
Molecular Phylogenetics and Comparative Examination of Voucher Museums Reveal Two New Species of Gymnophthalmid Lizards (Squamata, Gymnophthalmidae) from the Peruvian Andes, with Comments on Proctoporus guentheri (Boettger, 1891) †




Abstract
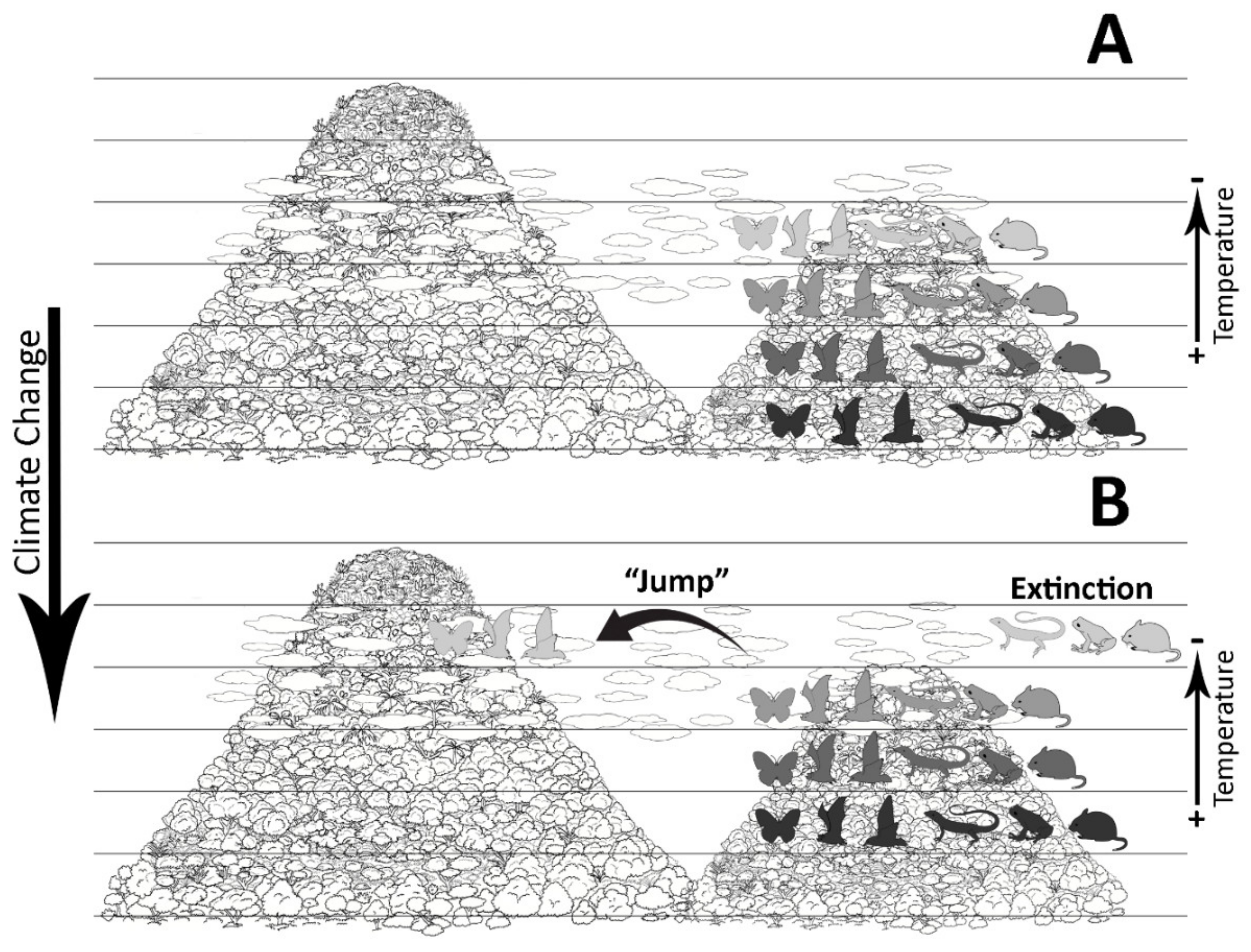
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, Amplification, and Sequencing
2.3. Phylogenetic Reconstruction
2.4. Taxonomic Treatment and Review of Museum Vouchers
3. Results
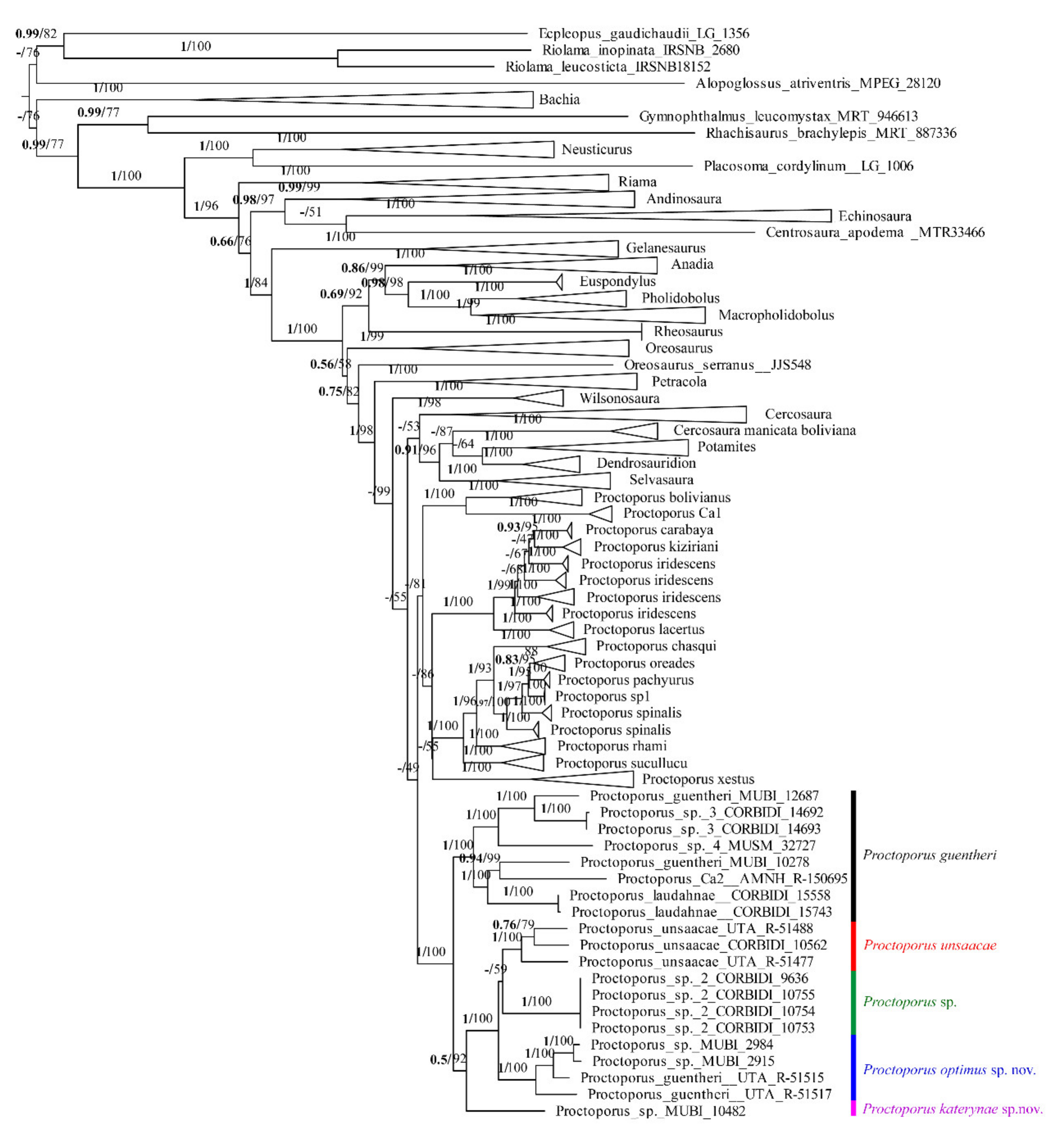
3.1. Phylogenetic Relationships
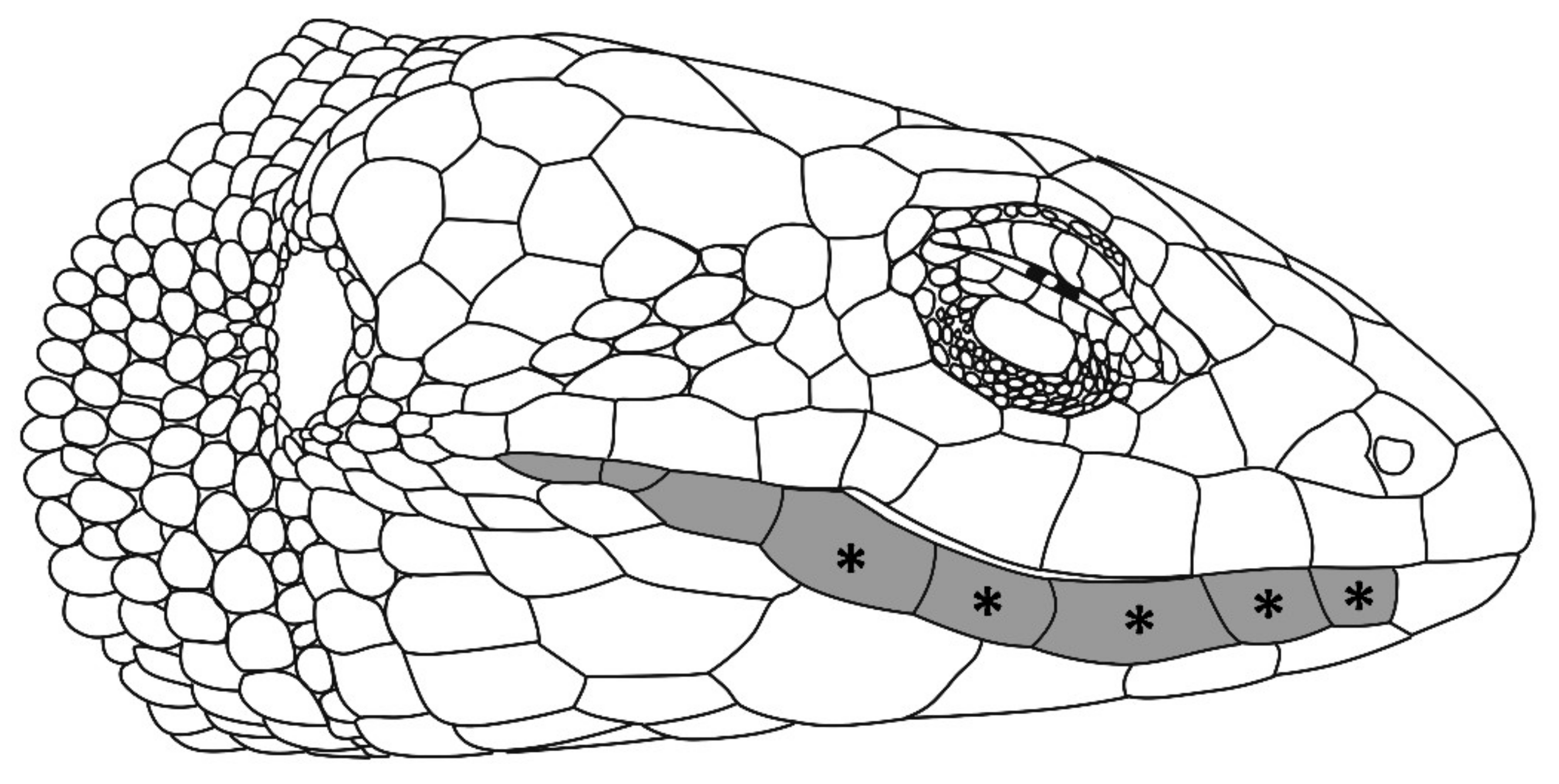
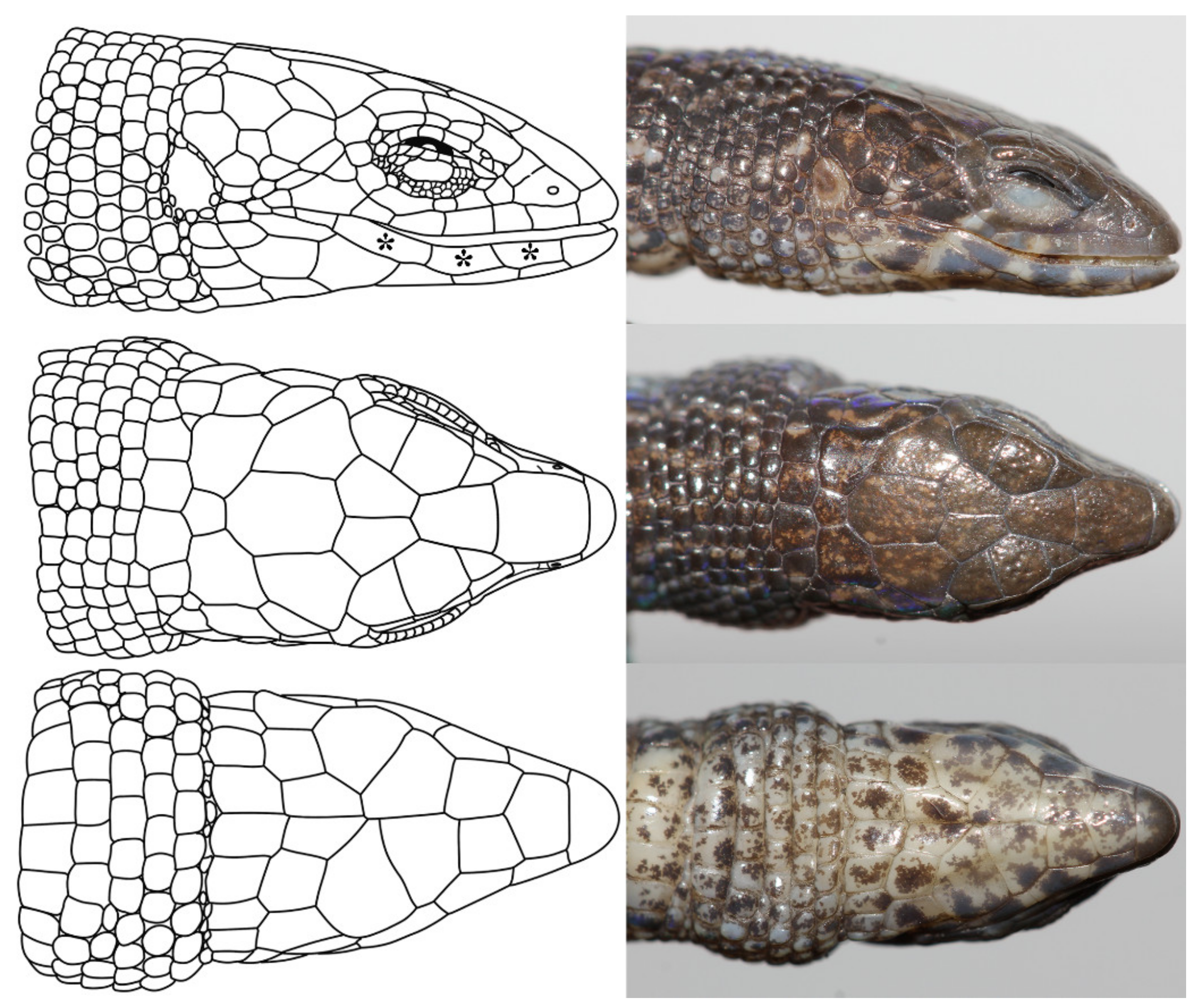
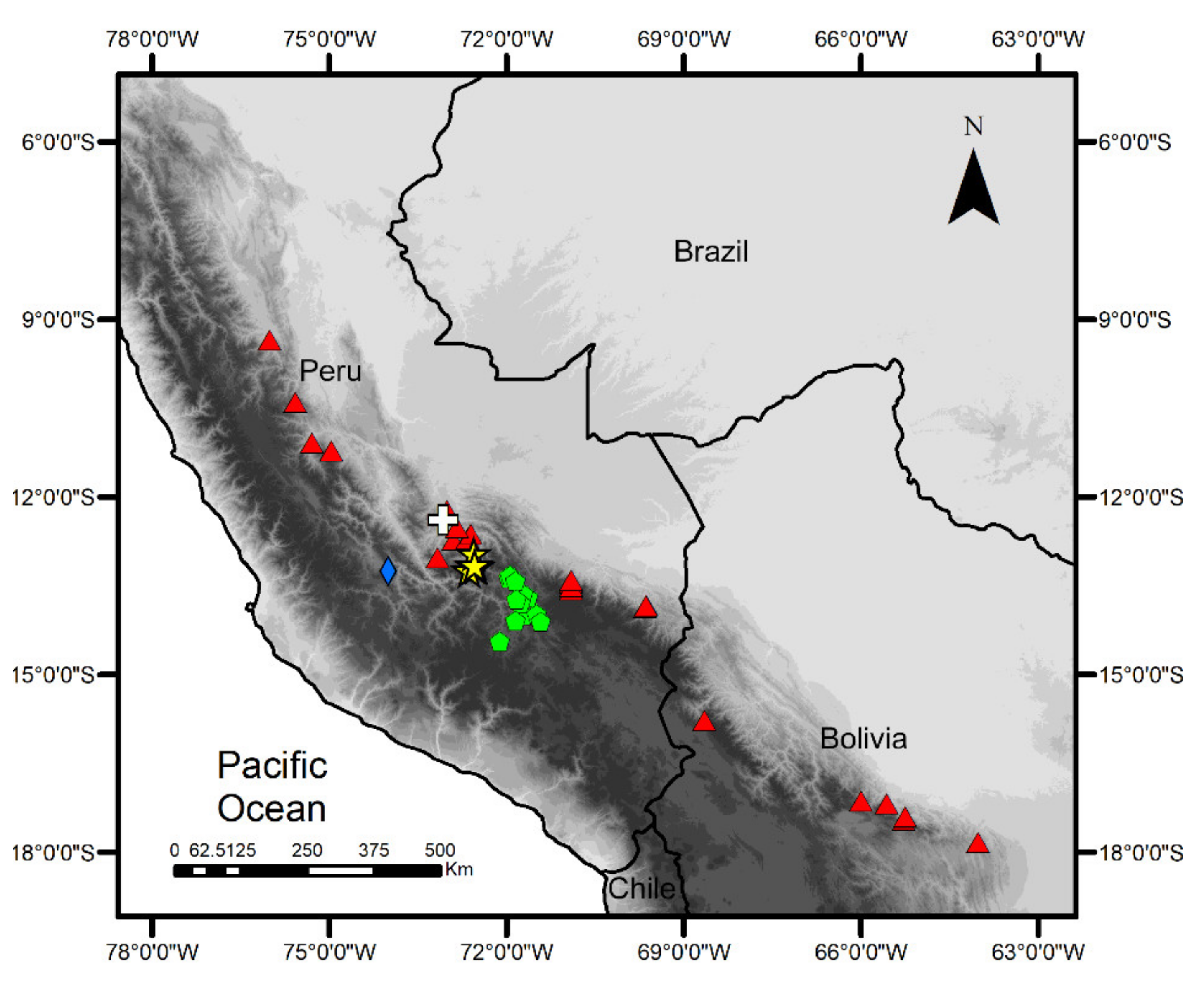
3.2. Specimens Voucher, Generic Assignment, and Species Reidentification
Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Specimens Examined
References
- Kizirian, D.A. A review of Ecuadorian Proctoporus (Squamata: Gymnophthalmidae) with descriptions of nine new species. Herpetol. Monogr. 1996, 10, 85–155. [Google Scholar] [CrossRef]
- Goicoechea, N.; Padial, J.M.; Chaparro, J.C.; Castroviejo-Fisher, S.; De la Riva, I. Molecular phylogenetics, species diversity, and biogeography of the Andean lizards of the genus Proctoporus (Squamata: Gymnophthalmidae). Mol. Phylogenet. Evol. 2012, 65, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Goicoechea, N.; Padial, J.M.; Chaparro, J.C.; Castroviejo-Fisher, S.; De La Riva, I. A taxonomic revision of Proctoporus bolivianus Werner (Squamata: Gymnophthalmidae) with the description of three new species and resurrection of Proctoporus lacertus Stejneger. Am. Mus. Novit. 2013, 3786, 1–32. [Google Scholar] [CrossRef]
- Doan, T.M.; Castoe, T.A. Phylogenetic taxonomy of the Cercosaurini (Squamata: Gymnophthalmidae), with new genera for species of Neusticurus and Proctoporus. Zool. J. Linn. Soc. 2005, 143, 405–416. [Google Scholar] [CrossRef]
- Mamani, L.; Goicoechea, N.; Chaparro, J.C. A new species of Andean lizard Proctoporus (Squamata: Gymnophthalmidae) from montane forest of the Historic Sanctuary of Machupicchu, Peru. Amphib. Reptile Conserv. 2015, 9, 1–11. [Google Scholar]
- Doan, T.M.; Castoe, T.A. Using morphological and molecular evidence to infer species boundaries within Proctoporus bolivianus Werner (Squamata: Gymnophthalmidae). Herpetologica 2003, 59, 432–449. [Google Scholar] [CrossRef]
- Uzzell, T.M. Teiid lizards of the genus Proctoporus from Bolivia and Peru. Postilla 1970, 142, 1–39. [Google Scholar]
- Diaz, M.I.; Ttito, A.; Mamani, L. A new locality and reassessment of the type series of the Machu Picchu Andean lizard Proctoporus machupicchu Mamani, Goicoechea and Chaparro, 2015 (Squamata: Gymnophthalmidae). Rev. Peru. Biol. 2019, 26, 503–508. [Google Scholar] [CrossRef]
- Carrillo, N.; Icochea, J. Lista taxonomica preliminar de los reptiles vivientes del Peru. Publ. Mus. Hist. Nat. UNMSM 1995, 49, 1–27. [Google Scholar]
- Castoe, T.A.; Doan, T.M.; Parkinson, C.L. Data partitions and complex models in Bayesian analysis: The phylogeny of gymnophthalmid lizards. Syst. Biol. 2004, 53, 448–469. [Google Scholar] [CrossRef] [Green Version]
- Torres-Carvajal, O.; Lobos, S.E.; Venegas, P.J.; Chávez, G.; Aguirre-Peñafiel, V.; Zurita, D.; Echevarría, L.Y. Phylogeny and biogeography of the most diverse clade of South American gymnophthalmid lizards (Squamata, Gymnophthalmidae, Cercosaurinae). Mol. Phylogenet. Evol. 2016, 99, 63–75. [Google Scholar] [CrossRef]
- Chávez, G.; Catenazzi, A.; Venegas, P.J. A new species of arboreal microteiid lizard of the genus Euspondylus (Gymnophtalmidae: Cercosaurinae) from the Andean slopes of central Peru with comments on Peruvian Euspondylus. Zootaxa 2017, 4350, 301–316. [Google Scholar] [CrossRef]
- Moravec, J.; Šmíd, J.; Štundl, J.; Lehr, E. Systematics of neotropical microteiid lizards (Gymnophthalmidae, cercosaurinae), with the description of a new genus and species from the andean montane forests. Zookeys 2018, 2018, 105–139. [Google Scholar] [CrossRef] [Green Version]
- Vásquez-Restrepo, J.D.; Ibáñez, R.; Sánchez-Pacheco, S.J.; Daza, J.M. Phylogeny, taxonomy and distribution of the Neotropical lizard genus Echinosaura (Squamata: Gymnophthalmidae), with the recognition of two new genera in Cercosaurinae. Zool. J. Linn. Soc. 2020, 189, 287–314. [Google Scholar] [CrossRef]
- Mamani, L.; Chaparro, J.C.; Correa, C.; Alarcón, C.; Salas, C.Y.; Catenazzi, A. A new species of Andean Gymnophthalmid Lizard (Squamata: Gymnophthalmidae) from the Peruvian Andes, and resolution of some taxonomic problems. Diversity 2020, 12, 361. [Google Scholar] [CrossRef]
- Boettger, O. Reptilien und Batrachier aus Bolivien. Zool. Anzeiger. 1891, 14, 343–347. [Google Scholar]
- Burt, C.E.; Burt, M.D. South American lizards in the collection of the American Museum of Natural History. Bull. Am. Mus. Nat. Hist. 1931, 61, 227–395. [Google Scholar]
- Werner, F. Über neue oder seltene reptilien des Naturhistorischen Museums in Hamburg: II. Eidechsen. Mitt. Nat. Mus. Hambg. 1910, 27, 1–46. [Google Scholar]
- Boulenger, G.A. Descriptions of new batrachians and reptiles from the Andes of Peru and Bolivia. Ann. Mag. Nat. Hist. 1902, 10, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Barbour, T.; Noble, G.K. Amphibians and reptiles from southern Peru collected by the Peruvian Expedition of 1914–1915 under the auspices of Yale University and the National Geographic Society. Proc. U. S. Natl. Mus. 1921, 58, 609–620. [Google Scholar] [CrossRef]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Paabo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrino, K.C.M.; Rodrigues, M.T.; Yonenaga-Yassuda, Y.; Sites, J.W. A molecular perspective on the evolution of microteiid lizards (Squamata, Gymnophthalmidae), and a new classification for the family. Biol. J. Linn. Soc. 2001, 74, 315–338. [Google Scholar] [CrossRef]
- Arévalo, E.; Sites, J.W.; Davis, S.K.; Arévalo, E. Mitochondrial DNA sequence divergence and phylogenetic relationships among eight chromosome races of the Sceloporus grammicus complex (phrynosomatidae) in central mexico. Syst. Biol. 1994, 43, 387–418. [Google Scholar] [CrossRef]
- Blair, C.; De La Cruz, F.R.M.; Ngo, A.; Lindell, J.; Lathrop, A.; Murphy, R.W. Molecular phylogenetics and taxonomy of leaf-toed geckos (Phyllodactylidae: Phyllodactylus) inhabiting the peninsula of Baja California. Zootaxa 2009, 2027, 28–42. [Google Scholar] [CrossRef]
- Saint, K.M.; Austin, C.C.; Donnellan, S.C.; Hutchinson, M.N. C-mos, a nuclear marker useful for Squamate phylogenetic analysis. Mol. Phylogenet. Evol. 1998, 10, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.61. Available online: http://www.mesquiteproject.org (accessed on 20 March 2021).
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.7.1. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 10 December 2021).
- Köhler, G. Two new species of Euspondylus (Squamata: Gymnophthalmidae) from Peru. Salamandra 2004, 39, 5–20. [Google Scholar]
- Freeman, B.G.; Scholer, M.N.; Ruiz-Gutierrez, V.; Fitzpatrick, J.W. Climate change causes upslope shifts and mountaintop extirpations in a tropical bird community. Proc. Natl. Acad. Sci. USA 2018, 115, 11982–11987. [Google Scholar] [CrossRef]
- Marris, E. The escalator effect. Nat. Clim. Change 2007, 1, 94–96. [Google Scholar] [CrossRef]
- Urban, M.C. Escalator to extinction. Proc. Natl. Acad. Sci. USA 2018, 115, 11871–11873. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Parmesan, C.; Williams-Anderson, A.; Moskwik, M.; Mikheyev, A.S.; Singer, M.C. Endangered Quino checkerspot butterfly and climate change: Short-term success but long-term vulnerability? J. Insect Conserv. 2015, 19, 185–204. [Google Scholar] [CrossRef] [Green Version]
- Barros, A.; Monz, C.; Pickering, C. Is tourism damaging ecosystems in the Andes? Current knowledge and an agenda for future research. Ambio 2015, 44, 82–98. [Google Scholar] [CrossRef]
- Peña-Candia, L.Z.; Baca-Zans, Y.R.; Costa, J.F. Efecto de las actividades antrópicas en la red de caminos Inka, santuario histórico de Machupicchu, Cusco, Perú. Rev. Interam. Ambient. Tur. 2019, 15, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Bonacic, C.; Almuna, R.; Ibarra, J.T. Biodiversity conservation requires management of feral domestic animals. Trends Ecol. Evol. 2019, 34, 683–686. [Google Scholar] [CrossRef]
- Cambray, J.A. The global impact of alien trout species—A review; with reference to their impact in south africa. Afr. J. Aquat. Sci. 2003, 28, 61–67. [Google Scholar] [CrossRef]
- Coleman, J.L.; Ford, N.B.; Herriman, K. A road survey of amphibians and reptiles in a bottomland hardwood forest. Southeast. Nat. 2008, 7, 339–348. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Catenazzi, A.; Lehr, E.; Rodriguez, L.O.; Vredenburg, V.T. Batrachochytrium dendrobatidis and the collapse of anuran species richness and abundance in the upper Manu National Park, Southeastern Peru. Conserv. Biol. 2010, 25, 382–391. [Google Scholar] [CrossRef]
- Sordello, R.; Ratel, O.; de Lachapelle, F.F.; Leger, C.; Dambry, A.; Vanpeene, S. Evidence of the impact of noise pollution on biodiversity: A systematic map. Environ. Evid. 2020, 9, 1–27. [Google Scholar] [CrossRef]
- Colino-Rabanal, V.J.; Lizana, M. Herpetofauna and roads: A review. Basic Appl. Herpetol. 2012, 26, 5–31. [Google Scholar] [CrossRef]
- Lucas, P.S.; de Carvalho, R.G.; Grilo, C. Railway disturbances on wildlife: Types, effects, and mitigation measures. In Railway Ecology; Springer: Cham, Switzerland, 2017; pp. 81–99. ISBN 9783319574967. [Google Scholar]
- Catenazzi, A.; Mamani, L.; Lehr, E.; von May, R. A new genus of terrestrial-breeding frogs (Holoadeninae, Strabomantidae, terrarana) from Southern Peru. Diversity 2020, 12, 184. [Google Scholar] [CrossRef]
- Hedges, B.S.; Duellman, W.E.; Heinicke, M.P. New World Direct-developing frogs (Anura: Terrarana): Molecular phylogeny, classification, biogeography, and conservation. Zootaxa 2008, 1737, 1–182. [Google Scholar] [CrossRef] [Green Version]
- De La Riva, I.; Chaparro, J.C.; Castroviejo-Fisher, S.; Padial, J.M. Underestimated anuran radiations in the high andes: Five new species and a new genus of holoadeninae, and their phylogenetic relationships (anura: Craugastoridae). Zool. J. Linn. Soc. 2018, 182, 129–172. [Google Scholar] [CrossRef]
- Sánchez-Pacheco, S.J.; Torres-Carvajal, O.; Aguirre-Peñafiel, V.; Nunes, P.M.S.; Verrastro, L.; Rivas, G.A.; Rodrigues, M.T.; Grant, T.; Murphy, R.W. Phylogeny of Riama (Squamata: Gymnophthalmidae), impact of phenotypic evidence on molecular datasets, and the origin of the Sierra Nevada de Santa Marta endemic fauna. Cladistics 2018, 34, 260–291. [Google Scholar] [CrossRef]
- Doan, T.M.; Markham, S.; Gregory, A.; Broadwater, C.O.; Floyd, A.; Goldberg, M.J.; Calder, B. Hot Lizards: Testing the Tolerance to Climate Warming of Thermoconformers in the Andes (Squamata: Gymnophthalmidae). Ichthyol. Herpetol. 2022, 110, 87–95. [Google Scholar] [CrossRef]
- Rapp, J.M.; Silman, M.R. Diurnal, seasonal, and altitudinal trends in microclimate across a tropical montane cloud forest. Clim. Res. 2012, 55, 17–32. [Google Scholar] [CrossRef]
- Dirksen, L.; De la Riva, I. The lizards and amphisbaenians of Bolivia (Reptilia, Squamata): Checklist, localities, and bibliography. Graellsia 1999, 55, 199–215. [Google Scholar] [CrossRef]
- De la Riva, I.; Köhler, J.; Lötters, S.; Reichle, S. Ten years of research on Bolivian amphibians: Updated checklist, distribution, taxonomic problems, literature and iconography. Rev. Esp. Herpetol. 2000, 14, 19–164. [Google Scholar]
- Fuentes, A. Una introducción a la vegetación de la región de Madidi. Ecol. Boliv. Rev. Inst. Ecol. 2005, 40, 1–31. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality | Coordinates | Voucher | 12S | 16S | ND4 | Cytb | c-mos |
|---|---|---|---|---|---|---|---|---|
| Proctoporus guentheri | Alto Matoriato, La Convención, Cusco. | 12°30′20″ S/72°49′59″ W | MUBI 10278 | OM893811 | OM893806 | OM885354 | OM885350 | OM885358 |
| Proctoporus guentheri | Tucantinas, La Convención, Cusco. | 12°43′21″ S/72°53′56″ W | MUBI 12687 | OM893812 | OM893807 | - | OM885351 | OM885359 |
| Proctoporus katerynae sp. nov. | Monte Carmelo, La Convención, Cusco. | 12°22′58″ S/73°4′20″ W | MUBI 10482 | OM893813 | OM893808 | OM885355 | - | OM885360 |
| Proctoporus optimus sp. nov. | Mesa Pelada, La Convención, Cusco. | 12°56′21″ S/72°35′56″ W | MUBI 2915 | OM893814 | OM893809 | OM885356 | OM885352 | OM885361 |
| Proctoporus optimus sp. nov. | Mesa Pelada, La Convención, Cusco. | 12°56′21″ S/72°35′56″ W | MUBI 2984 | OM893815 | OM893810 | OM885357 | OM885353 | OM885362 |
| Gene | Primer | Primers Sequence (5′–3′) | PCR Cycle | Reference |
|---|---|---|---|---|
| 12S | 12S1L | CAAACTGGGATTAGATACCCCACTAT | 94 °C/3 min; 33 × (95 °C/30 s, 57 °C/30 s, 72 °C/90 s); 72 °C/10 min | [21] |
| 12S2H | AGGGTGACGGGCGGTGTGT | |||
| 16S | 16sF.0 | CTGTTTACCAAAAACATMRCCTYTAGC | 96 °C/3 min; 40 × (95 °C/30 s, 51 °C/60 s, 72 °C/60 s); 72 °C/10 min | [21,22] |
| 16sR.0 | TAGATAGAAACCGACCTGGATT | |||
| ND4 | ND412931L | CTACCAAAAGCTCATGTAGAAGC | 96 °C/3 min; 40 × (95 °C/30 s, 52 °C/60, 72 °C/60 s); 72 °C/10 min | [23,24] |
| ND413824H | CATTACTTTTACTTGGATTTGCACCA | |||
| Cytb | L14841 | AAAAAGCTTCCATCCAACATCTCAGCATGATGAAA | 94 °C/5 min; 30 × (94 °C/60 s, 50 °C/60 s, 72 °C/60); 72 °C/10 min | [21] |
| H15149 | AAACTGCAGCCCCTCAGAATGATATTTGTCCTCA | |||
| c-mos | G73 | GCGGTAAAGCAGGTGAAGAAA | 96 °C/3 min; 35 × (95 °C/25 s, 52 °C/60 s, 72 °C/120 s), 72 °C/10 min | [25] |
| G74 | TGAGCATCCAAAGTCTCCAATC |
| Measurements (mm) | Proctoporus optimus sp. nov. | Proctoporus katerynae sp. nov. | |||
|---|---|---|---|---|---|
| Males (n = 4) | Females (n = 3) | MUBI 10482 | MUBI 10492 | MUBI 10493 | |
| Adult Female | Juvenile Male | Juvenile Female | |||
| Snout vent length | 44.7 (43.8–46.5) ± 3.3 | 44.9 (43–45.9) ±1.7 | 38.2 | 27.4 | 19.0 |
| Tail length | 62.9 (56–70.8) ± 8.1 | 67.6–80.3 | 58.5 | lost | 11.6 |
| Length between arm and leg | 24.5 (23.8–26.4) ± 1.9 | 24.0 (22–25.5) ± 1.8 | 21.3 | 14.0 | 8.3 |
| Head length to ear | 9 (8.4–10.4) ± 0.8 | 8.4 (8–8.7) ±0.4 | 7.4 | 5.2 | 4.8 |
| Head width | 6.53 (5.6–7.6) ± 0.7 | 6.5 (6–6.1) ± 0.1 | 4.8 | 3.8 | 3.1 |
| Frontal (mm) | 1.87 (1.8–2.1) ± 0.1 | 1.73 (1.7–1.8) ± 0.1 | 1.5 | 1.2 | 1.1 |
| Frontonasal (mm) | 2.1 (1.9–2.5) ± 0.2 | 2.07 (1.9–2.2) ± 0.2 | 1.8 | 1.4 | 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamani, L.; Cruz, R.; Mallqui, S.; Catenazzi, A. Molecular Phylogenetics and Comparative Examination of Voucher Museums Reveal Two New Species of Gymnophthalmid Lizards (Squamata, Gymnophthalmidae) from the Peruvian Andes, with Comments on Proctoporus guentheri (Boettger, 1891). Diversity 2022, 14, 215. https://doi.org/10.3390/d14030215
Mamani L, Cruz R, Mallqui S, Catenazzi A. Molecular Phylogenetics and Comparative Examination of Voucher Museums Reveal Two New Species of Gymnophthalmid Lizards (Squamata, Gymnophthalmidae) from the Peruvian Andes, with Comments on Proctoporus guentheri (Boettger, 1891). Diversity. 2022; 14(3):215. https://doi.org/10.3390/d14030215
Chicago/Turabian StyleMamani, Luis, Roxana Cruz, Sergio Mallqui, and Alessandro Catenazzi. 2022. "Molecular Phylogenetics and Comparative Examination of Voucher Museums Reveal Two New Species of Gymnophthalmid Lizards (Squamata, Gymnophthalmidae) from the Peruvian Andes, with Comments on Proctoporus guentheri (Boettger, 1891)" Diversity 14, no. 3: 215. https://doi.org/10.3390/d14030215