

Trichoderma Diversity in Mexico: A Systematic Review and Meta-Analysis

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Searching for Information
2.3. Eligibility Criteria in Published Articles
- Research articles and book chapters in English and Spanish in which the isolation and identification of native strains belonging to the genus Trichoderma within the Mexican territory was mentioned.
- Articles in which the identification of the fungus was conducted at the species level, either morphologically, molecularly, or both.
- Articles that presented a description of the sampling area where the species was recorded by mentioning the locality, geographical coordinates, or other georeferencing parameters.
- Articles that featured the use of commercial species were discarded.
- We also discarded articles that reported repeated strains of species for the same sampling site.
2.4. Eligibility Criteria for Data on the GBIF Platform
- We included records where the fungus strains were identified at the species levels.
- We included records that provided information about the sampling site or the georeferenced location where the strains were isolated.
- Records from the platform that were repeated in any other document found from other databases were discarded.
2.5. Data Extraction and Conditioning Process
2.6. Data Analysis
3. Result
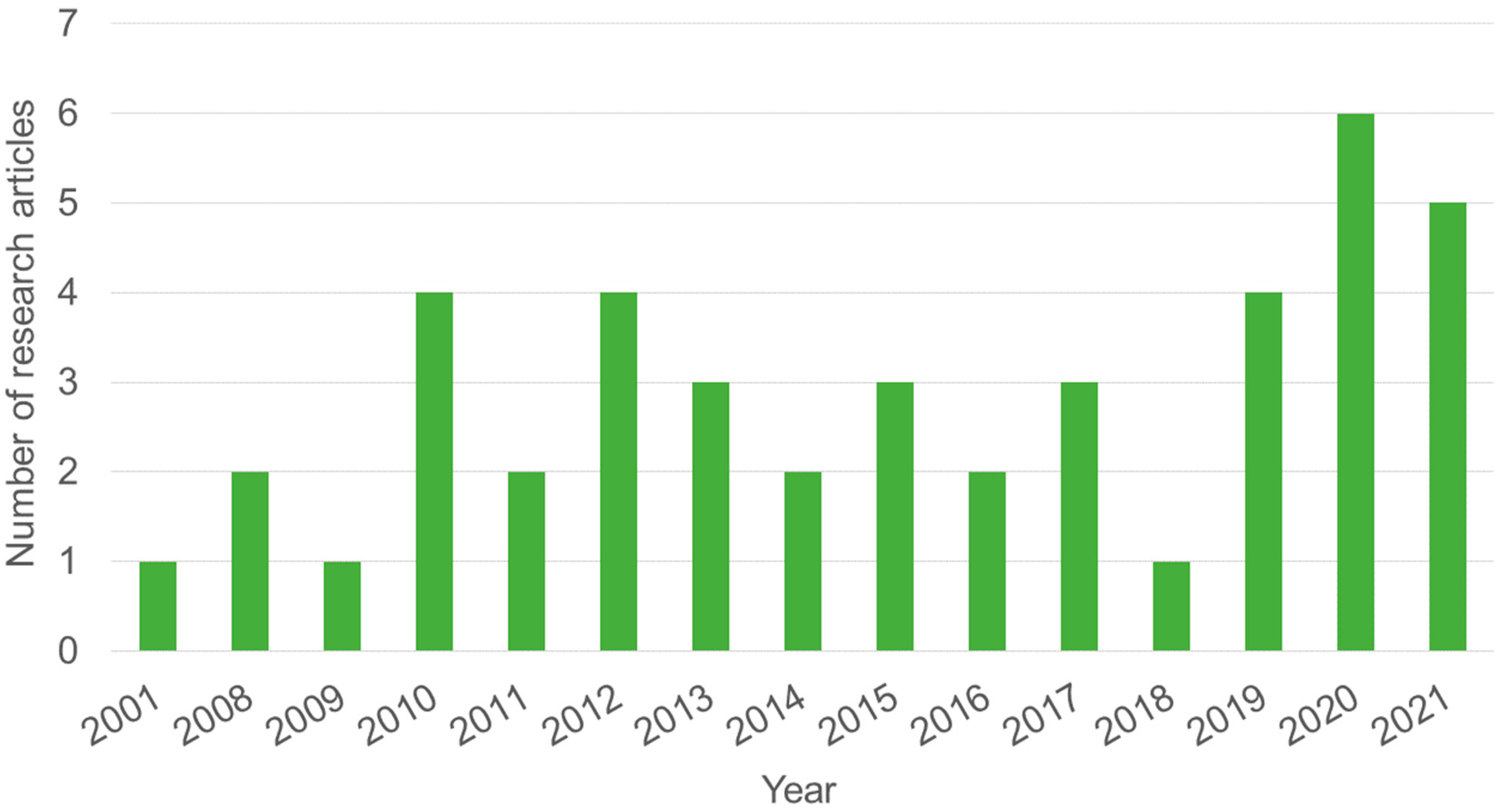
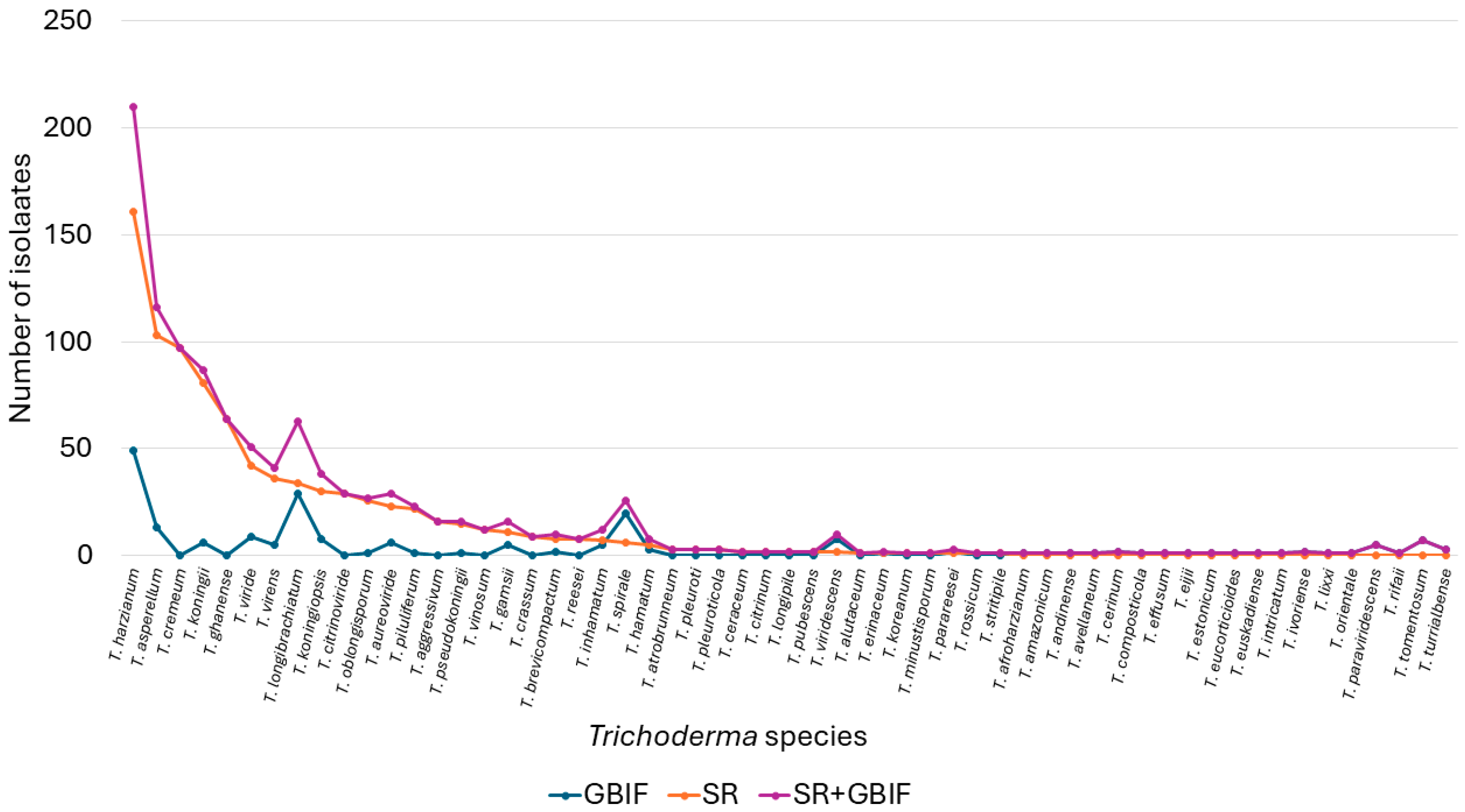
3.1. Qualitative Analysis
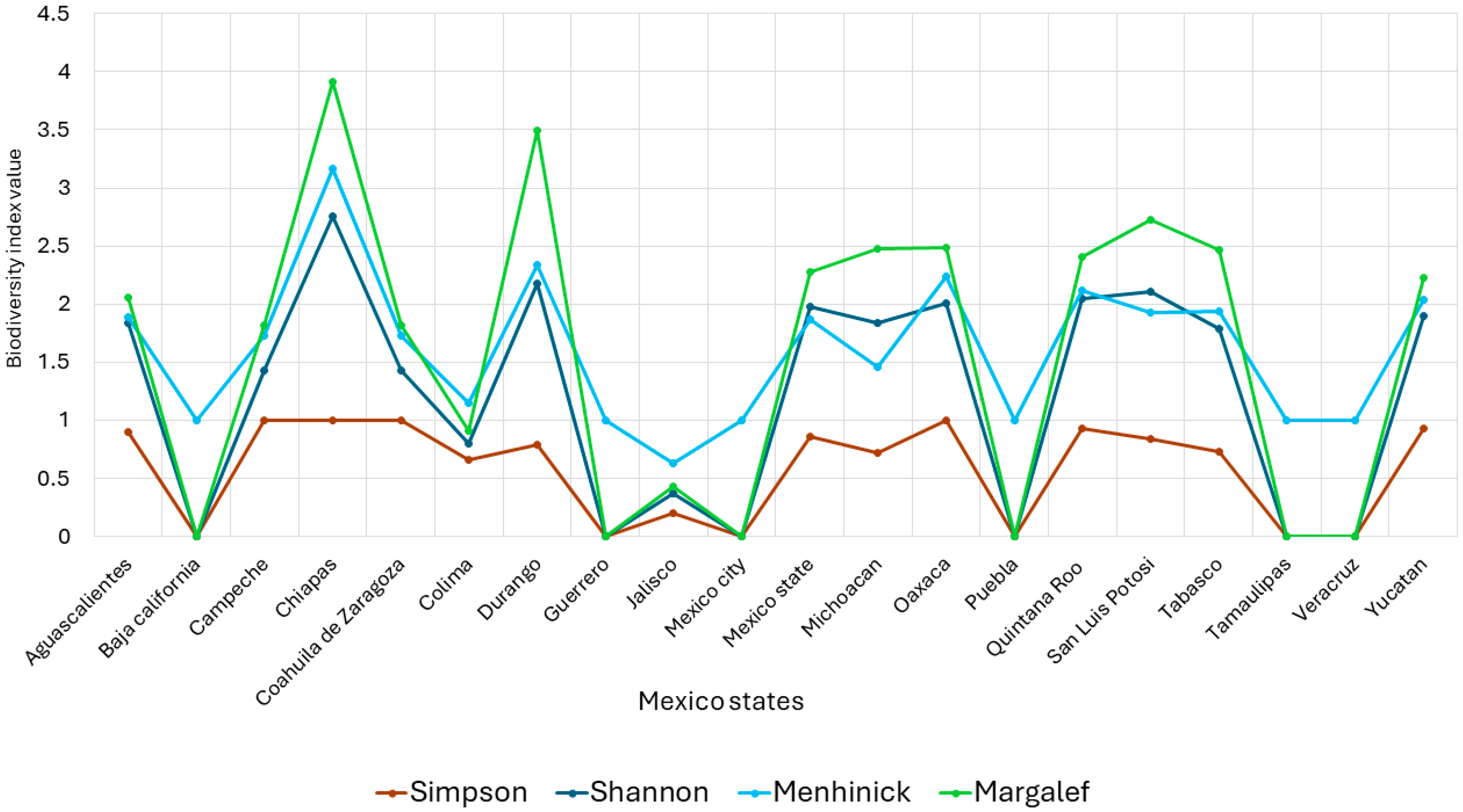
3.2. Quantitative Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cai, F.; Druzhinina, I.S. In Honor of John Bissett: Authoritative Guidelines on Molecular Identification of Trichoderma. Fungal Divers. 2021, 107, 1–69. [Google Scholar] [CrossRef]
- Sharma, S.; Kour, D.; Rana, K.L.; Dhiman, A.; Thakur, S.; Thakur, P.; Thakur, S.; Thakur, N.; Sudheer, S.; Yadav, N.; et al. Chapter 3. Trichoderma: Biodiversity, Ecological Significances, and Industrial Applications. In Recent Advancement in White Biotechnology through Fungi. Fungal Biology; Yavdav, A., Mishra, S., Sing, S., Gupta, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 85–120. [Google Scholar]
- Kubicek, C.P.; Bissett, J.; Druzhinina, I.; Kullnig-Gradinger, C.; Szakacs, G. Genetic and Metabolic Diversity of Trichoderma: A Case Study on South-East Asian Isolates. Fungal Genet. Biol. 2003, 38, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Macías-Rodríguez, L.; Contreras-Cornejo, H.A.; Adame-Garnica, S.G.; del-Val, E.; Larsen, J. The Interactions of Trichoderma at Multiple Trophic Levels: Inter-Kingdom Communication. Microbiol. Res. 2020, 240, 126552. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Sharma, P. Trichoderma: Host Pathogen Interactions and Applications; Springer Nature Singapore Pte Ltd.: Singapore, 2020; Volume 1, p. 319. ISBN 9789811533204. [Google Scholar] [CrossRef]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “Secrets” of a Multitalented Biocontrol Agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Andrade-Hoyos, P.; Silva-Rojas, H.V.; Romero-Arenas, O. Endophytic Trichoderma Species Isolated from Persea americana and Cinnamomum verum Roots Reduce Symptoms Caused by Phytophthora cinnamomi in Avocado. Plants 2020, 9, 1220. [Google Scholar] [CrossRef]
- Quiroz Sarmiento, V.F.; Ferrera Cerrato, R.; Alarcón, A.; María Encarnación, H.L. Antagonismo In Vitro de Cepas de Aspergillus y Trichoderma Hacia Hongos Filamentosos Que Afectan El Cultivo de Ajo. Rev. Mex. Micol. 2008, 26, 27–34. [Google Scholar]
- Osorio Nila, M.A.; Vázquez-García, L.M.; Salgado-Siclán, M.L.; González-Esquivel, C.E. Efecto de Dos Enmiendas Orgánicas y Trichoderma spp. Para Controlar Sclerotinia spp. En Lechuga. Rev. Chapingo Ser. Hortic. 2005, 11, 203–208. [Google Scholar] [CrossRef]
- Michel-Aceves, A.C.; Otero-Sánchez, M.A.; Ariza-Flores, R.; Barros-Ayala, A.; Alarcón-Cruz, N. Eficiencia Biológica de Cepas Nativas de Trichoderma spp., En El Control de Sclerotium rolfsii Sacc., En Cacahuate. Av. Investig. Agropecu. 2013, 17, 89–107. [Google Scholar]
- Tirado-Gallego, P.A.; Lopera-Álvarez, A.; Ríos-Osorio, L.A. Estrategias de Control de Moniliophthora roreri y Moniliophthora perniciosa En Theobroma cacao L.: Revisión Sistemática. Corpoica Cienc. Tecnol. Agropecu. Mosquera 2016, 17, 417–430. [Google Scholar] [CrossRef]
- Canseco Martínez, D.A.; Villegas Aparicio, Y.; Castañeda Hidalgo, E.; Carrillo Rodríguez, J.C.; Robles, C.; Santiago Martínez, G.M. Response of Coffea arabica L. to the application of organic fertilizers and biofertilizers. Rev. Mex. Cienc. Agrícolas 2020, 11, 1285–1298. [Google Scholar]
- Martínez-Salgado, S.J.; Andrade-Hoyos, P.; Romero-Arenas, O.; Villa-Ruano, N.; Landeta-Cortés, G.; Rivera-Tapia, J.A. Control in Vitro de Fusarium sp. Asociado al Cultivo de Cebolla Mediante Trichoderma harzianum. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2021, 39, 2. [Google Scholar] [CrossRef]
- López, G.; González González, C.; Andrés, P. Selección de Cepas Nativas de Trichoderma spp. Con Actividad Antagónica Sobre Phytophthora capsici Leonian y Promotoras de Crecimiento En El Cultivo de Chile (Capsicum annuum L.). Rev. Mex. Fitopatol. 2004, 22, 117–124. [Google Scholar]
- Hernández-Morales, J.; Romero-Rosales, T.; Michel-Aceves, A.C.; Vargas-Hernández, M.; Monteon-Ojeda, A.; Valenzuela-Lagarda, J.L. Antagonismo enzimático de Trichoderma spp., sobre Phytophthora parasitica y Fusarium oxysporum en jamaica (Hibiscus sabdariffa L.). Bioagro 2021, 33, 203–214. [Google Scholar] [CrossRef]
- Castro-del Ángel, E.; Sánchez-Arizpe, A.; Galindo Cepeda, M.E.; Ernesto-Vázquez Badillo, M. Trichoderma Species Effect on the Incidence by Fusarium Verticillioides in Four Maíz Genotypes. Trop. Subtrop. Agroecosyst. 2021, 67, 1–11. [Google Scholar]
- Sánchez-García, B.M.; Espinosa-Huerta, E.; Villordo-Pineda, E.; Rodríguez-Guerra, R.; Mora-Avilés, M.A. Trichoderma spp. Native Strains Molecular Identification and In Vitro Antagonistic Evaluation of Root Phithopathogenic Fungus of the Common Bean (Phaseolus vulgaris L.) Cv. Montcalm. Agrociencia 2017, 51, 63–79. [Google Scholar]
- Ruiz-Cisneros, M.F.; Rios-Velasco, C.; Berlanga-Reyes, D.I.; Ornelas-Paz, J.d.J.; Acosta-Muñiz, C.H.; Romo-Chacón, A.; Zamudio-Flores, P.B.; Pérez-Corral, D.A.; Salas-Marina, M.Á.; Ibarra-Rendón, E.; et al. Incidencia y Agentes Causales de Enfermedades de Raíz y Sus Antagonistas En Manzanos de Chihuahua, México. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2017, 35, 437–462. [Google Scholar] [CrossRef]
- Santos-Villalobos, S.d.l.; Guzmán-Ortiz, D.A.; Gómez-Lim, M.A.; Délano-Frier, J.P.; de-Folter, S.; Sánchez-García, P.; Peña-Cabriales, J.J. Potential Use of Trichoderma asperellum (Samuels, Liechfeldt et Nirenberg) T8a as a Biological Control Agent against Anthracnose in Mango (Mangifera indica L.). Biol. Control 2013, 64, 37–44. [Google Scholar] [CrossRef]
- Samaniego-Gaxiola, J.A.; Pedroza-Sandoval, A.; Chew-Madinaveitia, Y.; Gaytán-Mascorro, A. Desinfestación Reductiva, Suelo Desecado y Trichoderma Harzianum Para Controlar Phymatotrichopsis omnivora En Vivero de Nogal Pecanero. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2019, 37, 2. [Google Scholar] [CrossRef]
- David Maldonado Bonilla, L.; Claudia Sánchez-Espinosa, A.; Luis Villarruel-Ordaz, J. Mycoparasitic Antagonism of a Trichoderma harzianum Strain Isolated from Banana Plants in Oaxaca, Mexico. Biotecnia 2021, 23, 127–134. [Google Scholar] [CrossRef]
- Sánchez, V.; Rebolledo, O. Especies de Trichoderma En Suelos Cultivados Con Agave tequilana En La Región de Los Altos Sur, Jalisco y Valoración de Su Capacidad Antagónica Contra Thielaviopsis paradoxa. Rev. Mex. Micol. 2010, 32, 12–17. [Google Scholar]
- Hernández-Melchor, D.J.; Ferrera-Cerrato, R.; Alarcón, A. Trichoderma: Importancia Agrícola, Biotecnológica, y Sistemas de Fermentacion Para Producir Biomasa y Enzimas de Interés Industrial. Chil. J. Agric. Anim. Sci. Ex. Agro-Cienc. 2019, 35, 98–112. [Google Scholar] [CrossRef]
- Zafra, G.; Absalón, Á.E.; Cuevas, M.D.C.; Cortés-Espinosa, D.V. Isolation and Selection of a Highly Tolerant Microbial Consortium with Potential for PAH Biodegradation from Heavy Crude Oil-Contaminated Soils. Water Air Soil. Pollut. 2014, 225, 1826. [Google Scholar] [CrossRef]
- Halme, P.; Heilmann-Clausen, J.; Rämä, T.; Kosonen, T.; Kunttu, P. Monitoring Fungal Biodiversity—Towards an Integrated Approach. Fungal Ecol. 2012, 5, 750–758. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]
- Devkota, S.; Fang, W.; Arunachalam, K.; Phyo, K.M.M.; Shakya, B. Systematic Review of Fungi, Their Diversity and Role in Ecosystem Services from the Far Eastern Himalayan Landscape (FHL). Heliyon 2023, 9, e12756. [Google Scholar] [PubMed]
- Torres-De La Cruz, M.; Ortiz-García, C.F.; Bautista-Muñoz, C.; Ramírez-Pool, J.A.; Ávalos-Contreras, N.; Cappello-García, S.; De La Cruz-Pérez, A. Diversidad de Trichoderma En El Agroecosistema Cacao Del Estado de Tabasco, México. Rev. Mex. Biodivers. 2015, 86, 947–961. [Google Scholar] [CrossRef]
- Sánchez Hernández, L.; Arias Mota, R.M.; Rosique Gil, J.E.; Pacheco Figueroa, C.J. Diversidad del género Trichoderma (Hypocraceae) en un Área Natural Protegida en Tabasco, México. Acta Bot. Mex. 2018, 123, 167–182. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Michel, A.; Casimiro, A.; Domínguez, R.; Gutiérrez, L.; Moreno, O.; Elena, M.; Escamilla, M.; Carlos, J. Especies de Trichoderma En Suelos Cultivados Con Mango Afectados Por Escoba de Bruja y Su Potencial Inhibitorio Sobre Fusarium Oxysporum y F. Subglutinans. Rev. Mex. Fitopatol. 2001, 19, 154–160. [Google Scholar]
- Talavera-Ortiz, A.; Chaverri, P.; Díaz-Godínez, G.; Acosta-Urdapilleta, M.d.L.; Villegas, E.; Téllez-Téllez, M. Mycelial Inhibition of Trichoderma spp. (Hypocreaceae) Isolated from the Cultivation of Pleurotus Ostreatus (Pleurotaceae) with an Extract of Pycnoporus sp. (Polyporaceae). Acta Bot. Mex. 2020, 127. [Google Scholar] [CrossRef]
- Torres Preciado, C.O.; Rodríguez Alcántar, O.; Herrera-Fonseca, M.d.J.; Figueroa-García, D. Catálogo de La Micobiota Del Complejo Volcánico de Colima, México. Acta Bot. Mex. 2020, 127, e1686. [Google Scholar] [CrossRef]
- Osorio-Hernández, E.; Hernández-Castillo, F.D.; Gallegos-Morales, G.; Rodríguez-Herrera, R.; Castillo-Reyes, F. In-Vitro Behavior of Trichoderma spp. against Phytophthora capsici Leonian. Afr. J. Agric. Res. 2011, 6, 4594–4600. [Google Scholar]
- Vera-Núñez, J.A.; Luna-Martínez, F.; Barcos-Arias, M.S.; Ávila-Miranda, M.E.; Grageda-Cabrera, O.A.; Pena-Cabriales, J.J. Enhancing 15N-Uptake in Maize (Zea mays L.) by Native Trichoderma spp. Strains in Central Mexico. Afr. J. Biotechnol. 2019, 18, 478–488. [Google Scholar] [CrossRef]
- Ibarra Villarreal, A.L.; Parra Cota, F.I.; Yepez, E.A.; Gutiérrez Coronado, M.A.; Valdez Torres, L.C.; De los Santos Villalobos, S. Impacto Del Cambio En El Manejo Del Cultivo de Trigo de Convencional a Orgánico Sobre Las Comunidades Fúngicas Cultivables Del Suelo En El Valle Del Yaqui, México. Agrociencia 2021, 54, 643–659. [Google Scholar] [CrossRef]
- Larralde-Corona, C.P.; Santiago-Mena, M.R.; Sifuentes-Rincón, A.M.; Rodríguez-Luna, I.C.; Rodríguez-Pérez, M.A.; Shirai, K.; Narváez-Zapata, J.A. Biocontrol Potential and Polyphasic Characterization of Novel Native Trichoderma Strains against Macrophomina phaseolina Isolated from Sorghum and Common Bean. Appl. Microbiol. Biotechnol. 2008, 80, 167. [Google Scholar] [CrossRef] [PubMed]
- Toscano, L.; Montero, G.; Cervantes, L.; Stoytcheva, M.; Gochev, V.; Beltrán, M. Production and Partial Characterization of Extracellular Lipase from Trichoderma harzianum by Solid-State Fermentation. Biotechnol. Biotechnol. Equip. 2013, 27, 3776–3781. [Google Scholar] [CrossRef]
- Raymundo, T.; Soto-Agudelo, R.; Bautista-Hernández, S.; Morales-Campós, A.; Valenzuela, R. Catálogo de Los Ascomicetos Del Bosque Mesófilo de Montaña de Tlanchinol, Hidalgo (México). Bol. Soc. Micol. Madr. 2016, 40, 79–104. [Google Scholar]
- Paramo-Aguilera, L.A.; Ortega-Morales, B.O.; Narváez-Zapata, J.A. Culturable Fungi Associated with Urban Stone Surfaces in Mexico City. Electron. J. Biotechnol. 2012, 15, 4. [Google Scholar] [CrossRef]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Genetic and Metabolic Biodiversity of Trichoderma from Colombia and Adjacent Neotropics Regions. Fungal Genet. Biol. 2009, 46, 615–631. [Google Scholar] [CrossRef]
- Moo-Koh, F.A.; Cristóbal Alejo, J.; Reyes Ramírez, A.; Tun Suárez, J.M.; Gamboa Angulo, M. Identificación Molecular de Aislados de Trichoderma spp. y Su Actividad Promotora En Solanum lycopersicum L. Investig. Cienc. Univ. Autónoma Aguascalientes 2017, 25, 5–11. [Google Scholar] [CrossRef]
- López-Valenzuela, B.E.; Armenta-Bojórquez, A.D.; Hernández-Verdugo, S.; Apodaca-Sánchez, M.A.; Samaniego-Gaxiola, J.A.; Leyva-Madrigal, K.Y.; Valdez-Ortiz, A. Selección In Vitro e Identificación de Trichoderma spp. y Bacillus spp. Nativos Para El Control de Phymatotrichopsis omnivora. Inf. Técnica Económica Agrar. 2015, 111, 310–325. [Google Scholar] [CrossRef]
- Montoya-Gonzalez, A.; Quijano-Vicente, G.; Morales-Maza, A.; Ortiz-Uribe, N.; Hernández-Martínez, R. Isolation of Trichoderma spp. from Desert Soil, Biocontrol Potential Evaluation and Liquid Culture Production of Conidia Using Agricultural Fertilizers. J. Fertil. Pestic. 2016, 7, 1. [Google Scholar] [CrossRef]
- Zúñiga-Silgado, D.; Rivera-Leyva, J.C.; Coleman, J.J.; Sánchez-Reyez, A.; Valencia-Díaz, S.; Serrano, M.; De-Bashan, L.E.; Folch-Mallol, J.L. Soil Type Affects Organic Acid Production and Phosphorus Solubilization Efficiency Mediated by Several Native Fungal Strains from Mexico. Microorganisms 2020, 8, 1337. [Google Scholar] [CrossRef]
- Raymundo, T.; Valenzuela, R.; García-Martínez, Y.; Bravo-Álvarez, M.A.; Ramírez-Martínez, J.C.; Bautista-Hernández, S.; Palacios-Pacheco, M.; Luna-Vega, I. Ascomycetes (Fungi) from the Relic Forest of Fagus grandifolia Subsp. Mexicana in Eastern Mexico. Phytotaxa 2019, 418, 1–41. [Google Scholar] [CrossRef]
- Mendoza, H.J.L.; Sánchez Pérez, I.M.; García Olivares, J.G.; Mayek Pérez, N.; González Prieto, J.M.; Quiroz Velásquez, J.D.C. Caracterización Molecular y Agronómica de Aislados de Trichoderma spp Nativos Del Noreste de México. Rev. Colomb. Biotecnol. 2011, 13, 176–185. [Google Scholar]
- Michel-Aceves, A.C.; Hernández-Morales, J.; Toledo-Aguilar, R.; Sabino López, J.E.; Romero-Rosales, T. Antagonistic Capacity of Native Trichoderma spp. against Phytophthora parasitica and Fusarium oxysporum Isolated from Roselle Crops. Rev. Fitotec. Mex. 2019, 42, 235–241. [Google Scholar]
- Del Olmo-Ruiz, M.; Cifuentes-Blanco, J.; Vidal-Gaona, G.; Rosique-Gil, E. Micromicetos Del Suelo de Una Plantación de Plátano (Musa paradisiaca) En Teapa, Tabasco, México. Rev. Mex. Biodivers. 2010, 81, 97–102. [Google Scholar] [CrossRef]
- Vega-Torres, M.G.; Ruiz-Cisneros, M.F.; Pérez-Corral, D.A.; Berlanga-Reyes, D.I.; Ornelas-Paz, J.D.J.; Rios-Velasco, C.; Cambero-Campos, O.J.; Estrada-Virgen, M.O.; Luna-Esquivel, G.; Denise-Revérchon, F.L. Actividad Antifúngica in Vitro de Microorganismos Antagonistas Contra Fusarium oxysporum de Rizosfera de Árboles de Aguacate En Xalisco, Nayarit, México. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2019, 37, 57–64. [Google Scholar] [CrossRef]
- Rios-Velasco, C.; Caro-Cisneros, J.; Berlanga-Reyes, D.; Ruíz-Cisneros, M.F.; Ornelas-Paz, J.; Salas-Marina, M.; Villalobos-Pérez, E.; Guerrero-Prieto, V. Identification and Antagonistic Activity in Vitro of Bacillus spp. and Trichoderma spp. Isolates against Common Phytopathogenic Fungi. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2015, 34, 1. [Google Scholar] [CrossRef]
- Guigón-lópez, C.; Guerrero-prieto, V.; Investigación, C.D.; Cuauhtémoc, P.C.; Cp, M.; Vargas-albores, F.; Carvajal-millán, E.; Investigación, C.D.; Hermosillo, U.; Hermosillo, A.P.; et al. Identificación Molecular de Cepas Nativas de Trichoderma spp. Su Tasa de Crecimiento In Vitro y Antagonismo Contra Hongos Fitopatogenos. Rev. Mex. Fitopatol. 2010, 28, 87–96. [Google Scholar]
- Guadalupe, H.; Roberto, Á.; Rosa, M.; Jorge, C.; Ernesto, C. Morphological and Molecular Characterization of Native Isolates of Trichoderma and Its Potential Biocontrol against Phytophthora infestans. Rev. Mex. Fitopatol. 2017, 58–79. [Google Scholar] [CrossRef]
- Sánchez López, V.; Martínez Bolaños, L.; Zavala González, E.A.; Ramirez Lepe, M. Nuevos Registros de Trichoderma crassum Para México y Su Variación Morfológica En Diferentes Ecosistemas. Rev. Mex. Micol. 2012, 36, 17–26. [Google Scholar]
- Moreno-Pérez, P.; Gamboa-Angulo, M.; Gabriela, H.; Canto-Canché, B.; Rosado-Vallado, M.; Medina-Baizabal, I.L.; Rapa-Tussell, R. Antagonistic Properties of Micromycetes Isolated from Sinkholes of the Yucatán Peninsula against Fungal Phytopathogens. Rev. Mex. Micol. 2014, 40, 27–36. [Google Scholar]
- Francisco-Francisco, N.; Ortega-Ortiz, H.; Benavides-Mendoza, A.; Ramirez, H.; Fuentes-Lara, L.O.; Robledo-Torres, V. Inmovilizacion de Trichoderma harzianum En Hidrogeles de Quitosano y Su Uso En Tomate (Solanum lycopersicum). Terra Latinoam. 2012, 30, 47–57. [Google Scholar]
- Romero-Rosales, T.; Acuña-Soto, J.; Azuara-Domínguez, A.; Lázaro-Dzul, M.O.; Monteon-Ojeda, A.; Vargas-Madriz, H.; Secundino-Eusebio, Y. Identification and In Vitro Antagonism Evaluation of Native Strains of Trichoderma spp. on Phytopathogenic Fungi Associated with Chalice Spot in Jamaica (Hibiscus sabdariffa L.). Trop. Subtrop. Agroecosyst. 2021, 24, 1. [Google Scholar] [CrossRef]
- García Núñez, H.G.; Romero-Gómez, S.d.J.; González-Esquivel, C.E.; Naca-Bernal, E.G.; Martinez-Campos, A.R. Isolation of Native Strains of Trichoderma spp. from Horticultural Soils of the Valley of Toluca, for Potential Biocontrol of Sclerotinia. Trop. Subtrop. Agroecosyst. 2012, 15, 357–365. [Google Scholar]
- Manson, R.H.; Hernández-Ortiz, V.; Gallina, S. Agroecosistemas Cafetaleros de Veracruz: Biodiversidad, Manejo y Conservación; Instituto Nacional de Ecología: Oaxaca, Mexico, 2008; ISBN 970-709-112-6. [Google Scholar]
- Figueroa-Hernández, E.; Godínez-Montoya, L.; Rocha-Quiroz, J.; García-Núñez, R.M.; Pérez-Soto, F. Investigación En Matemáticas, Economía Ciencias Sociales y Agronomía; ECORFAN-México: Mexico City, Mexico, 2015; ISBN 9786078324774. [Google Scholar]
- Howell, C.R. Mechanisms Employed by Trichoderma Species in the Biological Control of Plant Diseases: The History and Evolution of Current Concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Dou, K.; Gao, J.; Zhang, C.; Yang, H.; Jiang, X.; Li, J.; Li, Y.; Wang, W.; Xian, H.; Li, S.; et al. Trichoderma Biodiversity in Major Ecological Systems of China. J. Microbiol. 2019, 57, 668–675. [Google Scholar] [CrossRef]
- Jaklitsch, W.M. European Species of Hypocrea Part II: Species with Hyaline Ascospores. Fungal Divers. 2011, 48, 1–250. [Google Scholar] [CrossRef]
- Jaklitsch, W.M. European Species of Hypocrea Part I. The Green-Spored Species. Stud. Mycol. 2009, 63, 1–91. [Google Scholar] [CrossRef]
- Sun, R.; Liu, Z.; Fu, K.; Fan, L.; Chen, J. Trichoderma Biodiversity in China. J. Appl. Genet. 2012, 53, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Trapero-Quintana, A.; Reyes-Tur, B.; Araújo, N.C. Esfuerzo de Muestreo Necesario Para Estimar La Riqueza Específica Máxima En Tres Comunidades de Odonata En Cuba Empleando Exuvias. Boletín Soc. Entomológica Aragonesa 2011, 49, 285–290. [Google Scholar]
- Chao, A.; Colwell, R.K.; Lin, C.-W.; Gotelli, N.J. Sufficient Sampling for Asymptotic Minimum Species Richness Estimators. Ecology 2009, 90, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Widden, P.; Abitbol, J.-J. Seasonality of Trichoderma Species in a Spruce-Forest Soil. Mycologia 1980, 72, 775–784. [Google Scholar] [CrossRef]
- de Sousa, L.P.; Guerreiro-Filho, O.; Mondego, J.M.C. The Rhizosphere Microbiomes of Five Species of Coffee Trees. Microbiol. Spectr. 2022, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Mulaw, T.B.; Druzhinina, I.S.; Kubicek, C.P.; Atanasova, L. Novel Endophytic Trichoderma spp. Isolated from Healthy Coffea arabica Roots Are Capable of Controlling Coffee Tracheomycosis. Diversity 2013, 5, 750–766. [Google Scholar] [CrossRef]
- Alfredo, L.; Rodrigo, Y.; Mamani-Huayhua, G.; Alfredo Palao-Iturregui, L.; Rodrigo Borja-Loza, Y.; Leon-Ttacca, B. Biocontrol of Coffee Yellow Rust (Hemileia vastatrix Berk. & Br.) with Trichoderma sp. Endophyte Strains. Cultiv. Trop. 2021, 42, 1. [Google Scholar]
- Guzmán, G. Inventorying the Fungi of Mexico. Biodivers. Conserv. 1998, 7, 369–384. [Google Scholar] [CrossRef]
- CONABIO. Ecosistemas de México Biodiversidad Mexicana. Available online: https://www.biodiversidad.gob.mx/ecosistemas/ecosismex (accessed on 25 December 2023).
- Samuels, G.J. Trichoderma: A Review of Biology and Systematics of the Genus. Mycol. Res. 1996, 100, 923–935. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, J.-L.; Chen, J.; Mao, L.-J.; Feng, X.-X.; Zhang, C.-L.; Lin, F.-C. Trichoderma Biodiversity of Agricultural Fields in East China Reveals a Gradient Distribution of Species. PLoS ONE 2016, 11, e0160613. [Google Scholar] [CrossRef]
- Gupta, V.K.; Schmoll, M.; Herrera-Estrella, A.; Upadhyay, R.S.; Druzhinina, I.; Tuohy, M.G. Biotechnology and Biology of Trichoderma; Elsevier: Amsterdam, The Netherlands, 2014; Volume 1, ISBN 9780444595768. [Google Scholar] [CrossRef]
- Gómez, I.; Chet, I.; Herrera-Estrella, A. Genetic Diversity and Vegetative Compatibility among Trichoderma harzianum Isolates. Mol. Gen. Genet. 1997, 256, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Li, G.; Zhang, Z.; Peng, X. Structures and Biological Activities of Secondary Metabolites from Trichoderma harzianum. Mar. Drugs 2022, 20, 701. [Google Scholar] [CrossRef]
- González-Oreja, J.A.; Lou, J. Midiendo La Diversidad Biológica: Más Allá Del Índice de Shannon. Acta Zoológica Lilloana 2012, 56, 3–14. [Google Scholar]
- Aguirre-Acosta, E.; Ulloa, M.; Aguilar, S.; Cifuentes, J.; Valenzuela, R. Biodiversidad de Hongos En México. Rev. Mex. Biodivers. 2014, 85, 76–81. [Google Scholar] [CrossRef]
- Lodge, D.J. Factors Related to Diversity of Decomposer Fungi in Tropical Forests. Biodivers. Conserv. 1997, 6, 681–688. [Google Scholar] [CrossRef]
- Zhang, C.; Druzhinina, I.S.; Kubicek, C.P.; Xu, T. Trichoderma Biodiversity in China: Evidence for a North to South Distribution of Species in East Asia. FEMS Microbiol. Lett. 2005, 251, 251–257. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Biodiversity of Trichoderma (Hypocreaceae) in Southern Europe and Macaronesia. Stud. Mycol. 2015, 80, 1–87. [Google Scholar] [CrossRef]
- Ma, J.; Tsegaye, E.; Li, M.; Wu, B.; Jiang, X. Biodiversity of Trichoderma from Grassland and Forest Ecosystems in Northern Xinjiang, China. 3 Biotech 2020, 10, 362. [Google Scholar] [CrossRef]
- Manzar, N.; Singh, Y.; Kashyap, A.S.; Sahu, P.K.; Rajawat, M.V.S.; Bhowmik, A.; Sharma, P.K.; Saxena, A.K. Biocontrol Potential of Native Trichoderma spp. against Anthracnose of Great Millet (Sorghum bicolour L.) from Tarai and Hill Regions of India. Biol. Control 2021, 152, 104474. [Google Scholar] [CrossRef]
- Bechem, E.T.; Afanga, Y.A. Morphological and Molecular Identification of Fungi Associated with Corm Rot and Blight Symptoms on Plantain (Musa paradisiaca) in Macro-Propagators. Int. J. Biol. Chem. Sci. 2018, 11, 2793. [Google Scholar] [CrossRef]
- Ahmad, M.L.; Rafiq, M.W.; Sheikh, S.; Sahay, S.; Suliman, D.M. Antagonistic Potentiality of Trichoderma harzianum against Cladosporium spherospermum, Aspergillus niger and Fusarium oxysporum. J. Biol. Agric. Healthc. 2012, 2, 72–76. [Google Scholar]
- Wang, H.; Tang, W.; Mao, Y.; Ma, S.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. Isolation of Trichoderma virens 6PS-2 and Its Effects on Fusarium proliferatum f. Sp. Malus Domestica MR5 Related to Apple Replant Disease (ARD) in China. Hortic. Plant J. 2022. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, R.; Duan, Y.; Jiang, W.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. The Endophytic Strain Trichoderma asperellum 6s-2: An Efficient Biocontrol Agent against Apple Replant Disease in China and a Potential Plant-Growth-Promoting Fungus. J. Fungi 2021, 7, 50. [Google Scholar] [CrossRef]
- Kumar, P.; Misra, A.K.; Modi, D.R.; Gupta, V.K. Biocontrol Potential of Trichoderma Species against Mango Malformation Pathogens. Arch. Phytopathol. Plant Prot. 2012, 45, 1237–1245. [Google Scholar] [CrossRef]
- Bellon, M.R.; Berthaud, J. Traditional Mexican Agricultural Systems and the Potential Impacts of Transgenic Varieties on Maize Diversity. Agric. Hum. Values 2006, 23, 3–14. [Google Scholar] [CrossRef]
- Kostyuchenko, N.I.; Lyakh, V.A. Diversity of Fungi in Rhizoplane, Rhizosphere and Edaphosphere of Sunflower at Different Stages of Its Development. Helia 2018, 41, 117–127. [Google Scholar] [CrossRef]
- Inglis, P.W.; Mello, S.C.M.; Martins, I.; Silva, J.B.T.; Macêdo, K.; Sifuentes, D.N.; Valadares-Inglis, M.C. Trichoderma from Brazilian Garlic and Onion Crop Soils and Description of Two New Species: Trichoderma azevedoi and Trichoderma peberdyi. PLoS ONE 2020, 15, e0228485. [Google Scholar] [CrossRef]
- Pavone Maniscalco, D.; Rivas Cordero, M. Diversity of Trichoderma Spp. on Theobroma cacao L. Fields in Carabobo State, Venezuela, and Its Biocontrol Capacity on Crinipellis pernicosa (Stahel) Singer. Interciencia 2010, 35, 777–783. [Google Scholar]
- Arrazate-Argueta, V.D.C.; Martínez-Bolaños, M.; Ayala-Escobar, V.; Avendaño-Arrazate, C.H.; Hernández-Morales, J. Diversidad Morfológica y Fisiológica de Cepas Nativas de Trichoderma spp. En Suelos Cacaoteros de México. Agro Product. 2019, 12. [Google Scholar] [CrossRef]
- Ayyandurai, M.; Akila, R.; Manonmani, K.; Theradimani, M.; Vellaikumar, S. Phytostimulation and Growth Promotion Activities of Trichoderma spp. on Groundnut (Arachis hypogaea L.) Crop. J. Appl. Nat. Sci. 2021, 13, 1172–1179. [Google Scholar] [CrossRef]
- Nawaz, K.; Shahid, A.A.; Bengyella, L.; Subhani, M.N.; Ali, M.; Anwar, W.; Iftikhar, S.; Ali, S.W. Diversity of Trichoderma Species in Chili Rhizosphere That Promote Vigor and Antagonism against Virulent Phytophthora capsici. Sci. Hortic. 2018, 239, 242–252. [Google Scholar] [CrossRef]
- Saldaña-Mendoza, S.A.; Ascacio-Valdés, J.A.; Palacios-Ponce, A.S.; Contreras-Esquivel, J.C.; Rodríguez-Herrera, R.; Ruiz, H.A.; Martínez-Hernandez, J.L.; Sugathan, S.; Aguilar, C.N. Use of Wastes from the Tea and Coffee Industries for the Production of Cellulases Using Fungi Isolated from the Western Ghats of India. Syst. Microbiol. Biomanufacturing 2021, 1, 33–41. [Google Scholar] [CrossRef]
- Alvarado-Marchena, L.; Rivera-Méndez, W. Molecular Identification of Trichoderma spp. in Garlic and Onion Fields and In Vitro Antagonism Trials on Sclerotium cepivorum. Rev. Bras. Cienc. Solo 2016, 40, e0150454. [Google Scholar] [CrossRef]
- Hermosa, M.R.; Keck, E.; Chamorro, I.; Rubio, B.; Sanz, L.; Vizcaíno, J.A.; Grondona, I.; Monte, E. Genetic Diversity Shown in Trichoderma Biocontrol Isolates. Mycol. Res. 2004, 108, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.L.; Rai, P.; Srivastava, A.K.; Kumar, S. Trichoderma for Climate Resilient Agriculture. World J. Microbiol. Biotechnol. 2017, 33, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bubb, P.; May, I.; Miles, L.; Sayer, J. Cloud Forest Agenda; UNEP-WCMC: Cambridge, UK, 2004; Available online: http://www.unep-wcmc.org/resources/publications/UNEP_WCMC_bio_series/20.htm (accessed on 25 December 2023).
- Del Olmo-Ruiz, M.; García-Sandoval, R.; Alcántara-Ayala, O.; Véliz, M.; Luna-Vega, I. Current Knowledge of Fungi from Neotropical Montane Cloud Forests: Distributional Patterns and Composition. Biodivers. Conserv. 2017, 26, 1919–1942. [Google Scholar] [CrossRef]
- Velez, P.; Tapia-Torres, Y.; García-Oliva, F.; Gasca-Pineda, J. Small-Scale Variation in a Pristine Montane Cloud Forest: Evidence on High Soil Fungal Diversity and Biogeochemical Heterogeneity. PeerJ 2021, 9, e11956. [Google Scholar] [CrossRef]
- Savín-Molina, J.; Hernández-Montiel, L.G.; Ceiro-Catasú, W.; Ávila-Quezada, G.D.; Palacios-Espinosa, A.; Ruiz-Espinoza, F.H.; Romero-Bastidas, M. Morphological Characterization and Biocontrol Potential of Trichoderma Species Isolated from Semi-Arid Soils. Rev. Mex. Fitopatol. Mex. J. Phytopathol. 2021, 39. [Google Scholar] [CrossRef]
- Błaszczyk, L.; Strakowska, J.; Chełkowski, J.; Gąbka-Buszek, A.; Kaczmarek, J. Trichoderma Species Occurring on Wood with Decay Symptoms in Mountain Forests in Central Europe: Genetic and Enzymatic Characterization. J. Appl. Genet. 2016, 57, 397–407. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Journal | Articles |
|---|---|
| Acta Botánica Mexicana | [29,32,33] |
| African Journal of Agricultural Research | [34] |
| African Journal of Biotechnology | [35] |
| Agrociencia | [17,36] |
| Applied Microbiology and Biotechnology | [37] |
| Avances en Investigación Agropecuaria | [10] |
| Bioagro | [15] |
| Biological control | [19] |
| Biotechnology & Biotechnological Equipment | [38] |
| Revista de Ciencias Biológicas y de la Salud | [21] |
| Boletin de la Sociedad Micológica de Madrid | [39] |
| Electronic Journal of Biotechnology | [40] |
| Fungal Genetics and Biology | [41] |
| Investigación y Ciencia de la Universidad Autónoma de Aguascalientes | [42] |
| información Técnica Económica Agraria | [43] |
| Journal of Fertilizers and Pesticides | [44] |
| Microorganisms | [45] |
| Phytotaxa | [46] |
| Plants | [7] |
| Revista Colombiana de Biotecnología | [47] |
| Revista Fitotecnia Mexicana | [48] |
| Revista Mexicana de Biodiversidad | [28,49] |
| Revista Mexicana de Fitopatología | [18,31,50,51,52,53] |
| Revista Mexicana de Micología | [8,22,54,55] |
| Terra Latinoamericana | [56] |
| Tropical and Subtropical Agroecosystems | [57,58] |
| Water, Air Soil Pollution | [24] |
| Book: Agroecosistemas cafetaleros de Veracruz: biodiversidad, manejo y conservación | [59] |
| Book: Investigación en Matemáticas, Economía, Ciencias sociales y Agronomía | [60] |
| States | Systematic Review | GBIF | |||
|---|---|---|---|---|---|
| Number of Articles | Number of Isolates | S | Number of Isolates | S | |
| Aguascalientes | - | - | - | 7 | 5 |
| Baja California | 1 | 1 | 1 | 1 | 1 |
| Baja California Sur | 1 | 6 | 6 | - | - |
| Campeche | - | - | - | 3 | 3 |
| Mexico City | 1 | 3 | 3 | 1 | 1 |
| Chiapas | 1 | 4 | 2 | 10 | 10 |
| Chihuahua de Zaragoza | 3 | 26 | 5 | 3 | 3 |
| Coahuila | 2 | 32 | 4 | - | - |
| Colima | 2 | 9 | 3 | 3 | 2 |
| Durango | - | - | - | 31 | 13 |
| Mexico state | 1 | 9 | 2 | 14 | 7 |
| Guanajuato | 3 | 46 | 5 | - | - |
| Guerrero | 7 | 36 | 7 | 1 | 1 |
| Hidalgo | 3 | 5 | 3 | - | - |
| Jalisco | 4 | 13 | 7 | 10 | 2 |
| Michoacan | 2 | 8 | 4 | 57 | 11 |
| Morelos | 2 | 10 | 5 | - | - |
| Nayarit | 1 | 4 | 3 | - | - |
| Oaxaca | 3 | 17 | 3 | 5 | 5 |
| Puebla | 2 | 2 | 2 | 1 | 1 |
| Quintana Roo | 1 | 9 | 2 | 8 | 6 |
| San Luis Potosi | - | - | - | 27 | 10 |
| Sinaloa | 1 | 3 | 3 | - | - |
| Sonora | 2 | 5 | 2 | - | - |
| Tabasco | 4 | 318 | 17 | 17 | 8 |
| Tamaulipas | 2 | 12 | 7 | 1 | 1 |
| Tlaxcala | 2 | 2 | 1 | - | - |
| Veracruz | 3 | 288 | 14 | 1 | 1 |
| Yucatan | 2 | 5 | 3 | 6 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahedo-Quero, H.O.; Aquino-Bolaños, T.; Ortiz-Hernández, Y.D.; García-Sánchez, E. Trichoderma Diversity in Mexico: A Systematic Review and Meta-Analysis. Diversity 2024, 16, 68. https://doi.org/10.3390/d16010068
Ahedo-Quero HO, Aquino-Bolaños T, Ortiz-Hernández YD, García-Sánchez E. Trichoderma Diversity in Mexico: A Systematic Review and Meta-Analysis. Diversity. 2024; 16(1):68. https://doi.org/10.3390/d16010068
Chicago/Turabian StyleAhedo-Quero, Hector Osvaldo, Teodulfo Aquino-Bolaños, Yolanda Donají Ortiz-Hernández, and Edgar García-Sánchez. 2024. "Trichoderma Diversity in Mexico: A Systematic Review and Meta-Analysis" Diversity 16, no. 1: 68. https://doi.org/10.3390/d16010068