Guo, J.H.; Liu, X.J.; Zhang, Y.; Shen, J.L.; Han, W.X.; Zhang, W.F.; Christie, P.; Goulding, K.W.T.; Vitousek, P.M.; Zhang, F.S. Significant Acidification in Major Chinese Croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef]
- NBSC (National Bureau of Statistics of China). China Statistical Yearbook; China Statistics Press: Beijing, China, 2015. [Google Scholar]
- Zhou, Y.; He, Z.H.; Sui, X.X.; Xia, X.C.; Zhang, X.K.; Zhang, G.S. Genetic Improvement of Grain Yield and Associated Traits in the Northern China Winter Wheat Region from 1960 to 2000. Crop Sci. 2007, 47, 245–253. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, E.; Yang, X.; Wang, J. Contributions of climatic and crop varietal changes to crop production in the North China Plain, since 1980. Glob. Chang. Biol. 2009, 16, 2287–2299. [Google Scholar] [CrossRef]
- Ngoze, S.; Riha, S.; Lehmann, J.; Verchot, L.; Kinyangi, J.; Mbuga, D.; Pell, A. Nutrient constraints to tropical agroecosystem productivity in long-term degrading soils. Glob. Chang. Biol. 2008, 14, 2810–2822. [Google Scholar] [CrossRef]
- Demay, J.; Ringeval, B.; Pellerin, S.; Nesme, T. Half of global agricultural soil phosphorus fertility derived from anthropogenic sources. Nat. Geosci. 2023, 16, 69–74. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Van Kauwenbergh, J. World Phosphorus Rock Reserves and Resources; International Fertilizer Development Center: Muscle Shoals, AL, USA, 2010. [Google Scholar]
- Kucey, R.M.; Janzen, H.H. Effects of VAM and reduced nutrient availability on growth and phosphorus and micronutrient uptake of wheat and field beans under greenhouse conditions. Plant Soil 1987, 104, 71–78. [Google Scholar] [CrossRef]
- Purakayastha, T.; Chhonkar, P. Influence of vesicular-arbuscular mycorrhizal fungi (Glomus etunicatum L.) on mobilization of zinc in wetland rice (Oryza sativa L.). Biol. Fertil. Soils 2001, 33, 323–327. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. (Eds.) Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, UK, 2008; pp. 13–41. [Google Scholar]
- Hajiboland, R.; Aliasgharzad, N.; Barzeghar, R. Influence of arbuscular mycorrhizal fungi on uptake of Zn and P by two contrasting rice genotypes. Plant Soil Environ. 2009, 55, 93–100. [Google Scholar] [CrossRef]
- Panneerselvam, P.; Saritha, B.; Mohandas, S.; Upreti, K.K.; Poovarasan, S.; Sulladmath, V.V.; Venugopalan, R. Effect of mycorrhiza-associated bacteria on enhancing colonization and sporulation of Glomus mosseae and growth promotion in sapota (Manilkara achras (Mill) Forsberg) seedlings. Biol. Agric. Hortic. 2013, 29, 118–131. [Google Scholar] [CrossRef]
- Lambers, H.; Teste, F.P. Interactions between arbuscular mycorrhizal and non-mycorrhizal plants: Do non-mycorrhizal species at both extremes of nutrient availability play the same game? Plant Cell Environ. 2013, 36, 1911–1915. [Google Scholar] [CrossRef]
- Bona, E.; Scarafoni, A.; Marsano, F.; Boatti, L.; Copetta, A.; Massa, N.; Gamalero, E.; D’Agostino, G.; Cesaro, P.; Cavaletto, M.; et al. Arbuscular mycorrhizal symbiosis affects the grain proteome of Zea mays: A field study. Sci. Rep. 2016, 6, 26439. [Google Scholar] [CrossRef]
- Fiorilli, V.; Vannini, C.; Ortolani, F.; Garcia-Seco, D.; Chiapello, M.; Novero, M.; Domingo, G.; Terzi, V.; Morcia, C.; Bagnaresi, P.; et al. Omics approaches revealed how arbuscular mycorrhizal symbiosis enhances yield and resistance to leaf pathogen in wheat. Sci. Rep. 2018, 8, 9625. [Google Scholar] [CrossRef]
- Lekberg, Y.; Koide, R.T.; Rohr, J.R.; Aldrich-Wolfe, L.; Morton, J.B. Role of niche restrictions and dispersal in the composition of arbuscular mycorrhizal fungal communities. J. Ecol. 2007, 95, 95–105. [Google Scholar] [CrossRef]
- Fitzsimons, M.S.; Miller, R.M.; Jastrow, J.D. Scale-dependent niche axes of arbuscular mycorrhizal fungi. Oecologia 2008, 158, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Wang, Y.; Liu, M.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 2020, 10, 12260. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.C.; Gehring, C.A. Mycorrhizas: Symbiotic Mediators of Rhizosphere and Ecosystem Processes. In The Rhizosphere; Elsevier Inc.: Amsterdam, The Netherlands, 2007; pp. 73–100. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Nelson, M.; Helgason, T.; Dytham, C.; Fitter, A.H. Relative roles of niche and neutral processes in structuring a soil microbial community. ISME J. 2010, 4, 337–345. [Google Scholar] [CrossRef]
- Ofiţyeru, I.D.; Lunn, M.K.; Curtis, T.P.; Wells, G.F.; Criddle, C.S.; Francis, C.A.; Sloan, W.T. Combined niche and neutral effects in a microbial wastewater treatment community. Proc. Natl. Acad. Sci. USA 2010, 107, 15345–15350. [Google Scholar] [CrossRef] [PubMed]
- Langenheder, S.; Szekely, A.J. Species sorting and neutral processes are both important during the initial assembly of bacterial communities. ISME J. 2011, 5, 1086–1094. [Google Scholar] [CrossRef]
- Chase, J.M. Stochastic Community Assembly Causes Higher Biodiversity in More Productive Environments. Science 2010, 328, 1388–1391. [Google Scholar] [CrossRef]
- Hubbell, S.P. The Unified Neutral Theory of Biodiversity and Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Chave, J. Neutral theory and community ecology. Ecol. Lett. 2004, 7, 241–253. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Phil. Trans. R. Soc. B 2011, 366, 2351–2363. [Google Scholar] [CrossRef]
- Fukami, T. Historical contingency in community assembly: Integrating niches, species pools, and priority efects. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 1–23. [Google Scholar] [CrossRef]
- Zhou, J.; Ning, D. Stochastic community assembly: Does it matter in microbial ecology? MMBR 2017, 81, e00002-17. [Google Scholar] [CrossRef] [PubMed]
- Vályi, K.; Mardhiah, U.; Rillig, M.C.; Hempel, S. Community assembly and coexistence in communities of arbuscular mycorrhizal fungi. ISME J. 2016, 10, 2341–2351. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Moora, M.; Jairus, T.; Vasar, M.; Öpik, M.; Zobel, M. Hierarchical assembly rules in arbuscular mycorrhizal (AM) fungal communities. Soil Biol. Biochem. 2016, 97, 63–70. [Google Scholar] [CrossRef]
- Saks, Ü.; Davison, J.; Öpik, M.; Vasar, M.; Moora, M.; Zobel, M. Root-colonizing and soil-borne communities of arbuscular mycorrhizal fungi in a temperate forest understorey. Botany 2014, 92, 277–285. [Google Scholar] [CrossRef]
- Ma, X.; Geng, Q.; Zhang, H.; Bian, C.; Chen, H.Y.; Jiang, D.; Xu, X. Global negative efects of nutrient enrichment on arbuscular mycorrhizal fungi, plant diversity and ecosystem multifunctionality. New Phytol. 2021, 229, 2957–2969. [Google Scholar] [CrossRef] [PubMed]
- Sheng, K.; Li, Y.; Zhang, D.; Si, Y.; Chu, L.; Li, L.; Wang, D.; Wang, Y. Effects of nitrogen, phosphorus and potassium combined application on yield and physiological characteristics of Jerusalem artichoke. Grassland 2022, 44, 81–90. [Google Scholar] [CrossRef]
- Du, S.; Gao, X.Z.; Li, K. Technical Specification of Soil Analysis, 2nd ed.; China Agriculture Press: Beijing, China, 2006. [Google Scholar]
- Lumini, E.; Orgiazzi, A.; Borriello, R.; Bonfante, P.; Bianciotto, V. Disclosing arbuscular mycorrhizal fungal biodiversity in soil through a land-use gradient using a pyrosequencing approach. Environ. Microbiol. 2010, 12, 2165–2179. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.; Chen, B.; Li, H. Specificity and selectivity of arbuscular mycorrhizal fungal polymerase chain reaction primers in soil samples by clone library analyses. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2016, 66, 333–339. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Ashton, P.D.; Aziz, N.; Feng, G.; Nelson, M.; Dytham, C.; Fitter, A.H.; Helgason, T. Distinct Seasonal Assemblages of Arbuscular Mycorrhizal Fungi Revealed by Massively Parallel Pyrosequencing. New Phytol. 2011, 190, 794–804. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.2-1. 2015. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1654-1103.2003.tb02228.x (accessed on 19 September 2021).
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize Implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef]
- Pierre, L. Studying beta diversity: Ecological variation partitioning by multiple regression and canonical analysis. Chin. J. Plant Ecol. 2007, 31, 976–981. [Google Scholar] [CrossRef]
- Harms, K.E.; Condit, R.S.; Hubbell, S.P.; Foster, R.B. Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, Z.; Bi, S.; Wang, X.; Ye, Y.; Svenning, J.-C. Macrofungal species distributions depend on habitat partitioning of topography, light, and vegetation in a temperate mountain forest. Sci. Rep. 2018, 8, 13589. [Google Scholar] [CrossRef]
- Xi, J.; Shao, Y.; Li, Z.; Zhao, P.; Ye, Y.; Li, W.; Chen, Y.; Yuan, Z. Distribution of Woody Plant Species among Different Disturbance Regimes of Forests in a Temperate Deciduous Broad-Leaved Forest. Front. Plant Sci. 2021, 12, 618524. [Google Scholar] [CrossRef]
- Chen, Y.; Xi, J.; Xiao, M.; Wang, S.; Chen, W.; Liu, F.; Shao, Y.; Yuan, Z. Soil fungal communities show more specificity than bacteria for plant species composition in a temperate forest in China. BMC Microbiol. 2022, 22, 208. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.; Deng, Y.; Tiedje, J.M.; Zhou, J. A general framework for quantitatively assessing ecological stochasticity. Proc. Natl. Acad. Sci. USA 2019, 116, 16892–16898. [Google Scholar] [CrossRef] [PubMed]
- Stone, L.; Roberts, A. The checkerboard score and species distributions. Oecologia 1990, 85, 74–79. [Google Scholar] [CrossRef]
- Swenson, N.G. Functional and Phylogenetic Ecology in R; Springer: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Tian, D.; Zhang, Y.; Mu, Y.; Zhou, Y.; Zhang, C.; Liu, J. The effect of drip irrigation and drip fertigation on N2O and NO emissions, water saving and grain yields in a maize field in the North China Plain. Sci. Total Environ. 2017, 575, 1034–1040. [Google Scholar] [CrossRef]
- Zhang, X.; Bol, R.; Rahn, C.; Xiao, G.; Meng, F.; Wu, W. Agricultural sustainable intensification improved nitrogen use efficiency and maintained high crop yield during 1980–2014 in Northern China. Sci. Total Environ. 2017, 596–597, 61–68. [Google Scholar] [CrossRef]
- Li, X.; Hou, X.; Mu, H.; Li, X.; Guo, F. P fertilization effects on the accumulation, transformation and availability of soil phosphorus. Acta Prataculturae Sin. 2015, 24, 218–224. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Jiang, S.; Deng, Y.; Christie, P.; Murray, P.J.; Li, X.; Zhang, J. Arbuscular mycorrhizal fungi in soil and roots respond differently to phosphorus inputs in an intensively managed calcareous agricultural soil. Sci. Rep. 2016, 6, 24902. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.C.; Angelard, C.; Sanders, I.R.; Kiers, E.T. Predicting community and ecosystem outcomes of mycorrhizal responses to global change. Ecol. Lett. 2013, 16, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Kahiluoto, H.; Ketoja, E.; Vestberg, M.; Saarela, I. Promotion of AM utilization through reduced P fertilization 2. Field studies. Plant Soil. 2001, 231, 65–79. [Google Scholar] [CrossRef]
- Camenzind, T.; Hempel, S.; Homeier, J.; Horn, S.; Velescu, A.; Wilcke, W.; Rillig, M.C. Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Chang. Biol. 2014, 20, 3646–3659. [Google Scholar] [CrossRef]
- Santos, J.C.; Finlay, R.D.; Tehler, A. Molecular analysis of arbuscular mycorrhizal fungi colonising a semi-natural grassland along a fertilisation gradient. New Phytol. 2006, 172, 159–168. [Google Scholar] [CrossRef]
- Alguacil, M.D.; Lozano, Z.; Campoy, M.; Roldán, A. Phosphorus fertilisation management modifies the biodiversity of AM fungi in a tropical savanna forage system. Soil Biol. Biochem. 2010, 42, 1114–1122. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, H.; Feng, G.; Christie, P.; Zhang, J.; Li, X.; Gai, J. Soil phosphorus availability modifies the relationship between AM fungal diversity and mycorrhizal benefits to maize in an agricultural soil. Soil Biol. Biochem. 2020, 144, 107790. [Google Scholar] [CrossRef]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef]
- Johnson, N.C. Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales. New Phytol. 2010, 185, 631–647. [Google Scholar] [CrossRef]
- Olsson, P.A.; Rahm, J.; Aliasgharzad, N. Carbon dynamics in mycorrhizal symbioses is linked to carbon costs and phosphorus benefits. FEMS Microbiol. Ecol 2010, 72, 125–131. [Google Scholar] [CrossRef]
- Werner, G.D.A.; Kiers, E.T. Partner selection in the mycorrhizal mutualism. New Phytol. 2015, 205, 1437–1442. [Google Scholar] [CrossRef]
- Johnson, D.; Leake, J.R.; Read, D.J. Novel in-growth core system enables functional studies of grassland mycorrhizal mycelial networks. New Phytol. 2001, 152, 555–562. [Google Scholar] [CrossRef]
- Koide, R.T.; Mosse, B. A history of research on arbuscular mycorrhiza. Mycorrhiza 2004, 14, 145–163. [Google Scholar] [CrossRef]
- Lambers, H.; Martinoia, E.; Renton, M. Plant adaptations to severely phosphorus-impoverished soils. Curr. Opin. Plant Biol. 2015, 25, 23–31. [Google Scholar] [CrossRef]
- Clavel, J.; Julliard, R.; Devictor, V. Worldwide decline of specialist species: Toward a global functional homogenization? Front. Ecol. Environ. 2011, 9, 222–228. [Google Scholar] [CrossRef]
- Xu, J.; Liu, S.; Song, S.; Guo, H.; Tang, J.; Yong, J.W.; Ma, Y.; Chen, X. Arbuscular mycorrhizal fungi influence decomposition and the associated soil microbial community under different soil phosphorus availability. Soil Biol. Biochem. 2018, 120, 181–190. [Google Scholar] [CrossRef]
- Olsson, P.A.; Baath, E.; Jakobsen, I. Phosphorus effects on the mycelium and storage structures of an arbuscular mycorrhizal fungus as studied in the soil and roots by analysis of Fatty Acid signatures. Appl. Environ. Microbiol. 1997, 63, 3531–3538. [Google Scholar] [CrossRef]
- Gosling, P.; Andrew, M.; Maude, P.; Hammond, J.P.; Bending, G.D. Contrasting arbuscular mycorrhizal communities colonizing different host plants show a similar response to a soil phosphorus concentration gradient. New Phytol. 2013, 198, 546–556. [Google Scholar] [CrossRef]
- Beauregard, M.S.; Gauthier, M.P.; Hamel, C.; Zhang, T.; Welacky, T.; Tan, C.S.; St-Arnaud, M. Various forms of organic and inorganic P fertilizers did not negatively affect soil- and root-inhabiting AM fungi in a maize-soybean rotation system. Mycorrhiza 2013, 23, 143–154. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical and Chemical Properties | Unit | Amount |
|---|---|---|
| pH | / | 7.8 |
| organic matter | g·kg−1 | 17.61 |
| total nitrogen (TN) | g·kg−1 | 0.82 |
| total phosphorus (TP) | g·kg−1 | 0.43 |
| available nitrogen (AN) | mg·kg−1 | 84.20 |
| available nitrogen (AP) | mg·kg−1 | 18.2 |
| available kalium (AK) | mg·kg−1 | 108.00 |
| Treatment | Low P (kg/hm2) | Medium P (kg/hm2) | High P (kg/hm2) |
|---|---|---|---|
| N | 150 | 150 | 150 |
| P(P2O5) | 20 | 60 | 120 |
| K(K2O) | 90 | 90 | 90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, F.; Wang, S.; Yuan, Y.; Chen, Y.; Guo, E.; Li, Y. The Addition of a High Concentration of Phosphorus Reduces the Diversity of Arbuscular Mycorrhizal Fungi in Temperate Agroecosystems. Diversity 2023, 15, 1045. https://doi.org/10.3390/d15101045
Miao F, Wang S, Yuan Y, Chen Y, Guo E, Li Y. The Addition of a High Concentration of Phosphorus Reduces the Diversity of Arbuscular Mycorrhizal Fungi in Temperate Agroecosystems. Diversity. 2023; 15(10):1045. https://doi.org/10.3390/d15101045
Chicago/Turabian StyleMiao, Feng, Senlin Wang, Yuxiang Yuan, Yun Chen, Erhui Guo, and Yuan Li. 2023. "The Addition of a High Concentration of Phosphorus Reduces the Diversity of Arbuscular Mycorrhizal Fungi in Temperate Agroecosystems" Diversity 15, no. 10: 1045. https://doi.org/10.3390/d15101045