

Haemosporidians in Non-Passerine Birds of Colombia: An Overview of the Last 20 Years of Research
 , ,
, ,
Abstract
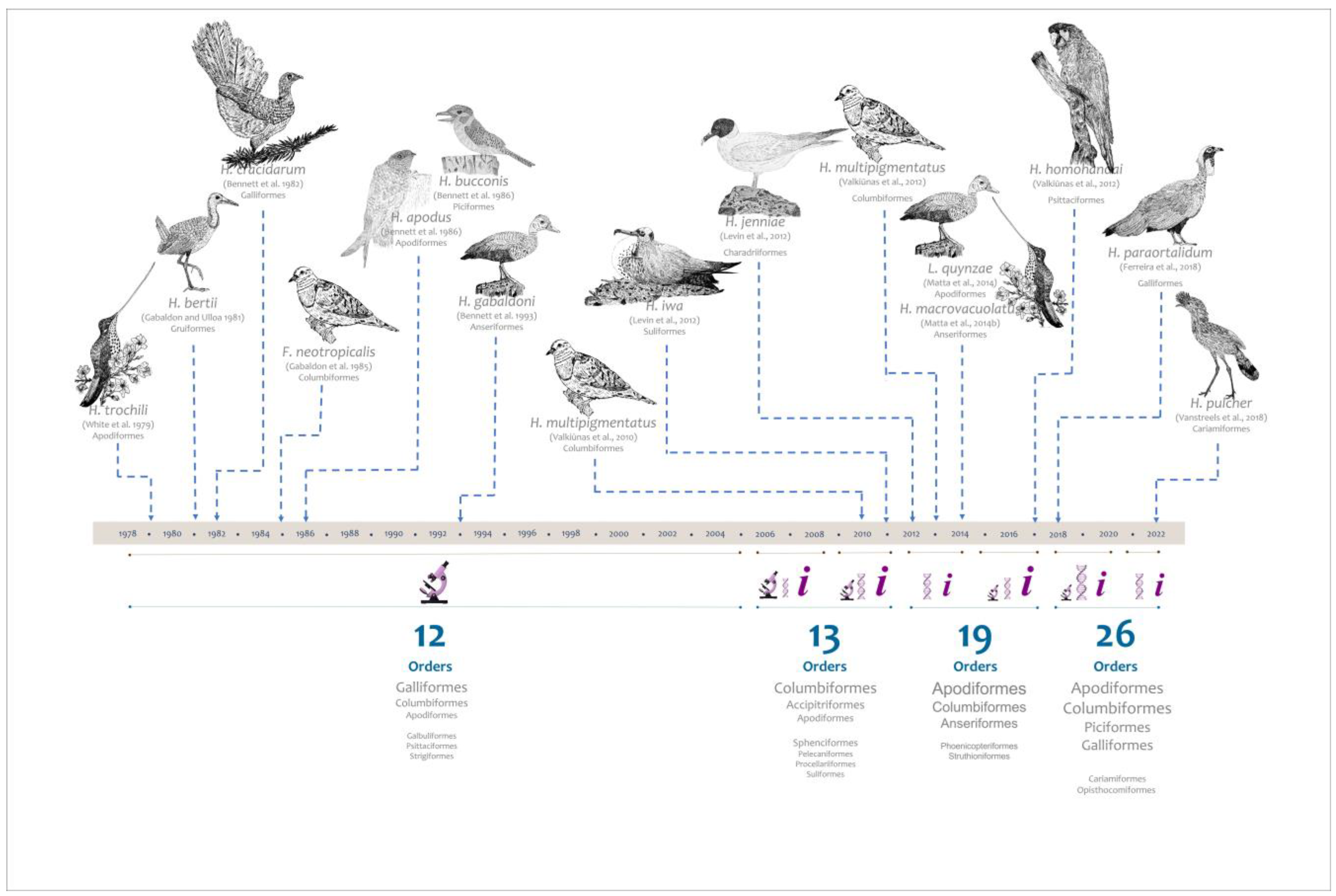
:1. Introduction
2. Materials and Methods
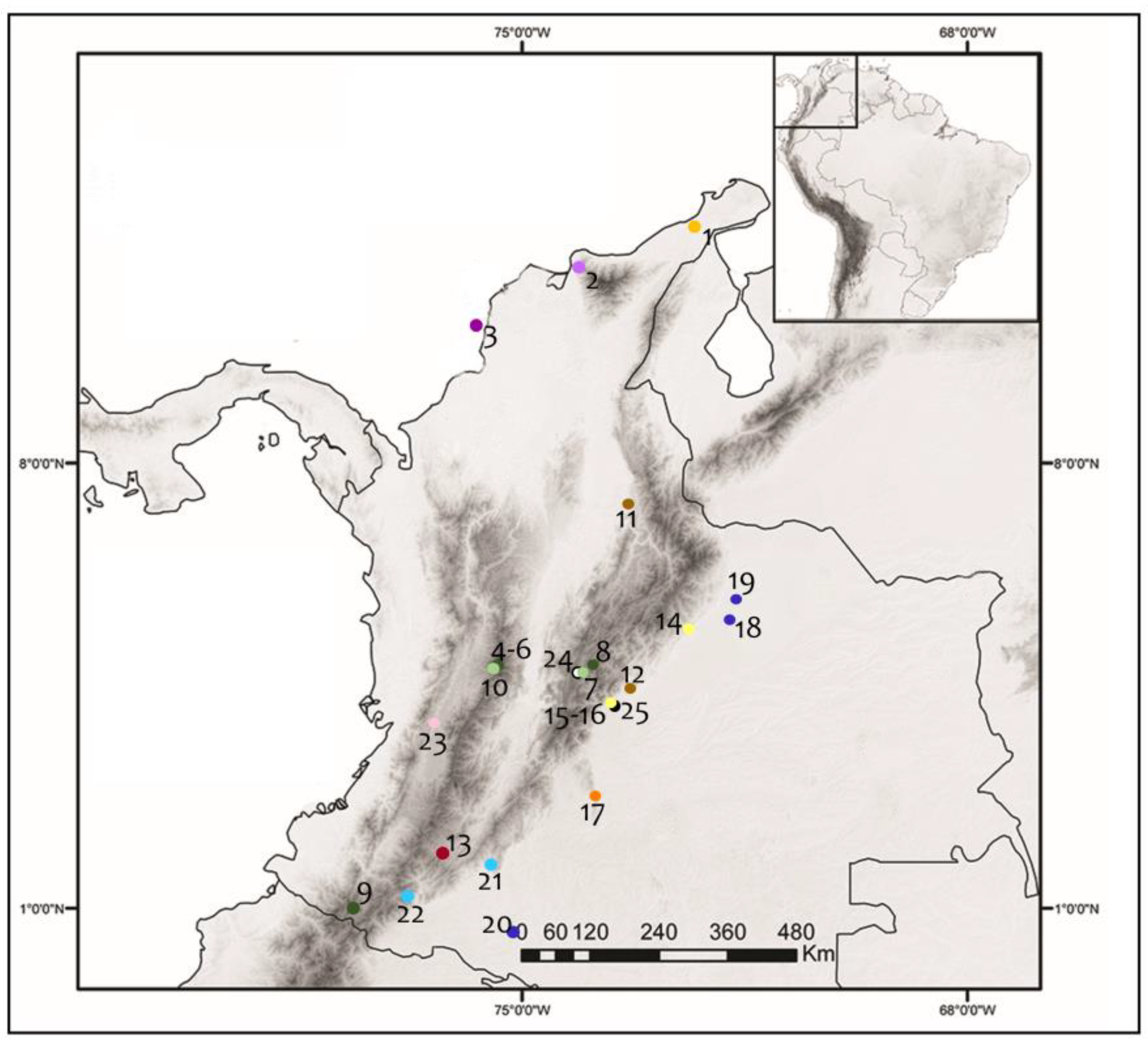
2.1. Samples
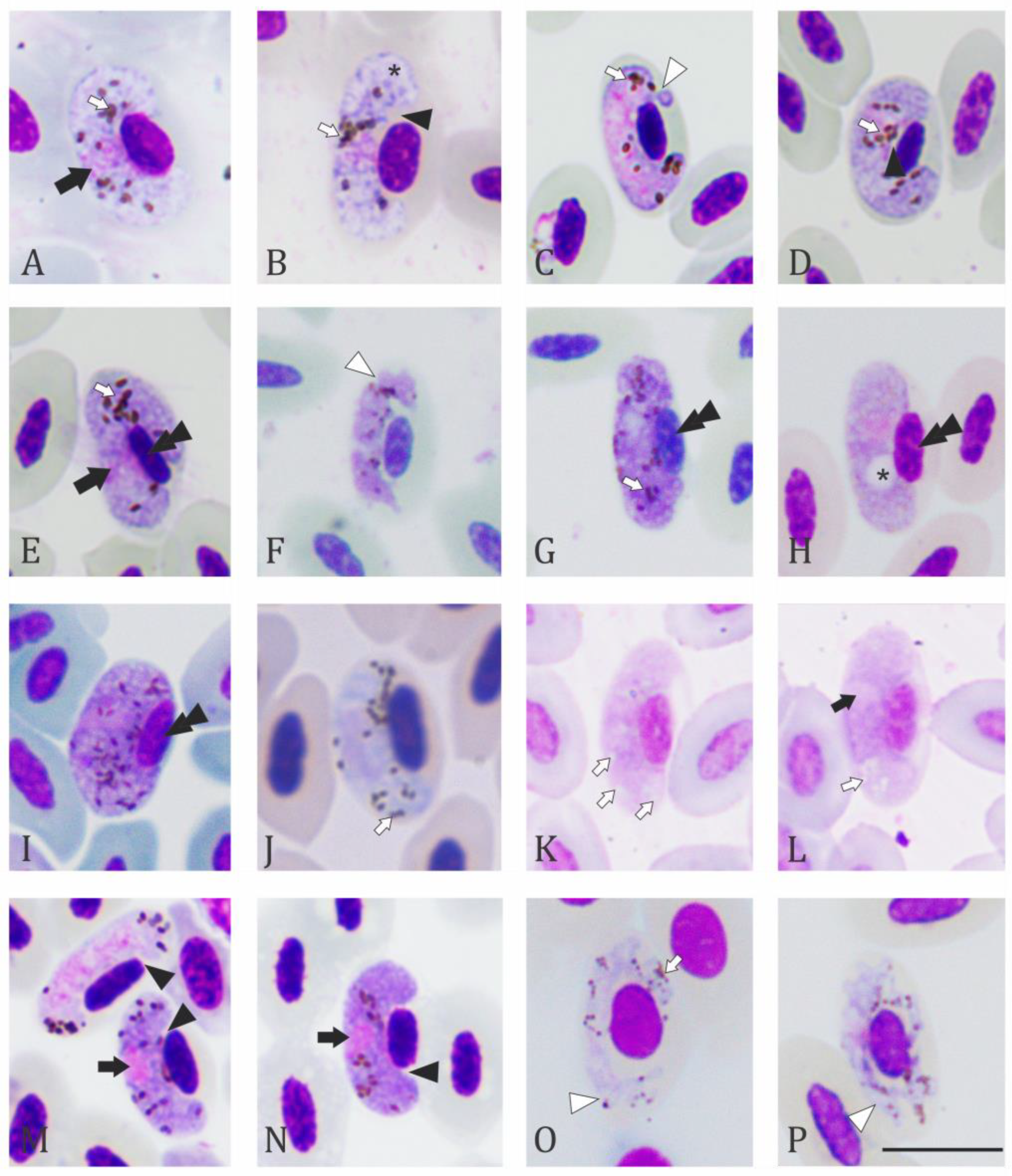
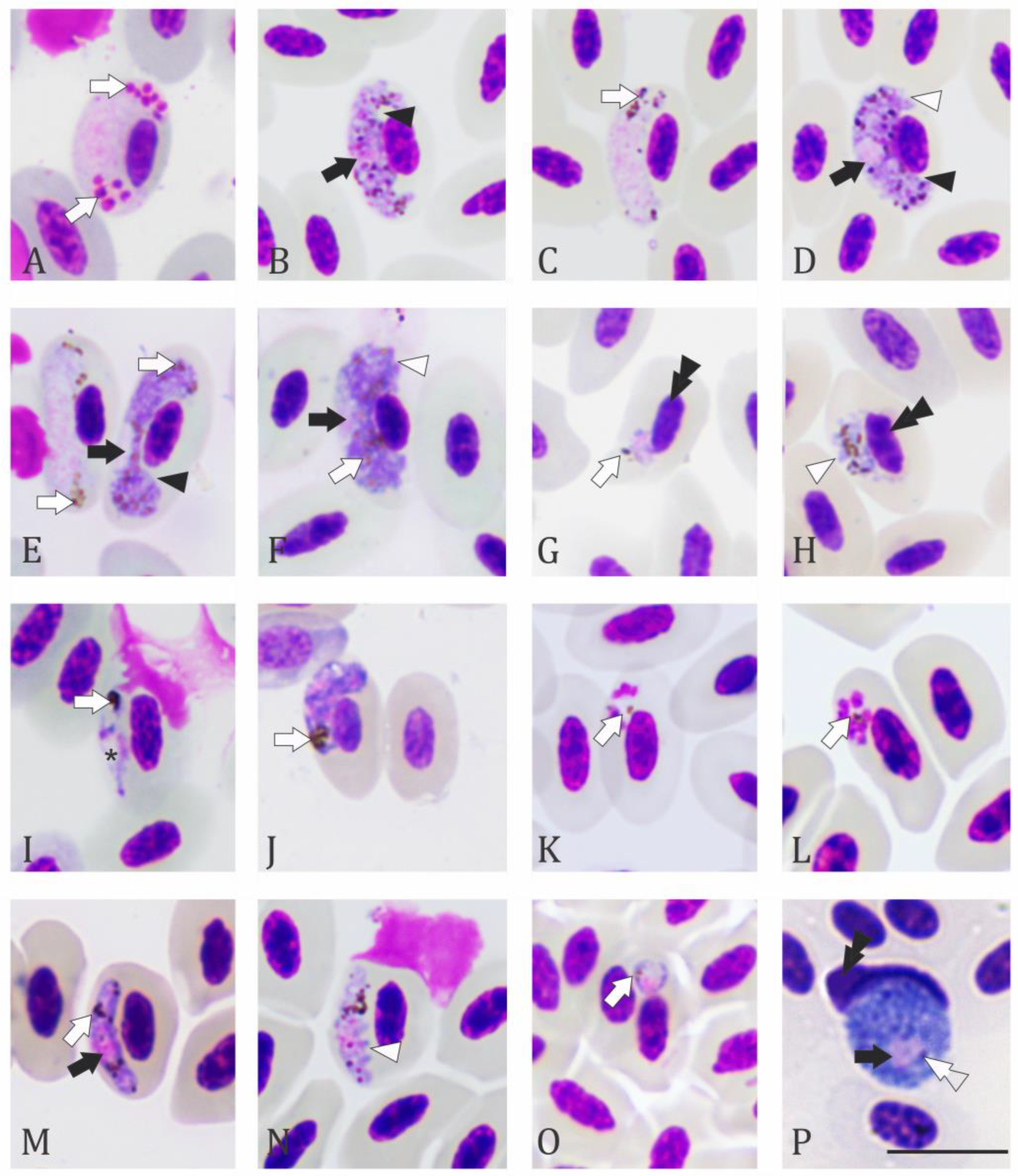
2.2. Microscopic Examination
2.3. DNA Extraction, Amplification of Cytochrome b (Cytb) Barcodes and mtDNA Genomes
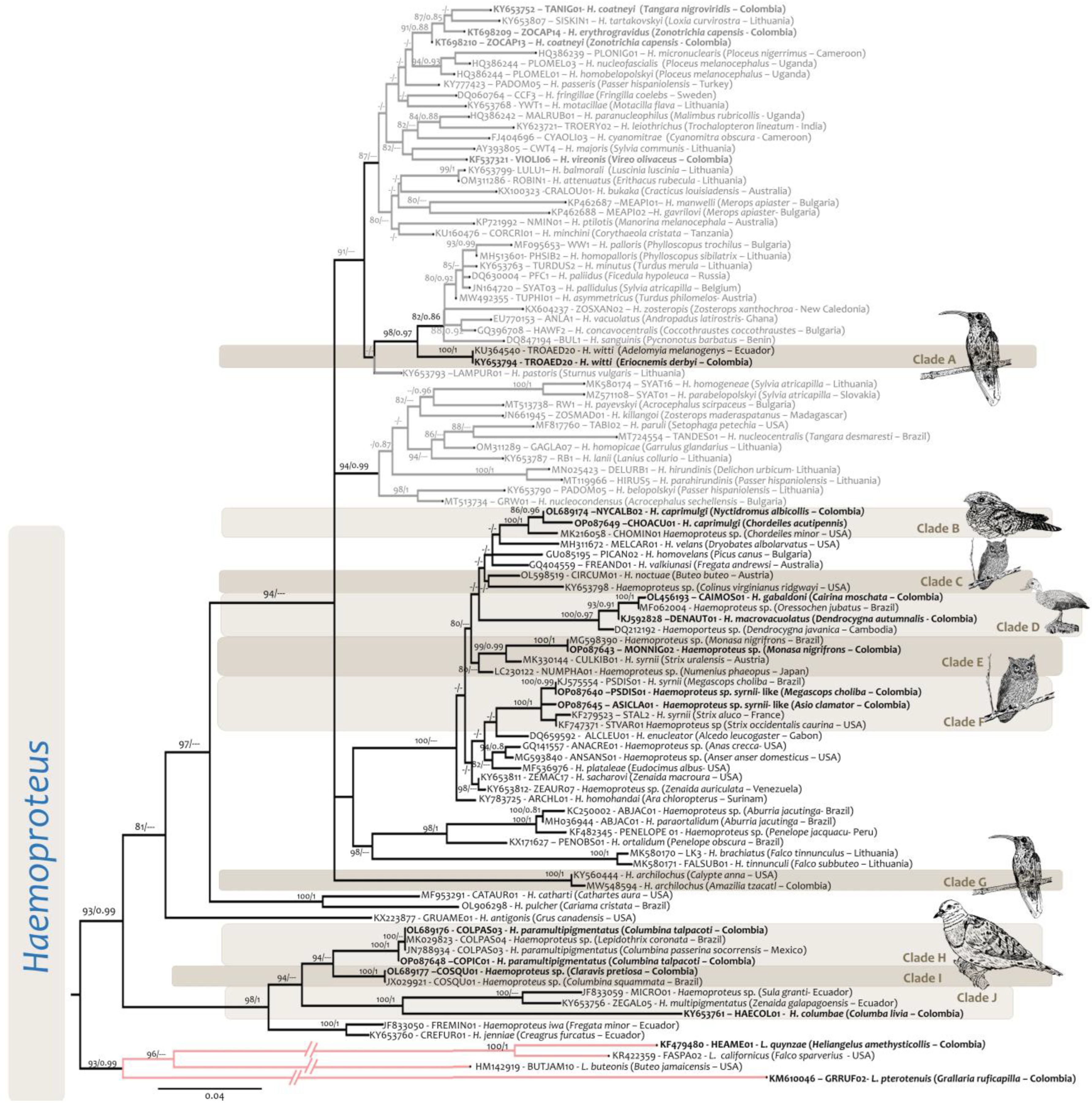
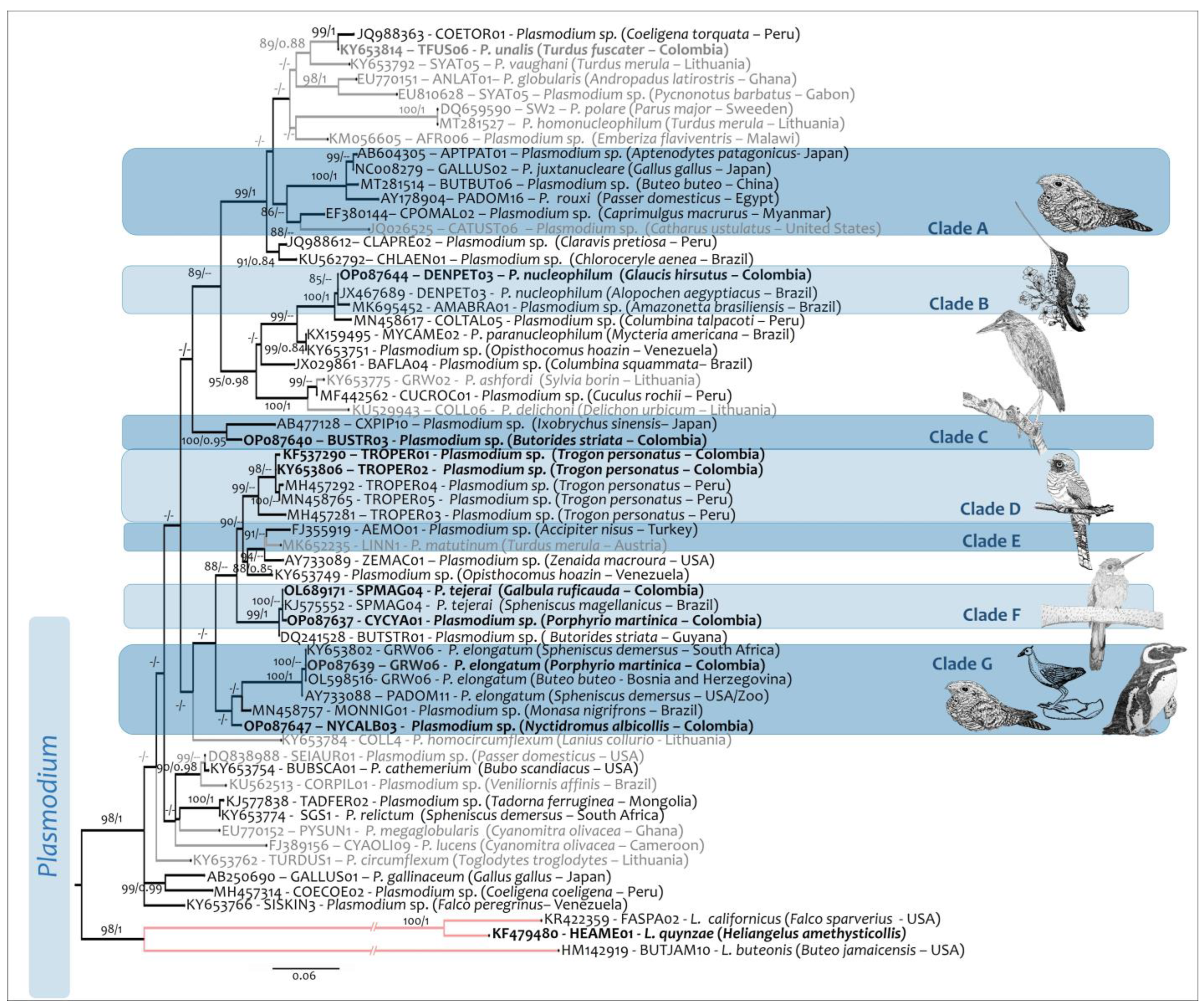
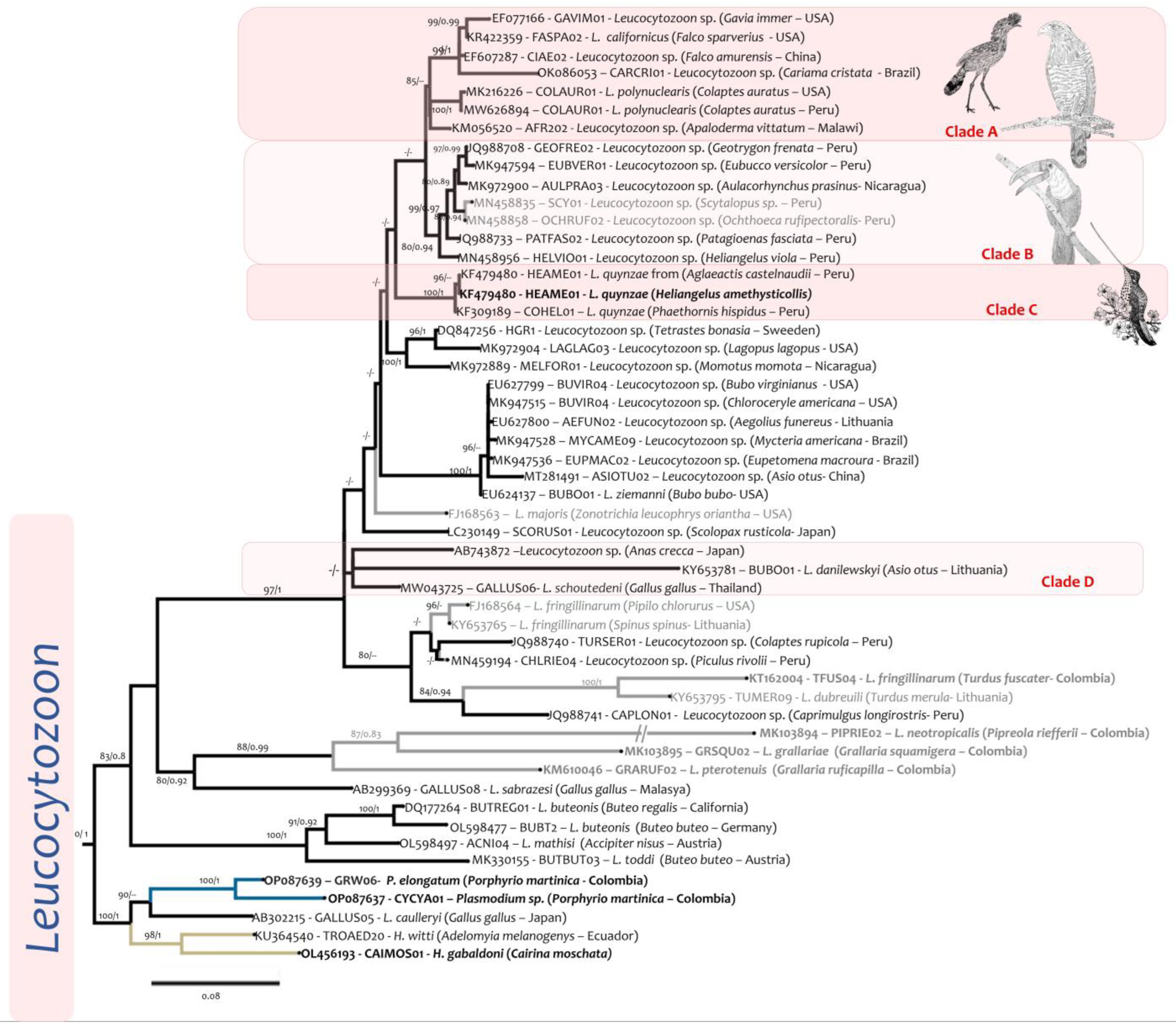
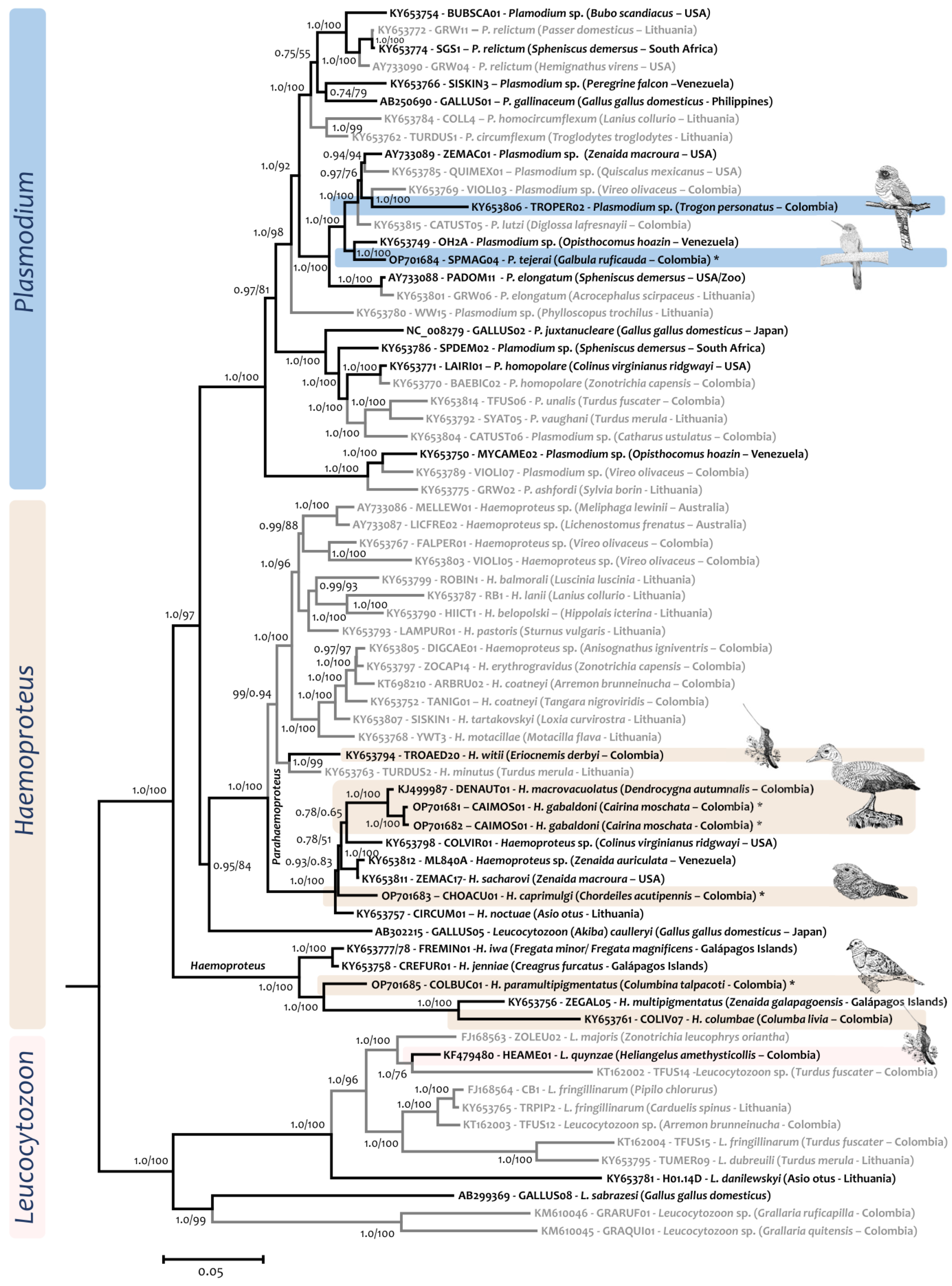
2.4. Phylogenetic Analysis
3. Results
3.1. Accipitriformes
3.2. Anseriformes
3.3. Apodiformes
3.4. Caprimulgiformes
3.5. Charadriiformes
3.6. Columbiformes
3.7. Galbuliformes
3.8. Gruiformes
3.9. Pelecaniformes
3.10. Strigiformes
3.11. Trogoniformes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levine, N. The Protozoan Phylum Apicomplexa; CRC Press: Boca Ratón, FL, USA, 1988. [Google Scholar] [CrossRef]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Ratón, FL, USA, 2005; ISBN 0-203-64379-8. [Google Scholar] [CrossRef]
- Svobodová, M.; Weidinger, K.; Peške, L.; Volf, P.; Votýpka, J.; Voříšek, P. Trypanosomes and Haemosporidia in the Buzzard (Buteo buteo) and Sparrowhawk (Accipiter nisus): Factors Affecting the Prevalence of Parasites. Parasitol. Res. 2015, 114, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Zizka, A.; Carvalho, F.A.; Scharn, R.; Bacon, C.D.; Silvestro, D.; Condamine, F.L. Amazonia Is the Primary Source of Neotropical Biodiversity. Proc. Natl. Acad. Sci. USA 2018, 115, 6034–6039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C. Terrestrial Ecoregions of the World: A New Map of Life on EarthA New Global Map of Terrestrial Ecoregions Provides an Innovative Tool for Conserving Biodiversity. BioScience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E. The Global 200: Priority Ecoregions for Global Conservation. Ann. Mo. Bot. Gard. 2002, 89, 199–224. [Google Scholar] [CrossRef]
- Birds of the World—Comprehensive Life Histories for All Bird Species and Families. Available online: https://birdsoftheworld.org/bow/home (accessed on 27 October 2022).
- Develey, P.F. Bird Conservation in Brazil: Challenges and Practical Solutions for a Key Megadiverse Country. Perspect. Ecol. Conserv. 2021, 19, 171–178. [Google Scholar] [CrossRef]
- Clark, N.J.; Clegg, S.M.; Lima, M.R. A Review of Global Diversity in Avian Haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New Insights from Molecular Data. Int. J. Parasitol. 2014, 44, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A Public Database of Malaria Parasites and Related Haemosporidians in Avian Hosts Based on Mitochondrial Cytochrome b Lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Alvarez-Londoño, J.; Cardona-Romero, M.; Martínez-Sánchez, E.T.; Ossa-López, P.A.; Pérez-Cárdenas, J.E.; Gonzalez, A.D.; Rivera-Páez, F.A.; Castaño-Villa, G.J. Avian Haemosporidian (Haemosporida: Plasmodium and Haemoproteus) in the Department of Arauca, Colombian Orinoquia Region. Parasitol. Res. 2022, 121, 1775–1787. [Google Scholar] [CrossRef]
- Anjos, C.C.; Chagas, C.R.; Fecchio, A.; Schunck, F.; Costa-Nascimento, M.J.; Monteiro, E.F.; Mathias, B.S.; Bell, J.A.; Guimarães, L.O.; Comiche, K.J. Avian Malaria and Related Parasites from Resident and Migratory Birds in the Brazilian Atlantic Forest, with Description of a New Haemoproteus Species. Pathogens 2021, 10, 103. [Google Scholar] [CrossRef]
- McNew, S.M.; Barrow, L.N.; Williamson, J.L.; Galen, S.C.; Skeen, H.R.; DuBay, S.G.; Gaffney, A.M.; Johnson, A.B.; Bautista, E.; Ordoñez, P. Contrasting Drivers of Diversity in Hosts and Parasites across the Tropical Andes. Proc. Natl. Acad. Sci. USA 2021, 118, e2010714118. [Google Scholar] [CrossRef]
- González Quevedo, C.; Pabón Vidal, A.L.; Rivera Gutiérrez, H.F. Prevalence of Haemosporidians in a Neotropical Endemic Bird Area. Avian Conserv. Ecol. 2016, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Aragão, H.D.B. Sobre o Cyclo Evolutivo Ea Transmissão Do Haemoproteus columbae. Rev. Médica De São Paulo 1908, 11, 409–416. [Google Scholar]
- Santiago-Alarcon, D.; Marzal, A. Research on Avian Haemosporidian Parasites in the Tropics before the Year 2000. In Avian Malaria and Related Parasites in the Tropics; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–44. [Google Scholar]
- White, E.M.; Greiner, E.C.; Bennett, G.F.; Herman, C.M. Distribution of the Hematozoa of Neotropical Birds. Rev. De Biol. Trop. 1978, 26, 43–102. [Google Scholar]
- Gabaldon, A.; Ulloa, G.; Zerpa, N. Fallisia (Plasmodioides) neotropicalis Subgen. Nov. Sp. Nov. from Venezuela. Parasitology 1985, 90, 217–225. [Google Scholar] [CrossRef]
- Bennett, G.F. Haemoproteus gabaldoni n. Sp.(Apicomplexa: Haemoproteidae) from the Muscovy Duck Cairina Moschata (Aves: Anatidae). Syst. Parasitol. 1993, 25, 119–123. [Google Scholar] [CrossRef]
- Matta, N.E.; Pacheco, M.A.; Escalante, A.A.; Valkiūnas, G.; Ayerbe-Quiñones, F.; Acevedo-Cendales, L.D. Description and Molecular Characterization of Haemoproteus macrovacuolatus n. Sp. (Haemosporida, Haemoproteidae), a Morphologically Unique Blood Parasite of Black-Bellied Whistling Duck (Dendrocygna autumnalis) from South America. Parasitol. Res. 2014, 11, 2991–3000. [Google Scholar] [CrossRef] [PubMed]
- Matta, N.E.; Lotta, I.A.; Valkiūnas, G.; González, A.D.; Pacheco, M.A.; Escalante, A.A.; Moncada, L.I.; Rodríguez-Fandiño, O.A. Description of Leucocytozoon quynzae Sp. Nov.(Haemosporida, Leucocytozoidae) from Hummingbirds, with Remarks on Distribution and Possible Vectors of Leucocytozoids in South America. Parasitol. Res. 2014, 113, 457–468. [Google Scholar] [CrossRef]
- Vanstreels, R.E.T.; Dos Anjos, C.C.; Leandro, H.J.; de Moraes Carvalho, A.; Santos, A.P.; Egert, L.; Hurtado, R.; de Carvalho, E.C.Q.; Braga, É.M.; Kirchgatter, K. A New Haemosporidian Parasite from the Red-Legged Seriema cariama cristata (Cariamiformes, Cariamidae). Int. J. Parasitol. Parasites Wildl. 2022, 18, 12–19. [Google Scholar] [CrossRef]
- White, E.M.; Bennett, G.F.; Williams, N.A. Avian Haemoproteidae. 11. The Haemoproteids of the Hummingbird Family Trochilidae. Can. J. Zool. 1979, 57, 908–913. [Google Scholar] [CrossRef]
- Durrant, K.L.; Beadell, J.S.; Ishtiaq, F.; Graves, G.R.; Olson, S.L.; Gering, E.; Peirce, M.A.; Milensky, C.M.; Schmidt, B.K.; Gebhard, C. Avian Hematozoa in South America: A Comparison of Temperate and Tropical Zones. Ornithol. Monogr. 2006, 60, 98–111. [Google Scholar] [CrossRef]
- Pacheco, M.A.; Ferreira, F.C.; Logan, C.J.; McCune, K.B.; MacPherson, M.P.; Albino Miranda, S.; Santiago-Alarcon, D.; Escalante, A.A. Great-Tailed Grackles (Quiscalus mexicanus) as a Tolerant Host of Avian Malaria Parasites. PloS ONE 2022, 17, e0268161. [Google Scholar] [CrossRef] [PubMed]
- Harrigan, R.J.; Sedano, R.; Chasar, A.C.; Chaves, J.A.; Nguyen, J.T.; Whitaker, A.; Smith, T.B. New Host and Lineage Diversity of Avian Haemosporidia in the Northern Andes. Evol. Appl. 2014, 7, 799–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotta, I.A.; Pacheco, M.A.; Escalante, A.A.; González, A.D.; Mantilla, J.S.; Moncada, L.I.; Adler, P.H.; Matta, N.E. Leucocytozoon Diversity and Possible Vectors in the Neotropical Highlands of Colombia. Protist 2016, 167, 185–204. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A. Keys to the Avian Malaria Parasites. Malar. J. 2018, 17, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; Volume 2. [Google Scholar]
- Hellgren, O.; Waldenström, J.; Bensch, S. A New PCR Assay for Simultaneous Studies of Leucocytozoon, Plasmodium, and Haemoproteus from Avian Blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, M.A.; Cepeda, A.S.; Bernotienė, R.; Lotta, I.A.; Matta, N.E.; Valkiūnas, G.; Escalante, A.A. Primers Targeting Mitochondrial Genes of Avian Haemosporidians: PCR Detection and Differential DNA Amplification of Parasites Belonging to Different Genera. Int. J. Parasitol. 2018, 48, 657–670. [Google Scholar] [CrossRef]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Örjan, Ö.; Hannson, B.; Westerdahl, H.; Pinheiro, R.T. Host Specificity in Avian Blood Parasites: A Study of Plasmodium and Haemoproteus Mitochondrial DNA Amplified from Birds. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, M.A.; Matta, N.E.; Valkiūnas, G.; Parker, P.G.; Mello, B.; Stanley Jr, C.E.; Lentino, M.; Garcia-Amado, M.A.; Cranfield, M.; Kosakovsky Pond, S.L. Mode and Rate of Evolution of Haemosporidian Mitochondrial Genomes: Timing the Radiation of Avian Parasites. Mol. Biol. Evol. 2018, 35, 383–403. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A Fast Online Phylogenetic Tool for Maximum Likelihood Analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2009, 27, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirce, M.; Marquiss, M. Haematozoa of British Birds. VII. Haematozoa of Raptors in Scotland with a Description of Haemoproteus nisi Sp. Nov. from the Sparrowhawk (Accipiter nisus). J. Nat. Hist. 1983, 17, 813–821. [Google Scholar] [CrossRef]
- González, A.D.; Lotta-Arevalo, I.; Fuentes, G.A.; Macías-Zacipa, J.; Acevedo-Cendales, L.D.; Matta, N.E. Haemoproteus gabaldoni a Valid Species? An Approach from Morphology and Molecular Tools Applied to Parasites of Anseriformes. Acta Trop. 2022, 233, 106540. [Google Scholar] [CrossRef]
- Cleland, S.J.B.; Johnston, T.H. Descriptions of New Haemoprotozoa from Birds in New South Wales, with a Note on the Resemblance between the Spermatozoa of Certain Honeyeaters (Fam. Meliphagidae) and Spirochaete-Trypanosomes; Royal Society of New South Wales: Sydney, Australia, 1909. [Google Scholar]
- Pacheco, M.A.; Escalante, A.A.; Garner, M.M.; Bradley, G.A.; Aguilar, R.F. Haemosporidian Infection in Captive Masked Bobwhite Quail (Colinus virginianus ridgwayi), an Endangered Subspecies of the Northern Bobwhite Quail. Vet. Parasitol. 2011, 182, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, O.A.; Matta, N.E. Blood Parasites in Some Birds from Eastern Plains of Colombia. Mem. Do Inst. Oswaldo Cruz 2001, 96, 1173–1176. [Google Scholar] [CrossRef] [Green Version]
- González, A.D.; Lotta, I.A.; García, L.F.; Moncada, L.I.; Matta, N.E. Avian Haemosporidians from Neotropical Highlands: Evidence from Morphological and Molecular Data. Parasitol. Int. 2015, 64, 48–59. [Google Scholar] [CrossRef]
- Galen, S.C.; Witt, C.C. Diverse Avian Malaria and Other Haemosporidian Parasites in Andean House Wrens: Evidence for Regional Co-diversification by Host-switching. J. Avian Biol. 2014, 45, 374–386. [Google Scholar] [CrossRef]
- Bradshaw, A.; Tell, L.A.; Ernest, H.B.; Bahan, S.; Carlson, J.; Sehgal, R. Detection and Prevalence of Haemoproteus archilochus (Haemosporida, Haemoproteidae) in Two Species of California Hummingbirds. Parasitol. Res. 2017, 116, 1879–1885. [Google Scholar] [CrossRef]
- Duarte-Moreno, A.N.; Villamizar-Escalante, D.; Rondón-González, F. PCR Detection of Haemoproteus archilochus in Rufous-Tailed Hummingbird Amazilia tzacatl (Trochilidae) in Colombia. Acta Biológica Colomb. 2022, 27, 140–143. [Google Scholar] [CrossRef]
- Manwell, R.D. How Many Species of Avian Malaria Parasites Are There? Am. J. Trop. Med. 1935, 15, 265–282. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Valkiūnas, G.; Nery, C.V.C.; Henrique, P.C.; Gonzalez, I.H.L.; Monteiro, E.F.; de Oliveira Guimarães, L.; Romano, C.M.; Kirchgatter, K. Plasmodium (Novyella) nucleophilum from an Egyptian Goose in São Paulo Zoo, Brazil: Microscopic Confirmation and Molecular Characterization. Int. J. Parasitol. Parasites Wildl. 2013, 2, 286–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, N.A.; Bennett, G.F.; Mahrt, J. Avian Haemoproteidae. 6. Description of Haemoproteus caprimulgi Sp. Nov., and a Review of the Haemoproteids of the Family Caprimulgidae. Can. J. Zool. 1975, 53, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.F. Avian Haemoproteidae. 10. The Haemoproteids of the Avian Family Scolopacidae. Can. J. Zool. 1979, 57, 901–907. [Google Scholar] [CrossRef]
- Kruse, W. Ueber Blutparasiten. Arch. Für Pathol. Anat. Und Physiol. Und Für Klin. Med. 1890, 120, 541–560. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A.; Evans, E.; Carlson, J.S.; Martínez-Gómez, J.E.; Sehgal, R.N. Two New Haemoproteus Species (Haemosporida: Haemoproteidae) from Columbiform Birds. J. Parasitol. 2013, 99, 513–521. [Google Scholar] [CrossRef]
- Cepeda, A.S.; Lotta-Arévalo, I.A.; Pinto-Osorio, D.F.; Macías-Zacipa, J.; Valkiūnas, G.; Barato, P.; Matta, N.E. The Experimental Characterization of Complete Life Cycle of Haemoproteus columbae, with Description of Natural Host-Parasite System to Study This Infection. Int. J. Parasitol. 2019, 49, 975–984. [Google Scholar] [CrossRef]
- Gabaldon, A.; Ulloa, G. Plasmodium (Haemamoeba) tejerai Sp. n. of the Domestic Turkey (Meleagris gallopavo) of Venezuela. Boletín De La Dir. De Malariol. Y Saneam. Ambient. 1977, 17, 255–273. [Google Scholar]
- Silveira, P.; Belo, N.O.; Lacorte, G.A.; Kolesnikovas, C.K.; Vanstreels, R.E.; Steindel, M.; Catão-Dias, J.L.; Valkiūnas, G.; Braga, E.M. Parasitological and New Molecular-Phylogenetic Characterization of the Malaria Parasite Plasmodium tejerai in South American Penguins. Parasitol. Int. 2013, 62, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Vanstreels, R.E.T.; Kolesnikovas, C.K.; Sandri, S.; Silveira, P.; Belo, N.O.; Ferreira Junior, F.C.; Epiphanio, S.; Steindel, M.; Braga, E.M.; Catao-Dias, J.L. Outbreak of Avian Malaria Associated to Multiple Species of Plasmodium in Magellanic Penguins Undergoing Rehabilitation in Southern Brazil. PLoS ONE 2014, 9, e94994. [Google Scholar] [CrossRef]
- Fecchio, A.; Bell, J.A.; Pinheiro, R.B.; Cueto, V.R.; Gorosito, C.A.; Lutz, H.L.; Gaiotti, M.G.; Paiva, L.V.; França, L.F.; Toledo-Lima, G. Avian Host Composition, Local Speciation and Dispersal Drive the Regional Assembly of Avian Malaria Parasites in South American Birds. Mol. Ecol. 2019, 28, 2681–2693. [Google Scholar] [CrossRef]
- Pacheco, M.A.; García-Amado, M.A.; Manzano, J.; Matta, N.E.; Escalante, A.A. Blood Parasites Infecting the Hoatzin (Opisthocomus hoazin), a Unique Neotropical Folivorous Bird. PeerJ 2019, 7, e6361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, C.G. Plasmodium elongatum n. Sp., an Avian Malarial Organism with an Elongate Gametocyte. Am. J. Hyg. 1930, 11, 385–391. [Google Scholar] [CrossRef]
- Palinauskas, V.; Žiegytė, R.; Iezhova, T.A.; Ilgūnas, M.; Bernotienė, R.; Valkiūnas, G. Description, Molecular Characterisation, Diagnostics and Life Cycle of Plasmodium elongatum (Lineage PERIRUB01), the Virulent Avian Malaria Parasite. Int. J. Parasitol. 2016, 46, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Nedorost, N.; Matt, J.; Kübber-Heiss, A.; Alic, A.; Konicek, C.; Weissenböck, H. Avian Haemosporidian Parasites of Accipitriform Raptors. Malar. J. 2022, 21, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Barino, G.T.M.; Rossi, M.F.; de Oliveira, L.; Reis Junior, J.L.; D’Agosto, M.; Dias, R.J.P. Haemoproteus syrnii (Haemosporida: Haemoproteidae) in Owls from Brazil: Morphological and Molecular Characterization, Potential Cryptic Species, and Exo-Erythrocytic Stages. Parasitol. Res. 2021, 120, 243–255. [Google Scholar] [CrossRef]
- DeBrock, S.; Cohen, E.; Balasubramanian, S.; Marra, P.P.; Hamer, S.A. Characterization of the Plasmodium and Haemoproteus Parasite Community in Temperate-Tropical Birds during Spring Migration. Int. J. Parasitol. Parasites Wildl. 2021, 15, 12–21. [Google Scholar] [CrossRef]
- Ellis, V.A.; Fecchio, A.; Ricklefs, R.E. Haemosporidian Parasites of Neotropical Birds: Causes and Consequences of Infection. Auk 2020, 137, ukaa055. [Google Scholar] [CrossRef]
- Bernal, R.; Gradstein, S.; Celis, M. Catalogue of Plants and Lichens of Colombia; JRS Biodiversity Foundation: Arlington, VA, USA, 2015. [Google Scholar]
- Matoso, R.V.; Cedrola, F.; Barino, G.T.M.; Dias, R.J.P.; Rossi, M.F.; D’Agosto, M. New Morphological and Molecular Data for Haemoproteus (H.) paramultipigmentatus in the Atlantic Forest of Brazil. Parasitol. Int. 2021, 84, 102375. [Google Scholar] [CrossRef] [PubMed]
- LaPointe, D.A.; Goff, M.L.; Atkinson, C.T. Thermal Constraints to the Sporogonic Development and Altitudinal Distribution of Avian Malaria Plasmodium relictum in Hawai’i. J. Parasitol. 2010, 96, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.M.; Ferreira, F.C.; Araújo, A.C.; Hirano, L.Q.L.; Paludo, G.R.; Braga, É.M. Molecular Detection of Leucocytozoon in Red-Legged Seriemas (Cariama cristata), a Non-Migratory Bird Species in the Brazilian Cerrado. Vet. Parasitol. Reg. Stud. Rep. 2022, 31, 100652. [Google Scholar] [CrossRef] [PubMed]
- Fecchio, A.; Silveira, P.; Weckstein, J.; Dispoto, J.; Anciães, M.; Bosholn, M.; Tkach, V.; Bell, J. First Record of Leucocytozoon (Haemosporida: Leucocytozoidae) in Amazonia: Evidence for Rarity in Neotropical Lowlands or Lack of Sampling for This Parasite Genus? J. Parasitol. 2018, 104, 168–172. [Google Scholar] [CrossRef]
- Rodríguez, O.A.; Moya, H.; Matta, N.E. Avian Blood Parasites in the National Natural Park Chingaza: High Andes of Colombia. Hornero 2009, 24, 1–6. [Google Scholar]
- Fecchio, A.; Ellis, V.A.; Bell, J.A.; Andretti, C.B.; D’horta, F.M.; Silva, A.M.; Tkach, V.V.; Weckstein, J.D. Avian Malaria, Ecological Host Traits and Mosquito Abundance in Southeastern Amazonia. Parasitology 2017, 144, 1117–1132. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E.; Swanson, B.L.; Fallon, S.M.; MartÍnez-AbraÍn, A.; Scheuerlein, A.; Gray, J.; Latta, S.C. Community Relationships of Avian Malaria Parasites in Southern Missouri. Ecol. Monogr. 2005, 75, 543–559. [Google Scholar] [CrossRef]
- Smith, R.B.; Greiner, E.C.; Wolf, B.O. Migratory Movements of Sharp-Shinned Hawks (Accipiter striatus) Captured in New Mexico in Relation to Prevalence, Intensity, and Biogeography of Avian Hematozoa. Auk 2004, 121, 837–846. [Google Scholar] [CrossRef]
- Ramey, A.M.; Reed, J.A.; Walther, P.; Link, P.; Schmutz, J.A.; Douglas, D.C.; Stallknecht, D.E.; Soos, C. Evidence for the Exchange of Blood Parasites between North America and the Neotropics in Blue-Winged Teal (Anas discors). Parasitol. Res. 2016, 115, 3923–3939. [Google Scholar] [CrossRef]
- Boal, C.W.; Hudelson, K.S.; Mannan, R.W.; Estabrook, T.S. Hematology and Hematozoa of Adult and Nestling Cooper’s Hawks in Arizona. J. Raptor Res. 1998, 32, 281–285. [Google Scholar]
- Reeves, A.B.; Smith, M.M.; Meixell, B.W.; Fleskes, J.P.; Ramey, A.M. Genetic Diversity and Host Specificity Varies across Three Genera of Blood Parasites in Ducks of the Pacific Americas Flyway. PLoS ONE 2015, 10, e0116661. [Google Scholar] [CrossRef] [PubMed]
- Pigeault, R.; Vézilier, J.; Cornet, S.; Zélé, F.; Nicot, A.; Perret, P.; Gandon, S.; Rivero, A. Avian Malaria: A New Lease of Life for an Old Experimental Model to Study the Evolutionary Ecology of Plasmodium. Philos. Trans. R. Soc. B 2015, 370, 20140300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagas, C.R.F.; Valkiūnas, G.; de Oliveira Guimarães, L.; Monteiro, E.F.; Guida, F.J.V.; Simões, R.F.; Rodrigues, P.T.; de Albuquerque Luna, E.J.; Kirchgatter, K. Diversity and Distribution of Avian Malaria and Related Haemosporidian Parasites in Captive Birds from a Brazilian Megalopolis. Malar. J. 2017, 16, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cocumelli, C.; Iurescia, M.; Diaconu, E.L.; Galietta, V.; Raso, C.; Buccella, C.; Stravino, F.; Grande, F.; Fiorucci, L.; De Liberato, C. Plasmodium matutinum Causing Avian Malaria in Lovebirds (Agapornis roseicollis) Hosted in an Italian Zoo. Microorganisms 2021, 9, 1356. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Huang, X.; Valkiūnas, G.; Yang, M.; Zheng, C.; Pu, T.; Zhang, Y.; Dong, L.; Suo, X.; Zhang, C. Malaria Parasites and Related Haemosporidians Cause Mortality in Cranes: A Study on the Parasites Diversity, Prevalence and Distribution in Beijing Zoo. Malar. J. 2018, 17, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanstreels, R.E.T.; de Angeli Dutra, D.; Ferreira-Junior, F.C.; Hurtado, R.; Egert, L.; Mayorga, L.F.S.; Bhering, R.C.; Braga, É.M.; Catão-Dias, J.L. Epidemiology, Hematology, and Unusual Morphological Characteristics of Plasmodium during an Avian Malaria Outbreak in Penguins in Brazil. Parasitol. Res. 2019, 118, 3497–3508. [Google Scholar] [CrossRef]
- Ishtiaq, F.; Rao, M.; Huang, X.; Bensch, S. Estimating Prevalence of Avian Haemosporidians in Natural Populations: A Comparative Study on Screening Protocols. Parasites Vectors 2017, 10, 127. [Google Scholar] [CrossRef] [Green Version]
- Almeida, G.G.; Costa, P.A.C.; da Silva Araujo, M.; Gomes, G.R.; Carvalho, A.F.; Figueiredo, M.M.; Pereira, D.B.; Tada, M.S.; Medeiros, J.F.; da Silva Soares, I. Asymptomatic Plasmodium vivax Malaria in the Brazilian Amazon: Submicroscopic Parasitemic Blood Infects Nyssorhynchus darlingi. PLoS Negl. Trop. Dis. 2021, 15, e0009077. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Loiseau, C.; Sehgal, R.N. Nested Cytochrome b Polymerase Chain Reaction Diagnostics Detect Sporozoites of Hemosporidian Parasites in Peripheral Blood of Naturally Infected Birds. J. Parasitol. 2009, 95, 1512–1515. [Google Scholar] [CrossRef]
- Gadalla, A.A.; Siciliano, G.; Farid, R.; Alano, P.; Ranford-Cartwright, L.; McCarthy, J.S.; Thompson, J.; Babiker, H.A. Real-Time PCR Assays for Detection and Quantification of Early P. falciparum Gametocyte Stages. Sci. Rep. 2021, 11, 19118. [Google Scholar] [CrossRef]
- Bertram, M.R.; Hamer, S.A.; Hartup, B.K.; Snowden, K.F.; Medeiros, M.C.; Outlaw, D.C.; Hamer, G.L. A Novel Haemosporida Clade at the Rank of Genus in North American Cranes (Aves: Gruiformes). Mol. Phylogenet. Evol. 2017, 109, 73–79. [Google Scholar] [CrossRef]
- Yabsley, M.J.; Vanstreels, R.E.; Martinsen, E.S.; Wickson, A.G.; Holland, A.E.; Hernandez, S.M.; Thompson, A.T.; Perkins, S.L.; West, C.J.; Bryan, A.L. Parasitaemia Data and Molecular Characterization of Haemoproteus catharti from New World Vultures (Cathartidae) Reveals a Novel Clade of Haemosporida. Malar. J. 2018, 17, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lotta, I.A.; Valkiūnas, G.; Pacheco, M.A.; Escalante, A.A.; Hernández, S.R.; Matta, N.E. Disentangling Leucocytozoon Parasite Diversity in the Neotropics: Descriptions of Two New Species and Shortcomings of Molecular Diagnostics for Leucocytozoids. Int. J. Parasitol. Parasites Wildl. 2019, 9, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, A.; de la Puente, J.; Onrubia, A.; Pérez-Tris, J. Molecular Characterization of Haemosporidian Parasites from Kites of the Genus Milvus (Aves: Accipitridae). Int. J. Parasitol. 2013, 43, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Honjo, Y.; Fukumoto, S.; Sakamoto, H.; Hikosaka, K. New PCR Primers Targeting on a Cytochrome b Gene Reveal Diversity of Leucocytozoon Lineages in an Individual Host. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Wandeler, P.; Hoeck, P.E.; Keller, L.F. Back to the Future: Museum Specimens in Population Genetics. Trends Ecol. Evol. 2007, 22, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Galen, S.C.; Nunes, R.; Sweet, P.R.; Perkins, S.L. Integrating Coalescent Species Delimitation with Analysis of Host Specificity Reveals Extensive Cryptic Diversity despite Minimal Mitochondrial Divergence in the Malaria Parasite Genus Leucocytozoon. BMC Evol. Biol. 2018, 18, 128. [Google Scholar] [CrossRef]
- Lepage, D.; Warnier, J. The Peters’ Check-List of the Birds of the World (1931–1987) Database. Available online: https://avibase.bsc-eoc.org/peterschecklist.jsp (accessed on 30 July 2022).
- Instituto de Hidrología. Meteorología y Estudios Ambientales Atlas Climatológico de Colombia. Available online: http://atlas.ideam.gov.co/visorAtlasClimatologico.html (accessed on 12 July 2022).
- Gómez Mercado, J.M.; Martínez Hincapie, Y.; Mosalvo, A.; Carpio, E.; Goyeneche, S.; Lubo, H.; Herran, V.; Gutierrez, W.; Frias, R.; Deluque, A. Plan de Desarrollo Municipal 2016–2019 Manaure Equidad, Efectividad y Ciudadanía. Available online: http://www.manaureguajira.gov.co/Documents/plandedesarrollo.pdf (accessed on 14 July 2022).
- Ministerio de Ambiente, Vivienda y Desarrollo Territorial; Corporación Autónoma Regional del Magdalena (Corpamag); Instituto de Investigacines Marinas y Costeras, «José Benito Vives de Andreis, Invemar» Plan de Manejo Santuario de Flora y Fauna de La Ciénaga Grande de Santa Marta. Available online: https://www.corpamag.gov.co/archivos/PMA/PlanManejoRBRamsar.pdf (accessed on 30 July 2022).
- Valle, A.G.; Osorno-Arango, A.M.; Gil-Agudelo, D.L. Estructura y Regeneración Del Bosque de Manglar de La Ciénaga de Cholón, Isla Barú, Parque Nacional Natural Corales Del Rosario y San Bernardo, Caribe Colombiano. Boletín De Investig. Mar. Y Costeras-INVEMAR 2011, 40, 115–130. [Google Scholar]
- Gil-Ospina, R.F.; Bedoya-Zuluaga, F.A.; Castaño-Villa, G.J. Tendencias Poblacionales En Algunas Especies de Aves Acuáticas En La Laguna Del Otún Entre 1998 y 2007. Bol. Cient. Mus. Hist. Nat. 2010, 14, 92–98. [Google Scholar]
- Rivera, D. Páramos de Colombia. Banco De Occidente. I/M Editores. Imprelibros Santiago De Cali Colomb. 2001. [Google Scholar]
- Cadena, D.; Estacio, H.; Erazo, L. Plan de Desarrollo “Es Tiempo de Avanzar”. Available online: http://www.pupiales-narino.gov.co/apc-aaaafiles/33663063366461626161306634386632/plan-de-desarrollopupiales-2012-2015.pdf (accessed on 14 July 2022).
- Vargas Ríos, O.; Pedraza, P. El Parque Nacional Natural Chingaza; Universidad Nacional de Colombia, Facultad de Ciencias, Departamento de Biología: Bogotá, Colombia, 2004; ISBN 958-33-6824-5. [Google Scholar]
- Ramsar Sites Information Service Annotated List of Wetlands of International Importance Colombia. Available online: https://rsis.ramsar.org/sites/default/files/rsiswp_search/exports/Ramsar-Sites-annotated-summary-Colombia.pdf?1658250317 (accessed on 14 July 2022).
- Department of Geosciences Universidad Nacional de Colombia Boletín Del Análisis de Las Variables Obtenidas En La Estación Meteorológica de La Universidad Nacional. Available online: http://www.geociencias.unal.edu.co/?niv=not¬=545&dep=12 (accessed on 30 April 2013).
- Lentijo, G.; Kattan, G. Estratificación Vertical de Las Aves En Una Plantación Mono Específica y En Bosque Nativo En La Cordillera Central de Colombia. Ornitol. Colomb. 2005, 3, 51–61. [Google Scholar]
- Usma, J.; Trujillo, F. Biodiversidad Del Casanare: Ecosistemas Estratégicos Del Departamento; Gobernación del Casanare-WWF Colombia: Bogotá, Colombia, 2011. [Google Scholar]
- Corporacion Autonoma del Valle del Cauca Plan de Manejo Ambiental Del Complejo de Humedales Del Alto Río Cauca Asociado a La Laguna de Sonso—Designado Como Sitio RAMSAR (Valle Del Cauca). Available online: https://www.cvc.gov.co/sites/default/files/2019-02/PMA%20Sitio%20Ramsar_VF_web.pdf (accessed on 19 December 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | nT | Haemosporidian | Biome | Locality | Parasite Species | Lineage | ||
|---|---|---|---|---|---|---|---|---|
| P | H | L | ||||||
| Accipitriformes | ||||||||
| Accipitridae | ||||||||
| Rupornis magnirostris | 4 | 1 | V, VI, X, IX | 6, 12, 17, 18 | Haemoproteus sp. | - | ||
| Buteo platypterus | 1 | 1 | 24 | Haemoproteus nisi | - | |||
| Anseriformes | ||||||||
| Anatidae | ||||||||
| Anas discors | 13 | 2 | II, XII | 2, 23 | Haemoproteus sp. | - | ||
| Cairina moschata | 18 | 2 | V, VI | 11,12 | H. gabaldoni | OL456193 a/OP701681 b-OP701682 b-CAIMOS01 | ||
| Dendrocygna autumnalis | 33 | 14 | X | 19 | H. macrovacuolatus | KJ592828 a/KJ499987 b-DENAUT01 | ||
| Apodiformes | ||||||||
| Trochilidae | ||||||||
| Adelomyia melanogenys | 24 | 1 | V, VII | 5, 8, 10, 13 | L. qyunzae | KF309188 a-HEAME01 | ||
| Boissonneaua flavescens | 3 | 1 | IV, V | 4, 5, 10 | H. witti | |||
| Chalybura buffonii | 2 | 1 | VI, VII, VIII | 1, 7, 15 | Haemoproteus sp. | - | ||
| Coeligena helianthea | 16 | 1 | V, IV | 2, 7, 13 | L. qyunzae | KF309189 a-COHEL01 | ||
| Ensifera ensifera | 9 | 2 | IV | 4, 5, 7 | H. witti | KY653794 a-TROAED20 | ||
| Eriocnemis cupreoventris | 14 | 1 | IV | 5, 7, 8 | H. witti | KY653794 a-TROAED20 | ||
| Eriocnemis derbyi | 51 | 1 | IV, V | 4, 10 | H. witti | KY653794 a-TROAED20 | ||
| Eriocnemis vestita | 33 | 3 | IV | 7 | H. witti | KY653794 a-TROAED20 | ||
| Glaucis hirsutus | 17 | 1 | 1 | VIII, X, XI | 16, 20, 21 | Plasmodium nucleophilum, | OP087644 a-DENPET03 | |
| Heliangelus amethysticollis | 6 | 1 | IV, V | 5, 7, 8,9 | L. qyunzae | KF309188 a/KF479480 b-HEAME01 | ||
| Heliangelus exortis | 37 | 1 | IV, V | 4, 5, 7 | Haemoproteus sp. | - | ||
| 1 | L. quynzae | KF309188 a-HEAME01 | ||||||
| Lafresnaya lafresnayi | 19 | 1 | IV, V | 4, 5, 7 | H. witti | - | ||
| Metallura tyrianthina | 56 | 6 | IV, V | 4, 5, 7, 10 | L. qyunzae | KF309188 a-HEAME01 | ||
| Opisthoprora euryptera | 2 | 1 | IV | 5, 7 | L. qyunzae | - | ||
| Caprimulgiformes | ||||||||
| Caprimulgidae | ||||||||
| Chordeiles acutipennis | 3 | 1 | X | 19 | H. caprimulgi | OP087649 a/OP701683 b-CHOACU01 * | ||
| Nyctidromus albicollis | 3 | 2 | VIII, XI | 14, 21 | H. caprimulgi, | OL689174 a-NYCTALB02 * | ||
| 1 | Plasmodium sp. | OP087647 a-NYCTALB03 * | ||||||
| Nyctipolus nigrescens | 1 | 1 | IX | 17 | H. caprimulgi | OL689174 a-NYCTALB02 * | ||
| Charadriiformes | ||||||||
| Scolopacidae | ||||||||
| Numenius phaeopus | 13 | 1 | II, III, XII | 2, 3,23 | H. contortus | - | ||
| Columbiformes | ||||||||
| Columbidae | ||||||||
| Claravis pretiosa | 6 | 3 | VIII, IX | 14, 17 | Haemoproteus sp. | OL689177 a-COSQU01 | ||
| Columba livia | 40 | 16 | IV, V, VI | 7, 8, 12 | H. columbae | KY653761 a-HECOL01 | ||
| Columbina minuta | 4 | 1 | IX, X | 17, 18 | Haemoproteus sp. | - | ||
| Columbina passerina | 3 | 2 | VIII | 16 | Haemoproteus sp. | - | ||
| Columbina squammata | 6 | 2 | VIII, X | 14, 16, 18 | Haemoproteus sp. | |||
| Columbina talpacoti | 13 | 6 | I, VIII, X | 14, 17, 18, 19 | H. paramultipigmentatus | OL689176 a/OP701685 b-COLPAS03 OP087648 a-COPIC01 | ||
| Leptotila rufaxilla | 12 | 1 | VIII, IX, X | 14, 15, 17, 18 | Haemoproteus sp. | - | ||
| Leptotila verreauxi | 11 | 2 | V, VIII, X | 10, 14, 16, 20 | Haemoproteus sp. | - | ||
| Patagioenas cayennensis | 1 | 1 | VIII | 14 | Haemoproteus sp. | - | ||
| Zenaida auriculata | 30 | 4 | IV, V, VIII, IX, XI | 8, 10, 16, 17, 22 | Haemoproteus sp. | - | ||
| Zentrygon frenata | 1 | 1 | VII | 13 | Haemoproteus sp. | - | ||
| Galbuliformes | ||||||||
| Galbulidae | ||||||||
| Galbula ruficauda | 8 | 1 | VIII, X | 14, 18 | P. tejerai | OL689171 a/OP701684 b-SPMAG04 | ||
| Bucconidae | ||||||||
| Monasa nigrifrons | 2 | 1 | XI | 21 | Haemoproteus sp. | OP087643 a-MONNIG02 | ||
| Gruiformes | ||||||||
| Rallidae | ||||||||
| Porphyrio martinica | 30 | 9 | V, - | 7, 8, 23, 24 | Plasmodium elongatum, | OP087639 a-GRW06 | ||
| 1 | Plasmodium sp. | OP087637 a-CYCYA01 | ||||||
| Pelecaniformes | ||||||||
| Ardeidae | ||||||||
| Butorides striata | 5 | 1 | V, XII, | 8, 23, 24 | Plasmodium sp. | OP087640 a-BUSTR03 * | ||
| Strigiformes | ||||||||
| Strigidae | ||||||||
| Megascops choliba | 1 | 1 | 24 | Haemoproteus syrnii-like | OP087641 a-PSDIS01 | |||
| Asio clamator | 4 | 4 | 25 | Haemoproteus syrnii-like | OP087645 a-ASICLA01 * | |||
| Trogoniformes | ||||||||
| Trogonidae | ||||||||
| Trogon personatus | 4 | 1 | V | 4, 6, 7, 9 | Plasmodium sp. | KF537290 a-TROPER01 KY653806 b-TROPER02 | ||
| Total infected | 15 | 82 | 11 | |||||
| Non infected | 1131 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lotta-Arévalo, I.A.; González, A.D.; Gamboa-Suárez, B.A.; Pacheco, M.A.; Escalante, A.A.; Moreno, C.; Rodríguez-Fandíño, O.; Cuervo, A.; Matta, N.E. Haemosporidians in Non-Passerine Birds of Colombia: An Overview of the Last 20 Years of Research. Diversity 2023, 15, 57. https://doi.org/10.3390/d15010057
Lotta-Arévalo IA, González AD, Gamboa-Suárez BA, Pacheco MA, Escalante AA, Moreno C, Rodríguez-Fandíño O, Cuervo A, Matta NE. Haemosporidians in Non-Passerine Birds of Colombia: An Overview of the Last 20 Years of Research. Diversity. 2023; 15(1):57. https://doi.org/10.3390/d15010057
Chicago/Turabian StyleLotta-Arévalo, Ingrid Astrid, Angie Daniela González, Brayan Andrés Gamboa-Suárez, M. Andreína Pacheco, Ananías A. Escalante, Carlos Moreno, Oscar Rodríguez-Fandíño, Andrés Cuervo, and Nubia E. Matta. 2023. "Haemosporidians in Non-Passerine Birds of Colombia: An Overview of the Last 20 Years of Research" Diversity 15, no. 1: 57. https://doi.org/10.3390/d15010057