
Mind the Gaps: Taxonomic, Geographic and Temporal Data of Marine Invertebrate Databases from Mozambique and São Tomé and Príncipe

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Compilation of Data
2.3. Taxonomic Coverage
2.4. Geographical and Temporal Coverage
3. Results
3.1. Compilation of Data
3.2. Taxonomic Coverage
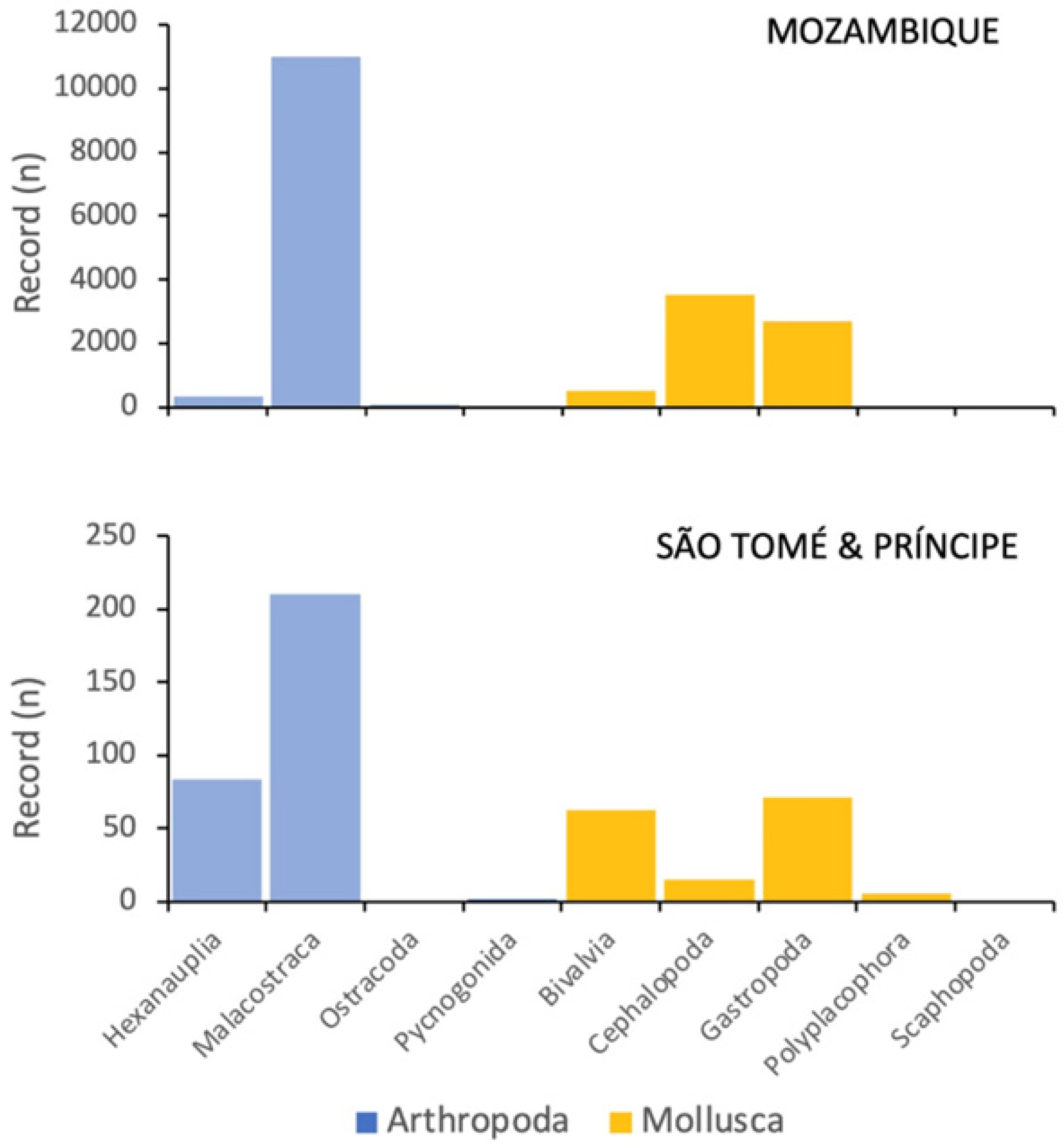
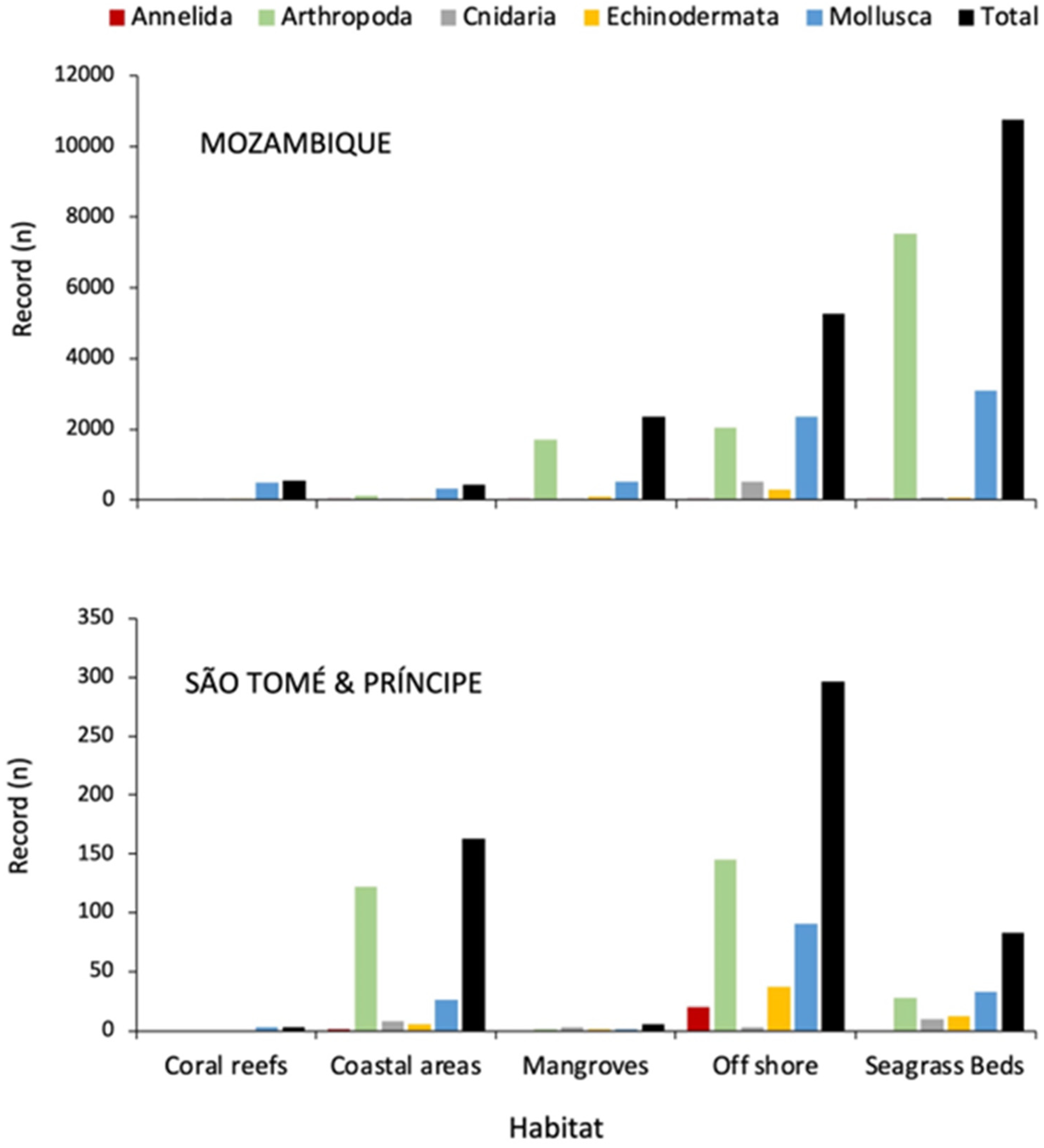
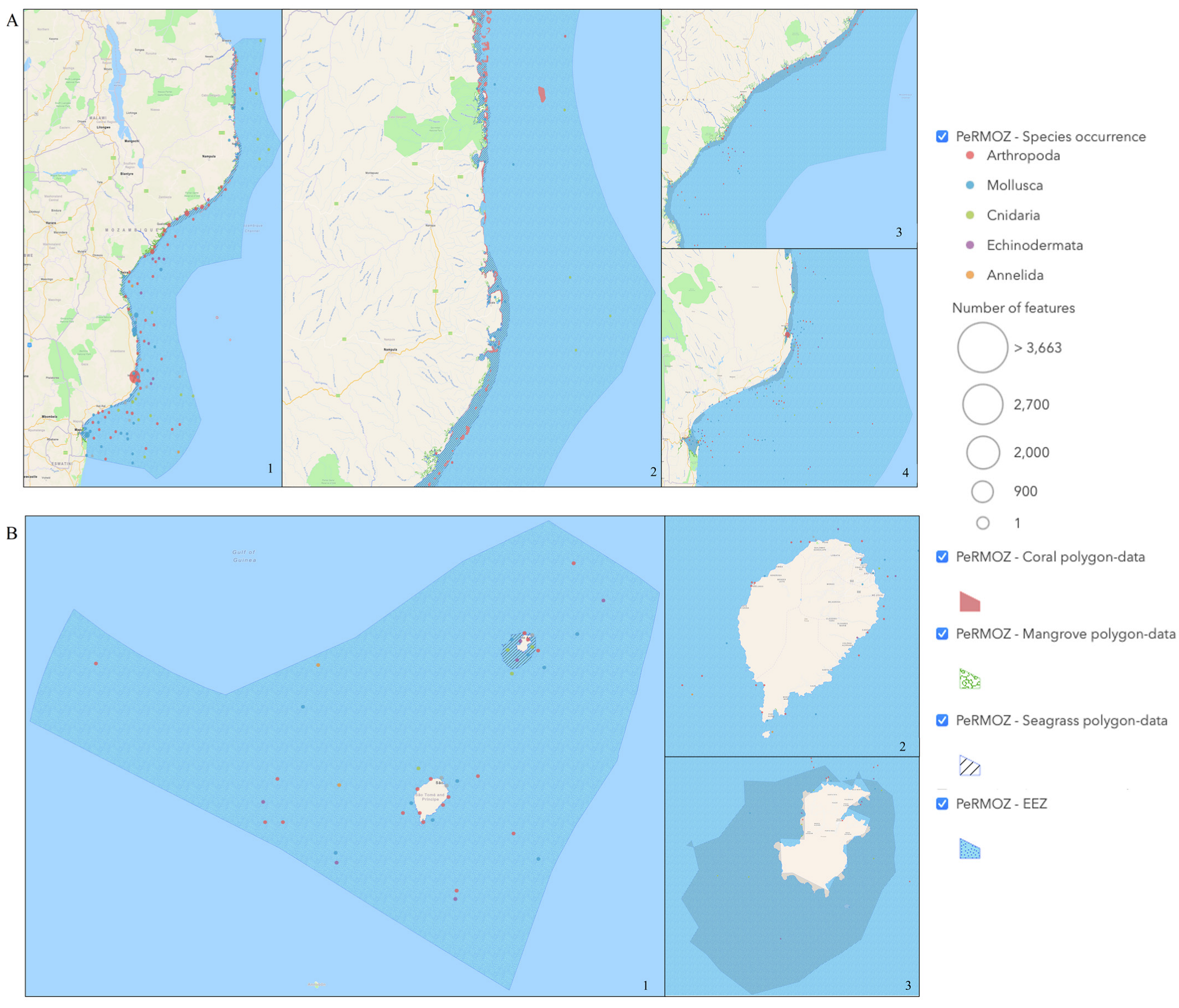
3.3. Geographical Coverage
3.4. Temporal Coverage
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix A.1. Mozambique Occurrence Data
- Acanthocephala: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.a9e0wq, accessed on 25 October 2018
- Annelida: GBIF.org Occurrence Download https://doi.org/10.15468/dl.cgskgn, accessed on 25 October 2018
- Arthropoda: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.rsvrwu, accessed on 25 October 2018
- Decapoda MUHNAC: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.wtl0gh, accessed on 3 December 2018
- Ascidiacea: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.ayjvmg, accessed on 10 October 2018
- Brachiopoda: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.lwdyvc, accessed on 25 October 2018
- Bryozoa: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.bamx4r, accessed on 25 October 2018
- Cnidaria: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.tdo85o, accessed on 25 October 2018
- Echinodermata: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.ob3jcd, accessed on 25 October 2018
- Hemichordata: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.rsyyeo, accessed on 25 October 2018
- Mollusca: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.uzkzcv, accessed on 25 October 2018
- Nematoda: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.guvpnx, accessed on 25 October 2018
- Nematomorpha: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.csf3en, accessed on 25 October 2018
- Nemertea: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.egimov, accessed on 25 October 2018
- Platyhelminthes: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.vlotks, accessed on 25 October 2018
- Porifera: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.kkre6f, accessed on 25 October 2018
- Rotifera: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.nb6gme, accessed on 25 October 2018
- Sipuncula: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.vwd9ve, accessed on 25 October 2018
- Tardigrada: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.zodhhs, accessed on 25 October 2018
- Thaliacea: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.ihck3c, accessed on 10 October 2018
Appendix A.2. São Tomé and Príncipe Occurrence Data
- Annelida: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.p7vvlf, accessed on 10 October 2018
- Arthropoda: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.q4jiv0, accessed on 10 October 2018
- Ascidiacea: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.thevng, accessed on 10 October 2018
- Bryozoa: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.2zowxw, accessed on 10 October 2018
- Cnidaria: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.lrjhwj, accessed on 10 October 2018
- Echinodermata: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.isduid, accessed on 10 October 2018
- Mollusca: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.px6i6k, accessed on 10 October 2018
- Nematoda: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.j8lkto, accessed on 10 October 2018
- Nematomorpha: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.e9gwz5, accessed on 10 October 2018
- Platyhelminthes: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.f107js, accessed on 10 October 2018
- Porifera: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.krzegd, accessed on 10 October 2018
- Sipuncula: GBIF.org GBIF Occurrence Download https://doi.org/10.15468/dl.dlbbow, accessed on 10 October 2018
Appendix A.3. Literature Occurrence Data
- Ahyong, S.T. Cymonomid crabs of the MAINBAZA Expedition (Decapoda: Brachyura). Zootaxa 2014, 3821, 384–390. https://10.11646/zootaxa.3821.3.7
- Baba, K.; Tirmizi, N. A New Chirostylid (Crustacea, Decapoda, Anomura) From Deeper Parts of the Japanese Waters and Off the East Coast of Africa. Proc. Jpn. Soc. Syst. Zool. 1979, 17, 52–57. https://10.19004/pjssz.17.0_52
- Berggren, M. Periclimenes Nomadophila and Tuleariocaris Sarec, Two New Species of Pontoniine Shrimps (Decapoda: Pontoniinae), from Inhaca Island, Mozambique. J. Crustac. Biol. 1994, 14, 782–802.
- Berggren, M. Athanopsis rubricinctuta, New Species (Decapoda: Natantia: Alpheidae), a Shrimp Associated with an Echiuroid at Inhaca Island, Moçambique. J. Crustac. Biol. 1991. https://10.2307/1548555
- Berggren, M. Dasella herdmaniae (Lebour) (Decapoda: Natantia: Pontoniinae) from Moçambique and Establishment of a New Species, Dasella brucei. J. Crustac. Biol. 1990, 10, 554–559.
- Brito, A.; Pena, A. Population structure and recruitment of penaeid shrimps from the Pungué River Estuary to the Sofala Bank Fishery, Mozambique. West. Indian Ocean. J. Mar. Sci. 2007, 147–158.
- Cabezas, P.; Macpherson, E.; Machordom, A. Allogalathea (Decapoda: Galatheidae): a monospecific genus of squat lobster? Zool. J. Linn. Soc. 2011, 162, 245–270. https://10.1111/j.1096-3642.2010.00681.x
- Cabezas, P.; MacPherson, E.; MacHordom, A. Taxonomic revision of the genus Paramunida Baba, 1988 (Crustacea: Decapoda: Galatheidae): A morphological and molecular approach. Zootaxa 2010. https://10.11646/zootaxa.2712.1.1
- Castro, P.; Williams, A.B.; Cooper, L.L. Revision of the family Latreilliidae Stimpson, 1858 (Crustacea, Decapoda, Brachyura). Zoosystema Paris 2003, 25, 601–634.
- Clark, P.F.; Galil, B.S. A revision of the xanthid genus Pilodius Dana, 1851 (Crustacea: Brachyura: Xanthoidea). J. Nat. Hist. 1993. https://10.1080/00222939300770691
- Clark, P.F.; Paula, J. Descriptions of ten xanthoidean (Crustacea: Decapoda: Brachyura) first stage zoeas from Inhaca Island, Mozambique. Raffles Bull. Zool. 2003, 51, 323–378.
- Farfante, I.P.; Ivanov, B.G. Mesopenaeus Mariae, a New Species of Shrimp (Penaeoidea: Solenoceridae), the First Record of the Genus in the Indo-west Pacific. J. Crustac. Biol. 1982, 2, 303–313.
- Flores, A.A.V.; Saraiva, J.; Paula, J. Sexual maturity, reproductive cycles, and juvenile recruitment of Perisesarma guttatum (Brachyura, Sesarmidae) at Ponta Rasa mangrove swamp, Inhaca Island, Mozambique. J. Crustac. Biol. 2002, 22, 143–156. https://10.1651/0278-0372(2002)022[0143:smrcaj]2.0.co;2
- Fratini, S.; Vannini, M.; Cannicci, S.; Schubart, C.D. Tree-climbing mangrove crabs: A case of convergent evolution. Evol. Ecol. Res. 2005, 7, 219–233.
- Freitas, A.J. The Penaeoidea of southeast Africa. III—The Family Penaeidae (excluding Genus Penaeus). Durban 1987.
- Freitas, A.J. The Penaeoidea of southeast Africa. II—The Families Aristeidae and Solenoceridae. Durban 1985.
- Galil, B.S. An examination of the genus Philyra Leach, 1817 (Crustacea, Decapoda, Leucosiidae) with descriptions of seven new genera and six new species. Zoosystema 2009, 31, 279–320. https://10.5252/z2009n2a4
- Galil, B.S. Crustacea Decapoda: review of the genera and species of the family Polychelidae Wood-Mason, 1874, Mémoires du Muséum national d’histoire naturelle. 2000.
- Hayashi, K.-I. Crustacea Decapoda: revision of Pasiphaea sivado (Risso, 1816) and related species, with descriptions of one new genus and five new species (Pasiphaeidae), Mémoires du Muséum national d’Histoire naturelle. 1999.
- Holthuis, L.B. The Indo-Pacific scyllarine lobsters (Crustacea, Decapoda, Scyllaridae). Zoosystema 2002, 24, 499–683.
- Komai, T.; Chan, T.-Y. Two new pandalid shrimps and the discovery of the second specimen of the rare hippolytid shrimp Leontocaris bulga Taylor & Poore, 1998 (Crustacea, Decapoda) from the Mozambique MAINBAZA Cruise. Zoosystema 2010, 32, 625–641. https://10.5252/z2010n4a6
- Komai, T.; Wada, K. A revision of the estuarine crab genus Ilyograpsus Barnard, 1955 (Crustacea: Decapoda: Brachyura: Macrophthalmidae), with descriptions of a new genus and one new species. Raffles Bull. Zool. 2008, 56, 357–384.
- Lai, J.C.Y.; Ng, P.K.L.; Davie, P.J.F. A revision of the Portunus pelagicus (Linnaeus, 1758) species complex (Crustacea: Brachyura: Portunidae), with the recognition of four species. Raffles Bull. Zool. 2010, 58, 199–237.
- Lemaitre, R. A worldwide review of hermit crab species of the genus Sympagurus Smith, 1883 (Crustacea: Decapoda: Parapaguridae), Tropical Deep-Sea Benthos. 2004.
- Lemaitre, R.; Rahayu, D.L.; Komai, T. A revision of “blanket-hermit crabs” of the genus paguropsis henderson, 1888, with the description of a new genus and five new species (Crustacea, anomura, diogenidae). Zookeys 2018, 2018, 17–97. https://10.3897/zookeys.752.23712
- Litulo, C. Population and reproductive biology of the fiddler crab Uca chlorophthalmus (Brachyura: Ocypodidae) from Inhaca Island, southern Mozambique. J. Mar. Biol. Assoc. United Kingd. 2006, 86, 737–742. https://10.1017/S0025315406013646
- Litulo, C.; Abreu, D.; Tudge. Reproductive biology of Calcinus laevimanus (Randall, 1840) (Decapoda: Anomura: Diogenidae) from Inhaca Island, southern Mozambique. Biol. Anomura II 2006, 6, 57–66.
- Litulo, C.; Macia, A.; Mantelatto, F.L.M. Fecundity and sexual maturity of the crab Macrophthalmus depressus (Brachyura: Ocypodidae) from Inhaca Island, Mozambique. Afr. J. Aquat. Sci. 2005, 30, 179–183. https://10.2989/16085910509503854
- Litulo, C.; Tudge, C. Population structure and breeding season of the hermit crab Diogenes brevirostris Stimpson, 1858 (Decapoda, Anomura, Diogenidae) from southern Mozambique. J. Nat. Hist. 2005, 39, 2887–2899. https://10.1080/00222930500206107
- Macia, A.; Quincardete, I.; Paula, J. A comparison of alternative methods for estimating population density of the fiddler crab Uca annulipes at Saco Mangrove, Inhaca Island (Mozambique). Hydrobiologia 2001, 449, 213–219. https://10.1023/A:1017502817087
- Macpherson, E.; Rodríguez-Flores, P.C.; Machordom, A. New sibling species and new occurrences of squat lobsters (Crustacea, Decapoda) from the western Indian ocean. Eur. J. Taxon. 2007, 2017, 1–61. https://10.5852/ejt.2017.343
- Mkare, T.K.; Groeneveld, J.C.; Teske, P.R.; Matthee, C.A. Comparative genetic structure in two high-dispersal prawn species from the south-west Indian Ocean. Afr. J. Mar. Sci. 2017, 39, 467–474. https://10.2989/1814232X.2017.1402089
- Penha-Lopes, G.; Torres, P.; Macia, A.; Paula, J. Population structure, fecundity and embryo loss of the sea grass shrimp Latreutes pymoeus (Decapoda: Hippolytidae) at Inhaca Island, Mozambique. J. Mar. Biol. Assoc. United Kingd. 2007, 87, 879–884. https://10.1017/S0025315407056767
- Saito, T.; Komai, T. A review of species of the genera Spongicola de Haan, 1844 and Paraspongicola de Saint Laurent & Cleva, 1981 (Crustacea, Decapoda, Stenopodidea, Spongicolidae). Zoosystema 2008, 30, 87–147.
- Sakai, K. Upogebiidae of the World (Decapoda, Thalassinidea). Crustac. Monogr. 2006, 6, 186.
- Sakai, K. Dr. R. Plante’s collection of the families Callianassidae and Gourretiidae (Decapoda, Thalassinidea) from Madagascar, with the description of two new genera and one new species of the Gourretiidae sakai, 1999 (new status) and two new species of the Callia. Crustaceana 2004, 77, 553–601. https://10.1163/1568540041718019
- Serène, R. Crustacés Décapodes Brachyoures de l’océan Indien Occidental et de la Mer Rouge, Xanthoidea: Xanthidae et Trapeziidae. Faune Trop. 1984.
- Simbine, L.; Marques, C.G.; Freitas, P.D.; Samucidine, K.E.; Gusmão, J.; Tavares, C.; Junior, P.G. Metapenaeus dobsoni (Miers, 1878), an alien Penaeidae in Mozambican coastal waters-confirmation by mtDNA and morphology analyses. West. Indian Ocean J. Mar. Sci. 2018, 17, 1–12.
- Spiridonov, V.A.; Apel, M. A new species and new records of deep-water Calappidae (Crustacea: Decapoda) from the Indian Ocean with a key to the Mursia Desmarest, 1823 species of the region. J. Nat. Hist. 2007, 41, 2851–2890. https://10.1080/00222930701770786
- Torres, P.; Penha-lopes, G.; Macia, A.; Paula, J. Population structure and egg production of the seagrass shrimp hippolyte kraussiana stimpson, 1860 (decapoda: Hippolytidae) at inhaca island, mozambique. Invertebr. Reprod. Dev. 2007, 50, 145–153. https://10.1080/07924259.2007.9652239
- Torres, P.; Penha-Lopes, G.; Narciso, L.; Macia, A.; Paula, J. Fecundity and brood loss in four species of fiddler crabs, genus Uca (Brachyura: Ocypodidae), in the mangroves of Inhaca Island, Mozambique. J. Mar. Biol. Assoc. United Kingd. 2009, 89, 371–378. https://10.1017/S0025315408002701
- Williams, A.B. Revision of the genus Latreilia Roux. Quad. del Lab. di Tecnol. della Pesca 1982, 3, 227–255.
References
- Smith, V.S.; Blagoderov, V. Bringing Collections out of the Dark. Zookeys 2012, 209, 1–6. [Google Scholar] [CrossRef]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial Bias in the GBIF Database and Its Effect on Modeling Species’ Geographic Distributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- Beck, J.; Ballesteros-Mejia, L.; Buchmann, C.M.; Dengler, J.; Fritz, S.A.; Gruber, B.; Hof, C.; Jansen, F.; Knapp, S.; Kreft, H.; et al. What’s on the Horizon for Macroecology? Ecography Cop. 2012, 35, 673–683. [Google Scholar] [CrossRef]
- Jetz, W.; McPherson, J.M.; Guralnick, R.P. Integrating Biodiversity Distribution Knowledge: Toward a Global Map of Life. Trends Ecol. Evol. 2012, 27, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.D. Principles and Methods of Data Cleaning: Primary Species and Species-Occurrence Data; Version 1; GBIF: Copenhagen, Denmark, 2005. [Google Scholar]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating Species Diversity and Distribution in the Era of Big Data: To What Extent Can We Trust Public Databases? Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandepitte, L.; Bosch, S.; Tyberghein, L.; Waumans, F.; Vanhoorne, B.; Hernandez, F.; De Clerck, O.; Mees, J. Fishing for Data and Sorting the Catch: Assessing the Data Quality, Completeness and Fitness for Use of Data in Marine Biogeographic Databases. Database 2015, 2015, bau125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, D.; Collen, B.; Ariño, A.; Patricia Koleff, P.K.; Guinotte, J.; Kerr, J.; Chavan, V. Bridging the Biodiversity Data Gaps: Recommendations to Meet Users’ Data Needs. Biodivers. Inform. 2013, 8, 41–58. [Google Scholar] [CrossRef]
- Chapman, A.D. Principles of Data Quality; GBIF: Copenhagen, Denmark, 2005. [Google Scholar]
- Costello, M.J.; Michener, W.K.; Gahegan, M.; Zhang, Z.Q.; Bourne, P.E. Biodiversity Data Should Be Published, Cited, and Peer Reviewed. Trends Ecol. Evol. 2013, 28, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.D. Uses of Primay Species-Occurrence Data; GBIF: Copenhagen, Denmark, 2005. [Google Scholar]
- Garcia, D.A.; Amori, M.; Giovanardi, F.; Piras, G.; Groppi, D.; Cumo, F.; de Santoli, L. An Identification and a Prioritisation of Geographic and Temporal Data Gaps of Mediterranean Marine Databases. Sci. Total Environ. 2019, 668, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Lam-Gordillo, O.; Baring, R.; Dittmann, S. Ecosystem Functioning and Functional Approaches on Marine Macrobenthic Fauna: A Research Synthesis towards a Global Consensus. Ecol. Indic. 2020, 115, 106379. [Google Scholar] [CrossRef]
- Soberón, J.; Peterson, A.T. Monitoring Biodiversity Loss with Primary Species-Occurrence Data: Toward National-Level Indicators for the 2010 Target of the Convention on Biological Diversity. AMBIO A J. Hum. Environ. 2009, 38, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Samoilys, M.; Pabari, M.; Andrew, T.; Maina, G.W.; Church, J.; Momanyi, A.; Mibei, B.; Monjane, M.; Shah, A.; Menomussanga, M.; et al. Resilience of Coastal Systems and Their Human Partners in the Western Indian Ocean; Fitzgerald, M.A., Ed.; IUCN ESARO, WIOMSA, CORDIO and the UNEP Naiorbi Convention: Nairobi, Kenya, 2015; ISBN 978-2-8317-1724-1. [Google Scholar]
- Alexandre, A.; Silva, J.; Ferreira, R.; Paulo, D.; Serrão, E.A.; Santos, R. First Description of Seagrass Distribution and Abundance in São Tomé and Príncipe. Aquat. Bot. 2017, 142, 48–52. [Google Scholar] [CrossRef]
- Ministério dos Recursos Naturais e Meio Ambiente Primeiro Relatório Nacional Da Biodiversidade. 2003. Available online: https://www.terrabrasilis.org.br/ecotecadigital/pdf/serie-biodiversidade--03-legislacao-ambiental-brasileira-grau-de-adequacao-a-convencao-sobre-diversidade-biologica.pdf (accessed on 25 November 2018).
- Haroun, R.; Herrero Barrencua, A.; Abreu, A.D. Mangrove Habitats in São Tomé and Príncipe (Gulf of Guinea, Africa): Conservation and Management Status. Coast. Res. Libr. 2018, 25, 589–605. [Google Scholar] [CrossRef]
- Wieczorek, J.; Bloom, D.; Guralnick, R.; Blum, S.; Döring, M.; Giovanni, R.; Robertson, T.; Vieglais, D. Darwin Core: An Evolving Community-Developed Biodiversity Data Standard. PLoS ONE 2012, 7, e29715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niza, H.; Bento, M.; Lopes, L.F.; Cartaxana, A.; Correia, A.M. A Picture Is Worth a Thousand Words: Using Digital Tools to Visualise Marine Invertebrate Diversity Data along the Coasts of Mozambique and Sao Tomé & Príncipe. Biodivers. Data J. 2021, 9, e68817. [Google Scholar] [CrossRef] [PubMed]
- Bento, M.; Niza, H.; Cartaxana, A.; Bandeira, S.; Paula, J.; Correia, A.M. A Dataset of Marine Macroinvertebrate Diversity from Mozambique and São Tomé and Príncipe. Available online: https://www.gbif.org/dataset/3e0e4ec9-1905-4cae-9691-c0fa79361ac3 (accessed on 19 October 2022).
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Arie, B.; Blomberg, N.; Boiten, J.-W.; Bonino da Silva Santos, L.; Bourne, P.E.; et al. Comment: The FAIR Guiding Principles for Scientific Data Management and Stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paula, J.; Bandeira, S. An Introduction to the Maputo Bay. In The Maputo Bay Ecosystem; Paula, J., Bandeira, S., Eds.; WIOMSA: Zanzibar Town, Tanzania, 2014; pp. 3–10. ISBN 978-9987-9559-3-0. [Google Scholar]
- Pola, M.; Gosliner, T.M. The First Molecular Phylogeny of Cladobranchian Opisthobranchs (Mollusca, Gastropoda, Nudibranchia). Mol. Phylogenet. Evol. 2010, 56, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Mora, C.; Tittensor, D.P.; Myers, R.A. The Completeness of Taxonomic Inventories for Describing the Global Diversity and Distribution of Marine Fishes. Proc. R. Soc. B Biol. Sci. 2008, 275, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, I.Q.; da Luz Mathias, M.; Bastos-Silveira, C. Mapping Knowledge Gaps of Mozambique’s Terrestrial Mammals. Sci. Rep. 2019, 9, 18184. [Google Scholar] [CrossRef] [PubMed]
- Howarth, W. The Marine Strategy Framework Directive. J. Water Law 2008, 19, 95–97. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | MOZ | STP | MOZ | STP |
|---|---|---|---|---|
| Annelida | 45 | 21 | 0.23% | 3.81% |
| Arthropoda | 11,411 | 296 | 58.94% | 53.72% |
| Cnidaria | 649 | 24 | 3.36% | 4.36% |
| Echinodermata | 469 | 56 | 2.42% | 10.16% |
| Mollusca | 6785 | 154 | 35.04% | 27.95% |
| Total | 19,359 | 551 | 100.00% | 100.00% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bento, M.; Niza, H.; Cartaxana, A.; Bandeira, S.; Paula, J.; Correia, A.M. Mind the Gaps: Taxonomic, Geographic and Temporal Data of Marine Invertebrate Databases from Mozambique and São Tomé and Príncipe. Diversity 2023, 15, 70. https://doi.org/10.3390/d15010070
Bento M, Niza H, Cartaxana A, Bandeira S, Paula J, Correia AM. Mind the Gaps: Taxonomic, Geographic and Temporal Data of Marine Invertebrate Databases from Mozambique and São Tomé and Príncipe. Diversity. 2023; 15(1):70. https://doi.org/10.3390/d15010070
Chicago/Turabian StyleBento, Marta, Henrique Niza, Alexandra Cartaxana, Salomão Bandeira, José Paula, and Alexandra Marçal Correia. 2023. "Mind the Gaps: Taxonomic, Geographic and Temporal Data of Marine Invertebrate Databases from Mozambique and São Tomé and Príncipe" Diversity 15, no. 1: 70. https://doi.org/10.3390/d15010070