
Do Mixed Pinus yunnanensis Plantations Improve Soil’s Physicochemical Properties and Enzyme Activities?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area Overview
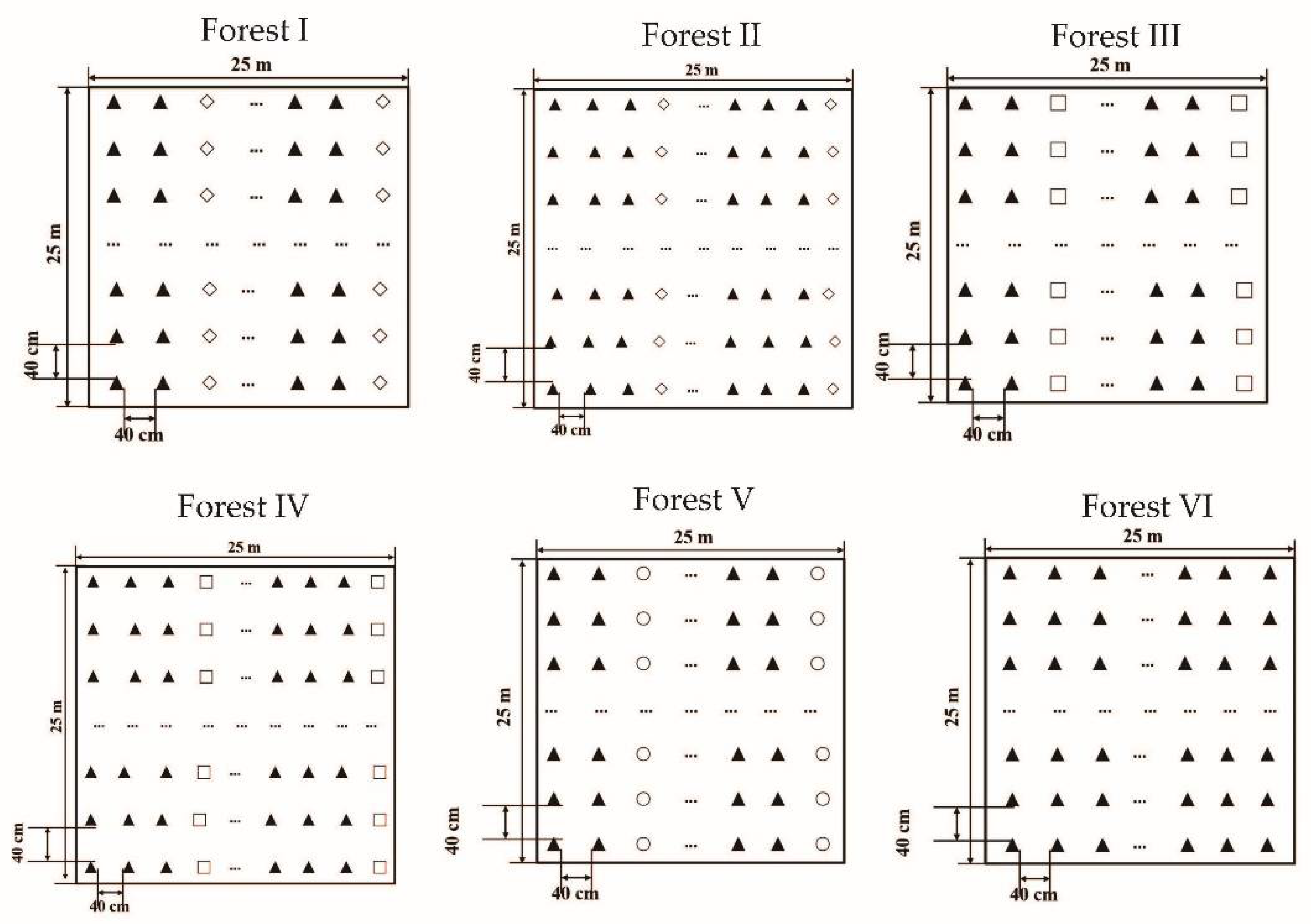
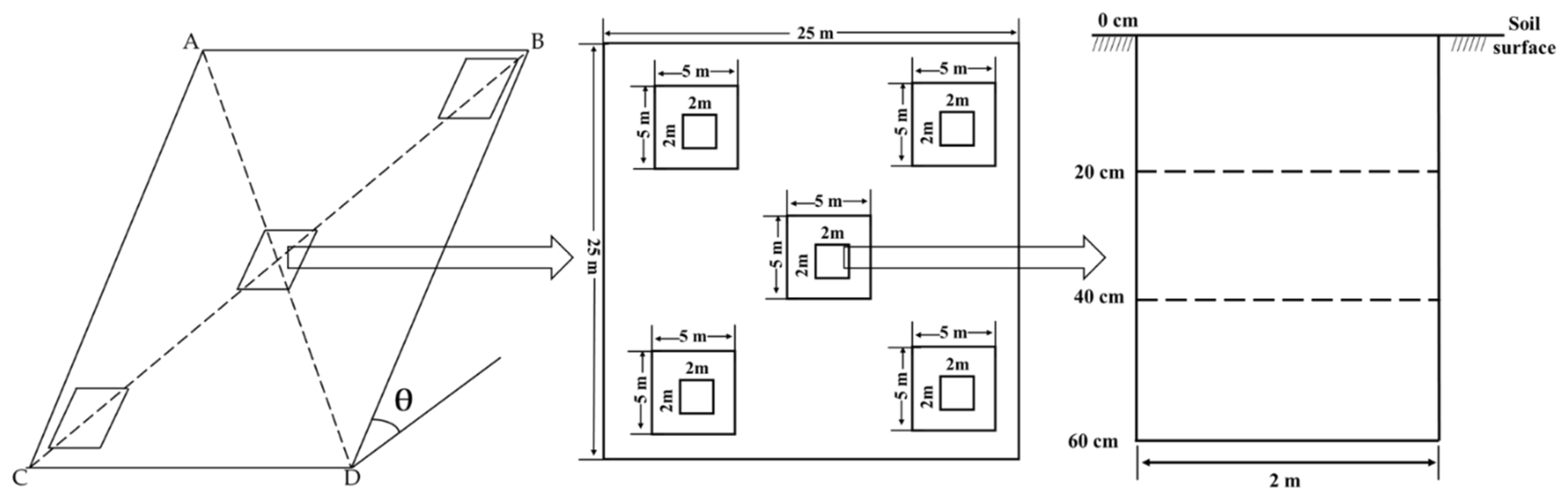
2.2. Sample Plot Selection and Soil Sample Collection
2.3. Measurement Items and Methods
2.4. Statistical Analysis
3. Results
3.1. Effects of Different Forest Types of P. yunnanensis on Soil’s Physicochemical Properties
3.1.1. Soil’s Physical Properties
3.1.2. Soil’s Chemical Properties
3.2. Effect of Different Forest Types of P. yunnanensis on Soil Enzyme Activity
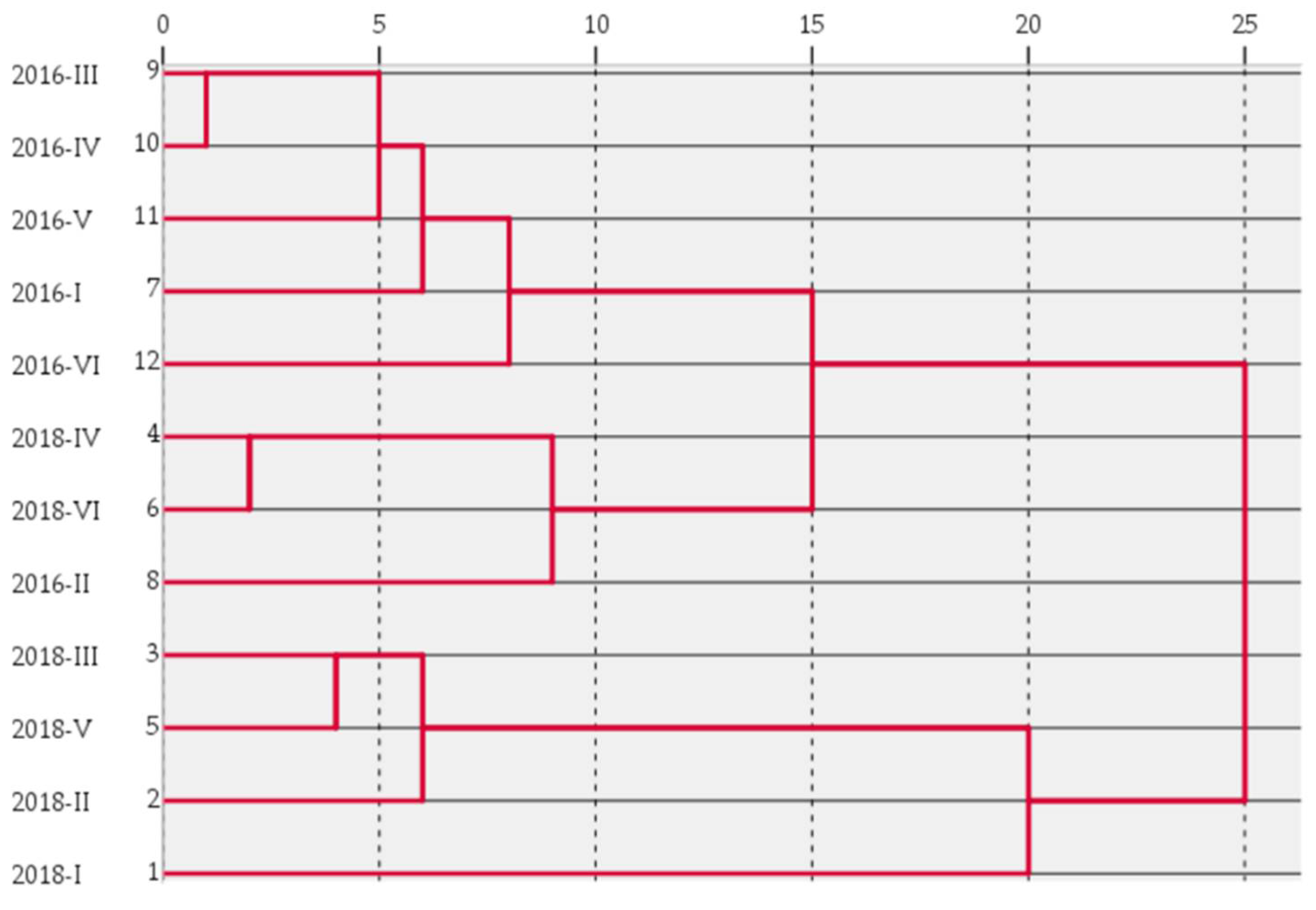
3.3. Principal Component Analysis and Cluster Analysis of Soil Fertility in Different Forest Types
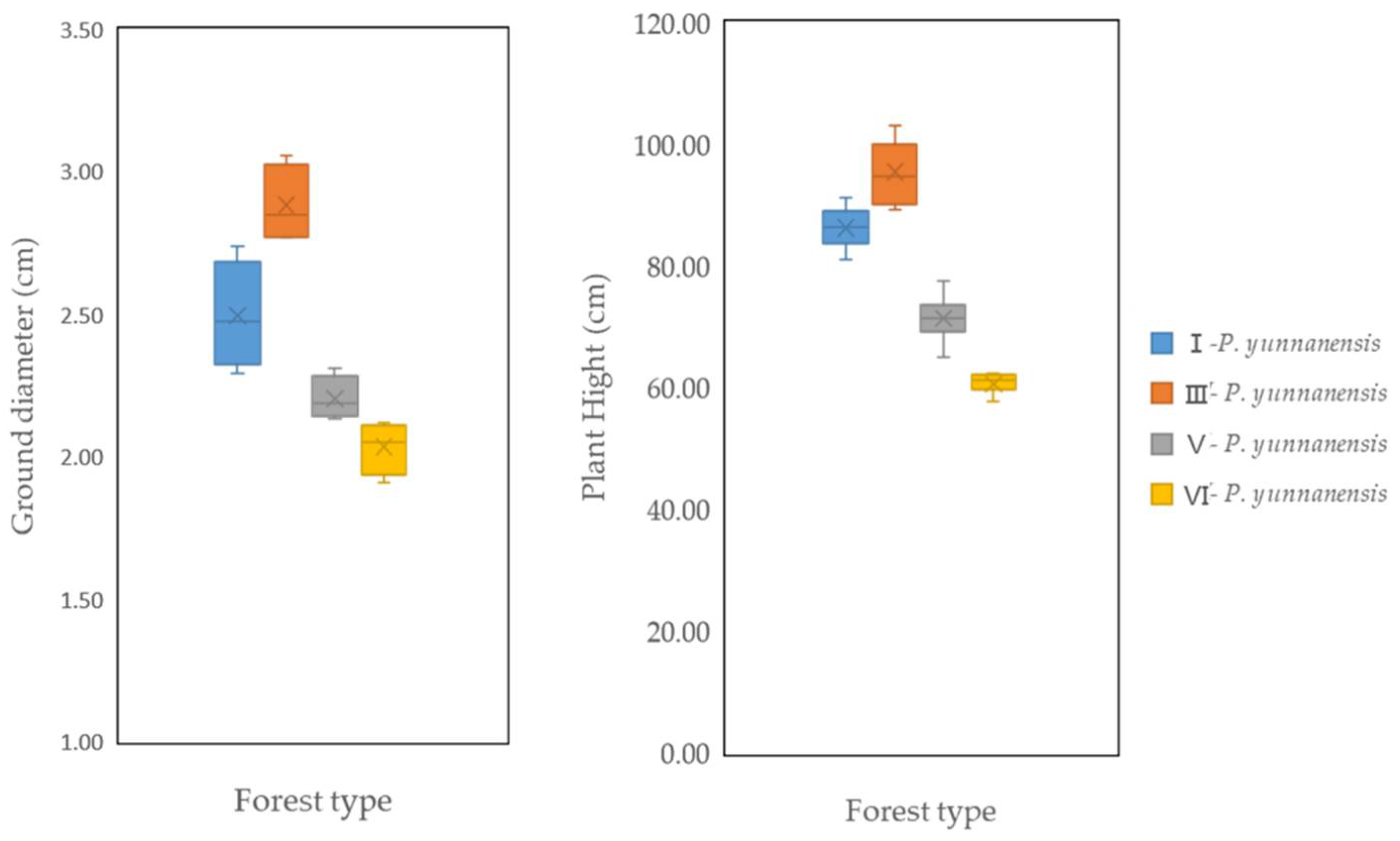
3.4. Growth Status of Various Tree Species after Two Years of Afforestation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, X.; Li, S.; Su, J.; Liu, W.; Lang, X. The relationship between species richness and ecosystem multifunctionality in the Pinus yunnanensis natural secondary forest. Biodivers. Sci. 2017, 25, 1182–1191. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Woeste, K.; Cai, N.; Kang, X.; Li, G.; Chen, S.; Duan, A. Variation in needle and cone traits in natural populations of Pinus yunnanensis. J. For. Res. 2016, 27, 41–49. [Google Scholar] [CrossRef]
- Rothe, A.; Binkley, D. Nutritional interactions in mixed species forests: A synthesis. Can. J. For. Res. 2001, 31, 1855–1870. [Google Scholar] [CrossRef]
- Castagneyrol, B.; Jactel, H.; Vacher, C.; Brockerhoff, E.G.; Koricheva, J. Effects of plant phylogenetic diversity on herbivory depend on herbivore specialization. J. Appl. Ecol. 2014, 51, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhao, X.; Gadow, K.V. Analyzing selective harvest events in three large forest observational studies in North Eastern China. For. Ecol. Manag. 2014, 316, 100–109. [Google Scholar] [CrossRef]
- Huang, X.; Liu, S.; You, Y.; Wen, Y.; Wang, H.; Wang, J. Microbial community and associated enzymes activity influence soil carbon chemical composition in Eucalyptus urophylla plantation with mixing N2-fixing species in subtropical China. Plant Soil 2017, 414, 199–212. [Google Scholar] [CrossRef]
- Zhang, P.; Pang, S.J.; Yang, B.G.; Liu, S.L.; Jia, H.Y.; Chen, J.B.; Guo, D.Q. Effects of different mixed models on Eucalyptus growth, litter and soil nutrients. J. Northwest A&F Univ. 2021, 49, 31–37. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Spehn, E.M.; Joshi, J.; Schmid, B.; Diemer, M.; Körner, C. Aboveground resource use increases with plant species richness in experimental grassland ecosystems. Funct. Ecol. 2000, 14, 326–337. [Google Scholar]
- Kowalchuk, G.A.; Buma, D.S.; de Boer, W.; Klinkhamer, P.G.L.; Van Veen, J.A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie Van Leeuwenhoek 2002, 81, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Heemsbergen, D.A.; Berg, M.P.; Loreau, M.; van Hal, J.R.; Faber, J.H.; Verhoef, H.A. Biodiversity Effects on Soil Processes Explained by Interspecific Functional Dissimilarity. Science 2004, 306, 1019–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhauer, N.; Reich, P.B.; Scheu, S. Increasing plant diversity effects on productivity with time due to delayed soil biota effects on plants. Basic Appl. Ecol. 2012, 13, 571–578. [Google Scholar] [CrossRef]
- Lavelle, P.; Lattaud, C.; Trigo, D.; Barois, I. Mutualism and biodiversity in soils. Plant Soil 1995, 170, 23–33. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, C.; Long, M. Variation of soil nutrients and bacterial community diversity of different land utilization types in Yangtze River Basin, Chongqing Municipality. PeerJ 2020, 8, e9386. [Google Scholar] [CrossRef] [PubMed]
- Schoenholtz, S.H.; Miegroet, H.V.; Burger, J.A. A review of chemical and physical properties as indicators of forest soil quality: Challenges and opportunities. For. Ecol. Manag. 2000, 138, 335–356. [Google Scholar] [CrossRef]
- Hayicho, H.; Alemu, M.; Kedir, H. Assessing the Effects of Land-Use and Land Cover Change and Topography on Soil Fertility in Melka Wakena Catchment of Sub-Upper Wabe-Shebelle Watershed, South Eastern Ethiopia. J. Environ. Prot. 2019, 10, 672–693. [Google Scholar] [CrossRef] [Green Version]
- Dise, N.B.; Matzner, E.; Forsius, M. Evaluation of organic horizon C:N ratio as an indicator of nitrate leaching in conifer forests across Europe. Environ. Pollut. 1998, 102, 453–456. [Google Scholar] [CrossRef]
- Salam, A.K.; Katayama, A.; Kimura, M. Activities of some soil enzymes in different land use systems after deforestation in hilly areas of West Lampung, South Sumatra, Indonesia. Soil Sci. Plant Nutr. 1998, 44, 93–103. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Jin, Y.H.; Gu, X.N.; Jia, W.; Tao, Y.; He, H.S.; Wang, A.L.; Liu, Y.X.; Zhu, L.P. Vegetation change in relation to soil microbes, enzyme activity and soil fertility in the tundra of Changbai Mountain. Chin. J. Ecol. 2017, 36, 3086–3093. [Google Scholar] [CrossRef]
- Karlen, D.L.; Ditzler, C.A.; Andrews, S.S. Soil quality: Why and how? Geoderma 2003, 114, 145–156. [Google Scholar] [CrossRef]
- Smaling, E.M.A.; Stoorvogel, J.J.; Windmeijer, P.N. Calculating soil nutrient balances in Africa at different scales. Fertil. Res. 1993, 35, 237–250. [Google Scholar] [CrossRef]
- Ling, Q.; Zhao, X.; Wu, P.; Gao, X.; Sun, W. Effect of the fodder species canola (Brassica napus L.) and daylily (Hemerocallis fulva L.) on soil physical properties and soil water content in a rainfed orchard on the semiarid Loess Plateau, China. Plant Soil 2020, 453, 209–228. [Google Scholar] [CrossRef]
- Shiau, Y.-J.; Chiu, C.-Y. Changes in Soil Biochemical Properties in a Cedar Plantation Invaded by Moso Bamboo. Forests 2017, 8, 222. [Google Scholar] [CrossRef]
- Li, C.; Yan, L.; Jian, M. Spatial heterogeneity of soil chemical properties at fine scales induced by Haloxylon ammodendron (Chenopodiaceae) plants in a sandy desert. Ecol. Res. 2011, 26, 385–394. [Google Scholar] [CrossRef]
- Zhu, T.; Zhang, J.; Meng, T.; Zhang, Y.; Yang, J.; Müller, C.; Cai, Z. Tea plantation destroys soil retention of NO3− and increases N2O emissions in subtropical China. Soil Biol. Biochem. 2014, 73, 106–114. [Google Scholar] [CrossRef]
- Qin, F.; Lirong, S.U.; Zeng, C.; Qin, L.; Tieguang, H.E.; Yuefeng, Y.U.; Nan, W.E.I.; Yuanqing, M.E.N.G.; Aina, W.E.I.; Jinshan, W.E.I.; et al. Improving Soil Fertility with Different Planting Patterns in Rocky Desertification Areas. Agric. Biotechnol. 2020, 9, 119–124. [Google Scholar]
- Ziadi, N.; Whalen, J.K.; Messiga, A.J.; Morel, C. Assessment and Modeling of Soil Available Phosphorus in Sustainable Cropping Systems. Adv. Agron. 2013, 122, 85–126. [Google Scholar] [CrossRef]
- Gopal, M.; Gupta, A.; Arunachalam, V.; Magu, S. Impact of azadirachtin, an insecticidal allelochemical from neem on soil microflora, enzyme and respiratory activities. Bioresour. Technol. 2007, 98, 3154–3158. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-M.; Cao, W.-Q.; Chen, X.-X.; Yu, B.-G.; Lang, M.; Chen, X.-P.; Zou, C.-Q. The responses of soil enzyme activities, microbial biomass and microbial community structure to nine years of varied zinc application rates. Sci. Total Environ. 2020, 737, 140245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, G.; Xue, S.; Song, Z. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Farrer, E.C.; Goldberg, D.E. Litter drives ecosystem and plant community changes in cattail invasion. Ecol. Appl. 2009, 19, 398–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Gao, P.; Li, P.; Zhang, J.C.; Dong, J.W.; Xu, J.W.; Tun, X.J. Effects of soil microbial community on the litter decomposition in mixed Quercus acutissima Carruth. and Robinia pseudoacacia L. forest. Acta Ecol. Sin. 2021, 41, 2315–2325. [Google Scholar]
- Zhou, L.; Sun, Y.; Saeed, S.; Zhang, B.; Luo, M. The difference of soil properties between pure and mixed Chinese fir (Cunninghamia lanceolata) plantations depends on tree species. Glob. Ecol. Conserv. 2020, 22, e01009. [Google Scholar] [CrossRef]
- Li, D.S.; Zhang, P.; Zhang, S.L.; Yin, J.D.; Lu, F.D. A Study on Water Conservation Capacity of Forest Soil in Huangqian Reservoir Area. J. Nanjing For. Univ. 2004, 47, 25. [Google Scholar]
- Ming-Liang, L.U.; Chen, Y.F.; Fang, Z.Z.; He, F. Soil physical and chemical properties of different types of ecological forest stands in Kecheng District. J. Zhejiang For. Sci. Technol. 2010, 30, 70–72. [Google Scholar] [CrossRef]
- Chen, X.F.; Lin, G.W.; Hong, T.; Chen, J.Z.; Su, S.C.; Hong, W.; Wu, C.Z.; Lin, H. Soil Physical and Chemical Properties of Cunninghamia lanceolata-Aleurites Montana Mixed Forestand Cunninghamia lanceolata Pure Forest with Different Age. Chin. J. Trop. Crop. 2017, 38, 1660–1665. [Google Scholar]
- Gray, A.N.; Spies, T.A.; Easter, M.J. Microclimatic and soil moisture responses to gap formation in coastal Douglas-fir forests. Can. J. For. Res. 2002, 32, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Scharenbroch, B.; Bockheim, J. Pedodiversity in an old-growth northern hardwood forest in the Huron Mountains, Upper Peninsula, Michigan. Can. J. For. Res. 2007, 37, 1106–1117. [Google Scholar] [CrossRef]
- Zhang, K.; Zheng, H.; Chen, F.L.; Ouyang, Z.Y.; Wang, Y.; Wu, Y.F.; Lan, J.; Fu, M.; Xiang, X.W. Changes in soil quality after converting Pinus to Eucalyptus plantations in southern China. Solid Earth 2015, 6, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Yesilonis, I.; Szlavecz, K.; Pouyat, R.; Whigham, D.; Xia, L. Historical land use and stand age effects on forest soil properties in the Mid-Atlantic US. For. Ecol. Manag. 2016, 370, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, W.; Wu, Y.; Hao, Z.; Zhang, Q.; Bu, Q.; Gao, X. Combined impacts of land use and soil property changes on soil erosion in a mollisol area under long-term agricultural development. Sci. Total Environ. 2018, 613–614, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Pan, K.; Xu, Z.; Blumfield, T.; Totua, S.; Lu, M. In situ mineral 15N dynamics and fate of added 15NH4 + in hoop pine plantation and adjacent native forest in subtropical Australia. J. Soils Sediments 2008, 8, 398–405. [Google Scholar] [CrossRef]
- Zhou, Y.D.; Wu, J.S.; Zhao, S.W.; Guo, S.L.; Lu, P. Change of soil organic matter and water holding ability during vegetation succession in Ziwuling region. Acta Bot. Boreali-Occident. Sin. 2003, 06, 895–900. [Google Scholar]
- Dawud, S.M.; Raulund-Rasmussen, K.; Domisch, T.; Finér, L.; Jaroszewicz, B.; Vesterdal, L. Is tree species diversity or species identity the more important driver of soil carbon stocks, C/N ratio, and pH? Ecosystems 2016, 19, 645–660. [Google Scholar] [CrossRef] [Green Version]
- Stewart, W.D.P. A Quantitative Study of Fixation and Transfer of Nitrogen in Alnus. J. Exp. Bot. 1962, 13, 250–256. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L. On the success and failure of mixed-species tree plantations: Lessons learned from a model system of Eucalyptus globulus and Acacia mearnsii. For. Ecol. Manag. 2005, 209, 147–155. [Google Scholar] [CrossRef]
- Liao, Z.Y.; Hui, Y.; Wang, S.J.; Chen, P. Leaf litter decomposition and nutrient return characteristics of Pinus yunnanensis at different forest ages. Ecol. Environ. Sci. 2018, 27, 1981–1986. [Google Scholar] [CrossRef]
- Liu, W.D.; Su, J.R.; Li, S.F.; Zhang, Z.J.; Li, Z.J. Stoichiometry study of C,N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Acta Ecol. Sin. 2010, 30, 6581–6590. [Google Scholar]
- Wang, Z.Q.; Wang, Q.C. The spatial heterogeneity of soil physical properties in forests. Acta Ecol. Sin. 2000, 20, 945–950. [Google Scholar]
- Wei, Q.; Zhang, G.; Lei, L.; Chai, C. Water conservation function of litter and soil layer under main forest types in Xinglong Mountain of Gansu. J. Nanjing For. Univ. Nat. Sci. Ed. 2013, 56, 78. [Google Scholar]
- Yang, Y.; Yang, Y.; Geng, Y.; Huang, G.; Cui, X.; Hou, M. Effects of Different Land Types on Soil Enzyme Activity in the Qinghai Lake Region. Wetlands 2018, 38, 711–721. [Google Scholar] [CrossRef]
- Liu, Z.H.; Li, G.; Chen, R.; Wu, M.; Tong, Q.; Tong, F.P. Soil chemical properties of different mixed forests in Hunan province. J. Cent. South Univ. For. Technol. 2018, 38, 7. [Google Scholar] [CrossRef]
- Zou, J.; Li, Y.Y.; Zhang, Y.W.; Yu, L.F. Soil sucrase, phosphatase and catalase enzyme activity change of degraded Karst Vegetation restoration research. Guangdong Agric. Sci. 2013, 40, 88–91. [Google Scholar] [CrossRef]
- Mo, J.; Yan, W.D.; Liu, S.G.; Wu, X.h. Soil enzyme activities and their relations with soil fertility in Camellia oleifera peanut intercropping. J. Cent. South Univ. For. Technol. 2017, 37, 89–95. [Google Scholar] [CrossRef]
- Zhou, Z.Y. Soil enzyme activities under different vegetation types in Beipan River Karst Gorge District. Sci. Silvae Sin. 2010, 46, 136–141. [Google Scholar] [CrossRef]
- Wei, L.; Hao, H.L.; Wu, W.; Wei, Q.K.; Chen, Y.X.; Thies, J.E. Transgenic Bt rice does not affect enzyme activities and microbial composition in the rhizosphere during crop development. Soil Biol. Biochem. 2008, 40, 475–486. [Google Scholar]
- Hoogmoed, M.; Cunningham, S.; Baker, P.; Beringer, J.; Cavagnaro, T. N-fixing trees in restoration plantings: Effects on nitrogen supply and soil microbial communities. Soil Biol. Biochem. 2014, 77, 203–212. [Google Scholar] [CrossRef]
- Wang, F.; Li, Z.; Xia, H.; Zou, B.; Li, N.; Liu, J.; Zhu, W. Effects of nitrogen-fixing and non-nitrogen-fixing tree species on soil properties and nitrogen transformation during forest restoration in southern China. Soil Sci. Plant Nutr. 2010, 56, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Qu, G.H.; Guo, J.X. The relationship between different plant communities and soil characteristics in Songnen grassland. Acta Prataculturae Sin. 2003, 12, 18–22. [Google Scholar]
- Chang, C.; Xie, Z.Q.; Xiong, G.M.; Zhao, C.M.; Shen, G.Z.; Lai, J.S.; Xu, X.W. Characteristics of soil nutrients of different vegetation types in the Three Gorges reservoir area. Acta Ecol. Sin. 2009, 29, 5978–5985. [Google Scholar]
- Trasar-Cepeda, C.; Leirós, C.; Gil-Sotres, F.; Seoane, S. Towards a biochemical quality index for soils: An expression relating several biological and biochemical properties. Biol. Fertil. Soils 1998, 26, 100–106. [Google Scholar] [CrossRef]
- Wang, W.; Linglin, X.U.; Zeng, C.; Tong, C.; Zhang, L. Carbon, nitrogen and phosphorus ecological stoichiometric ratios among live plant-litter-soil systems in estuarine wetland. Acta Ecol. Sin. 2011, 31, 7119–7124. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Stands | Altitude (m) | Slope (°) | Latitude | Longitude |
|---|---|---|---|---|
| I | 1890 | 3 | N 25°25′13.46″ | E 100°29′41.41″ |
| II | 1890 | 5 | N 25°25′15.17″ | E 100°29′39.50″ |
| III | 1879 | 5 | N 25°25′14.17″ | E 100°29′34.59″ |
| IV | 1895 | 2 | N 25°25′12.13″ | E 100°29′52.64″ |
| V | 1895 | 2 | N 25°25′12.13″ | E 100°29′52.64″ |
| VI | 1879 | 2 | N 25°24′56.76″ | E 100°29′36.00″ |
| Year | Depth (cm) | I | II | III | IV | V | VI |
|---|---|---|---|---|---|---|---|
| BD (g/cm3) | |||||||
| 2016 | 0–20 | 1.29 ± 0.04 Aa | 1.26 ± 0.06 Aa | 1.25 ± 0.04 Ba | 1.26 ± 0.02 Ba | 1.29 ± 0.04 Aa | 1.28 ± 0.03 Ba |
| 20–40 | 1.37 ± 0.05 ABa | 1.37 ± 0.07 ABa | 1.40 ± 0.08 Aa | 1.39 ± 0.02 Aa | 1.38 ± 0.07 Aa | 1.39 ± 0.04 Aa | |
| 40–60 | 1.4 ± 0.05 Aa | 1.41 ± 0.03 Aa | 1.42 ± 0.04 Aa | 1.42 ± 0.04 Aa | 1.42 ± 0.06 Aa | 1.41 ± 0.02 Aa | |
| 2018 | 0–20 | 1.14 ± 0.07 Aab | 1.24 ± 0.04 Aab | 1.10 ± 0.02 Aab | 1.22 ± 0.02 Aab | 1.07 ± 0.15 Ab | 1.27 ± 0.03 Ba |
| 20–40 | 1.19 ± 0.07 Aab | 1.28 ± 0.03 Aab | 1.20 ± 0.08 ABab | 1.31 ± 0.07 Aab | 1.13 ± 0.12 Ab | 1.36 ± 0.01 Aa | |
| 40–60 | 1.39 ± 0.18 Aa | 1.34 ± 0.05 Aa | 1.30 ± 0.06 Aa | 1.35 ± 0.07 Aa | 1.25 ± 0.05 Aa | 1.39 ± 0.04 Aa | |
| TOP (%) | |||||||
| 2016 | 0–20 | 44.53 ± 2.30 Aa | 46.88 ± 5.13 Aa | 46.86 ± 3.01 Aa | 45.88 ± 9.36 Aa | 45.36 ± 6.17 Aa | 44.61 ± 0.46 Ca |
| 20–40 | 41.03 ± 1.18 ABa | 41.05 ± 7.02 Aa | 41.97 ± 2.00 Aba | 41.25 ± 4.76 Aa | 41.55 ± 1.89 Aa | 41.69 ± 1.47 Aa | |
| 40–60 | 37.51 ± 1.32 Bb | 37.65 ± 4.15 Ab | 37.43 ± 2.47 Bb | 37.21 ± 1.91 Ab | 37.21 ± 5.99 Ab | 37.75 ± 0.73 Ba | |
| 2018 | 0–20 | 56.07 ± 0.57 Aab | 51.75 ± 1.61 Abc | 58.33 ± 4.50 Aab | 53.46 ± 3.08 Abc | 62.96 ± 2.79 Aa | 47.88 ± 1.46 Ac |
| 20–40 | 52.89 ± 3.48 ABab | 48.83 ± 2.41 ABbc | 55.59 ± 1.84 Aab | 50.63 ± 1.87 ABbc | 59.2 ± 3.90 Aba | 43.61 ± 1.32 Bc | |
| 40–60 | 48.90 ± 0.70 Bab | 46.18 ± 1.04 Bb | 51.16 ± 1.92 Aa | 47.75 ± 1.38 Bab | 52.27 ± 2.71 Ba | 38.28 ± 1.76 Cc | |
| MC (%) | |||||||
| 2016 | 0–20 | 27.18 ± 1.01 Aa | 26.63 ± 6.58 Aa | 27.07 ± 11.31 Aa | 27.41 ± 4.49 Aa | 28.14 ± 6.36 Aa | 27.99 ± 1.20 Aa |
| 20–40 | 22.32 ± 2.40 Ba | 22.1 ± 6.12 Aa | 23.84 ± 10.45 Aa | 22.07 ± 4.15 Aa | 22.78 ± 8.12 Aa | 22.72 ± 1.65 Ba | |
| 40–60 | 18.12 ± 0.24 Ca | 18.5 ± 4.94 Aa | 19.04 ± 4.45 Aa | 18.48 ± 5.35 Aa | 19.55 ± 6.60 Aa | 18.80 ± 1.33 Ca | |
| 2018 | 0–20 | 38.09 ± 2.31 Ba | 34.69 ± 1.02 Cb | 39.44 ± 0.74 Cab | 33.22 ± 0.51 Cb | 42.05 ± 2.56 Bab | 28.90 ± 0.71 Cc |
| 20–40 | 34.36 ± 1.19 Aab | 29.34 ± 0.74 Abc | 34.27 ± 2.55 Aa | 28.53 ± 1.04 Ac | 40.64 ± 2.82 Aa | 23.65 ± 0.70 Ad | |
| 40–60 | 29.36 ± 1.07 Ab | 25.52 ± 0.32 Bc | 27.56 ± 2.27 Bb | 24.16 ± 1.49 Bc | 27.77 ± 0.82 Aa | 19.65 ± 1.35 Bd | |
| Year | Depth (cm) | I | II | III | IV | V | VI |
|---|---|---|---|---|---|---|---|
| pH | |||||||
| 2016 | 0–20 | 4.71 ± 0.16 Ba | 4.74 ± 0.26 Aa | 4.74 ± 0.53 Ba | 4.74 ± 0.15 Aa | 4.69 ± 0.53 Aa | 4.72 ± 0.2 Aa |
| 20–40 | 4.79 ± 0.02 ABa | 4.82 ± 0.19 Aa | 4.83 ± 0.52 ABa | 4.85 ± 0.40 Aa | 4.78 ± 0.69 Aa | 4.81 ± 0.18 Aa | |
| 40–60 | 4.92 ± 0.78 Aa | 4.91 ± 0.22 Aa | 4.96 ± 0.94 Aa | 4.92 ± 1.14 Aa | 4.94 ± 0.43 Aa | 4.93 ± 0.35 Aa | |
| 2018 | 0–20 | 4.50 ± 0.72 Aa | 4.48 ± 0.51 Aa | 5.08 ± 0.34 Aa | 4.61 ± 0.27 Aa | 4.60 ± 0.57 Aa | 4.96 ± 0.32 Aa |
| 20–40 | 5.30 ± 0.49 Aa | 4.79 ± 0.77 Aa | 5.05 ± 0.67 Aa | 4.68 ± 0.33 Aa | 4.75 ± 0.36 Aa | 4.83 ± 0.23 Aa | |
| 40–60 | 5.09 ± 0.72 Aa | 5.47 ± 0.38 Aa | 5.24 ± 0.35 Aa | 5.05 ± 0.16 Aa | 5.02 ± 0.21 Aa | 4.78 ± 0.32 Aa | |
| TN (g/kg) | |||||||
| 2016 | 0–20 | 0.46 ± 0.06 Aa | 0.51 ± 0.16 Aa | 0.50 ± 0.15 Aa | 0.47 ± 0.10 Aa | 0.47 ± 0.28 Aa | 0.48 ± 0.04 Aa |
| 20–40 | 0.17 ± 0.02 Ba | 0.22 ± 0.08 Ba | 0.22 ± 0.09 Ba | 0.23 ± 0.01 Ba | 0.21 ± 0.03 Ba | 0.19 ± 0.04 Ba | |
| 40–60 | 0.15 ± 0.04 Ba | 0.17 ± 0.05 Ba | 0.16 ± 0.06 Ba | 0.18 ± 0.05 Ba | 0.13 ± 0.02 Ba | 0.14 ± 0.03 Ba | |
| 2018 | 0–20 | 0.74 ± 0.03 Aab | 0.71 ± 0.04 Abc | 0.81 ± 0.04 Aa | 0.62 ± 0.02 Ac | 0.68 ± 0.05 Abc | 0.51 ± 0.04 Ad |
| 20–40 | 0.49 ± 0.05 Ba | 0.29 ± 0.04 Bb | 0.56 ± 0.03 Ba | 0.26 ± 0.04 Bb | 0.32 ± 0.03 Bb | 0.22 ± 0.04 Bb | |
| 40–60 | 0.24 ± 0.03 Cab | 0.19 ± 0.04 Cab | 0.28 ± 0.04 Ca | 0.18 ± 0.03 Cb | 0.21 ± 0.03 Cab | 0.17 ± 0.04 Cb | |
| TP (g/kg) | |||||||
| 2016 | 0–20 | 0.24 ± 0.14 Aa | 0.23 ± 0.08 Aa | 0.27 ± 0.11 Aa | 0.23 ± 0.06 Aa | 0.27 ± 0.02 Aa | 0.25 ± 0.03 Aa |
| 20–40 | 0.17 ± 0.1 Aa | 0.18 ± 0.05 ABa | 0.20 ± 0.07 Aa | 0.17 ± 0.05 ABa | 0.18 ± 0.06 Ba | 0.19 ± 0.04 Ba | |
| 40–60 | 0.15 ± 0.06 Aa | 0.14 ± 0.04 Ba | 0.14 ± 0.04 Aa | 0.14 ± 0.08 Ba | 0.15 ± 0.08 Ba | 0.16 ± 0.04 Ba | |
| 2018 | 0–20 | 0.51 ± 0.34 Aab | 0.29 ± 0.06 Aab | 0.37 ± 0.05 Aab | 0.35 ± 0.04 Aab | 0.51 ± 0.23 Aa | 0.27 ± 0.02 Ab |
| 20–40 | 0.29 ± 0.06 ABab | 0.24 ± 0.04 Aab | 0.31 ± 0.03 Aab | 0.27 ± 0.02 Bab | 0.34 ± 0.04 Ba | 0.21 ± 0.05 Ab | |
| 40–60 | 0.26 ± 0.03 Ba | 0.2 ± 0.08 Aa | 0.25 ± 0.03 Ba | 0.26 ± 0.03 Ba | 0.28 ± 0.02 Ca | 0.19 ± 0.06 Ba | |
| Available P (mg/kg) | |||||||
| 2016 | 0–20 | 2.29 ± 1.00 Aa | 2.31 ± 0.63 Aa | 2.32 ± 0.47 Aa | 2.31 ± 0.47 Aa | 2.24 ± 0.58 Aa | 2.27 ± 0.14 Aa |
| 20–40 | 2.10 ± 0.37 Aa | 2.12 ± 0.24 ABa | 2.14 ± 0.75 Aa | 2.13 ± 0.22 Aa | 2.08 ± 0.54 Aa | 2.03 ± 0.06 Ba | |
| 40–60 | 1.91 ± 0.82 Aa | 1.93 ± 0.39 Ba | 1.94 ± 0.96 Aa | 1.95 ± 0.48 Aa | 1.84 ± 0.64 Aa | 1.87 ± 0.16 Ca | |
| 2018 | 0–20 | 2.97 ± 0.05 Aa | 2.68 ± 0.28 Aab | 2.78 ± 0.24 Aab | 2.73 ± 0.15 Aab | 2.94 ± 0.16 Aa | 2.36 ± 0.11 Ab |
| 20–40 | 2.53 ± 0.12 Ba | 2.27 ± 0.10 Bab | 2.45 ± 0.14 Bab | 2.25 ± 0.09 Bab | 2.32 ± 0.17 Bab | 2.14 ± 0.12 ABb | |
| 40–60 | 2.25 ± 0.21 Ca | 2.11 ± 0.07 Ba | 2.19 ± 0.14 Ba | 2.16 ± 0.03 Ba | 2.13 ± 0.30 Ba | 2.02 ± 0.21 Ba | |
| Year | Depth (cm) | I | II | III | IV | V | VI |
|---|---|---|---|---|---|---|---|
| Ure (μg/g·24 h) | |||||||
| 2016 | 0–20 | 268.61 ± 6.89 Ac | 345.85 ± 7.75 Aa | 229.45 ± 2.29 Ae | 226.32 ± 6.09 Ae | 248.72 ± 1.30 Ad | 294.97 ± 6.79 Ab |
| 20–40 | 154.73 ± 4.61 Bcd | 196.69 ± 6.26 Ba | 178.14 ± 5.39 Bb | 199.66 ± 9.20 Ba | 144.16 ± 8.78 Bd | 167.48 ± 3.25 Bbc | |
| 40–60 | 127.03 ± 2.14 Ca | 95.00 ± 7.00 Cb | 121.93 ± 4.96 Ca | 116.7 ± 4.89 Ca | 115.06 ± 6.51 Ca | 72.12 ± 9.43 Cc | |
| 2018 | 0–20 | 563.4 ± 12.56 Aa | 462.99 ± 5.06 Aab | 481.52 ± 1.27 Aa | 378.52 ± 11.33 Aab | 496.9 ± 5.39 Aab | 378.11 ± 7.04 Ab |
| 20–40 | 447.34 ± 12.74 Ba | 346.08 ± 8.88 Bab | 329.24 ± 7.32 Bab | 263.08 ± 11.81 Bbc | 341.41 ± 5.65 Bb | 214.98 ± 4.18 Bc | |
| 31.93 | 125.54 ± 9.72 Ca | 122.84 ± 7.08 Bbc | 181.94 ± 5.26 Ca | 153.38 ± 4.92 Cbc | 154.58 ± 7.34 Cab | 148.88 ± 3.95 Bbc | |
| Suc (mg/g·24 h) | |||||||
| 2016 | 0–20 | 20.53 ± 1.62 Aa | 20.47 ± 2.78 Aa | 21.11 ± 1.57 Aa | 19.63 ± 2.35 Aa | 20.89 ± 3.57 Aa | 20.68 ± 2.36 Aa |
| 20–40 | 13.15 ± 1.45 Ba | 14.17 ± 4.74 Aa | 12.94 ± 3.60 Ba | 14.04 ± 1.48 Ba | 13.91 ± 2.61 Ba | 13.27 ± 2.17 Ba | |
| 40–60 | 6.41 ± 1.40 Ba | 6.75 ± 0.57 Ba | 6.47 ± 1.83 Ca | 6.27 ± 0.61 Ca | 6.16 ± 0.87 Ca | 6.38 ± 0.58 Ca | |
| 2018 | 0–20 | 28.58 ± 2.12 Bab | 24.79 ± 2.31 Abc | 33.23 ± 3.81 Aa | 27.91 ± 3.27 Aab | 31.93 ± 3.46 Aa | 22.59 ± 1.37 Ac |
| 20–40 | 19.76 ± 3.58 Ba | 18.56 ± 11.67 ABa | 23.48 ± 2.78 Ba | 18.73 ± 1.99 Ba | 21.13 ± 3.36 Ba | 15.56 ± 1.99 Ba | |
| 40–60 | 9.16 ± 0.67 Cbc | 10.46 ± 1.04 Bbc | 13.56 ± 2.54 Ca | 8.97 ± 1.93 Cbc | 11.53 ± 1.92 Cab | 7.63 ± 0.58 Cc | |
| Cat (mg/g·24 h) | |||||||
| 2016 | 0–20 | 7.09 ± 1.06 Aa | 7.37 ± 0.45 Aa | 7.1 ± 0.51 Aa | 7.28 ± 0.77 Aa | 7.01 ± 0.58 Aa | 7.05 ± 0.14 Aa |
| 20–40 | 6.51 ± 0.72 Aa | 6.87 ± 0.70 Ba | 6.46 ± 0.28 Aa | 6.81 ± 0.17 Aa | 6.45 ± 1.05 Aa | 6.49 ± 0.21 Ba | |
| 40–60 | 6.07 ± 0.78 Aa | 6.17 ± 0.48 Ba | 6.02 ± 1.04 Aa | 6.46 ± 0.47 Aa | 6.19 ± 1.55 Aa | 6.06 ± 0.15 Ca | |
| 2018 | 0–20 | 8.12 ± 0.21 Aa | 7.48 ± 0.61 Aab | 7.52 ± 0.26 Aab | 7.36 ± 0.2 Aab | 7.74 ± 0.45 Aab | 7.11 ± 0.77 Ab |
| 20–40 | 6.84 ± 0.64 Ba | 6.96 ± 0.77 Aa | 6.93 ± 0.52 ABa | 6.91 ± 0.69 Aa | 6.85 ± 0.51 Ba | 6.63 ± 0.18 Aa | |
| 40–60 | 6.65 ± 0.72 Ba | 6.23 ± 0.50 Aa | 6.56 ± 0.32 Ba | 6.54 ± 0.49 Aa | 6.48 ± 0.23 Ba | 6.18 ± 0.48 Ab | |
| Forest Type | FAC1 | F1 | FCA2 | F2 | Foverall score | Ranking |
|---|---|---|---|---|---|---|
| 2018-I | 1.532 | 4.353 | −1.369 | −1.485 | 3.611 | 2 |
| 2018-II | 0.411 | 1.168 | −1.130 | −1.226 | 0.863 | 5 |
| 2018-III | 1.237 | 3.515 | 2.500 | 2.712 | 3.413 | 3 |
| 2018-IV | 0.384 | 1.091 | −0.359 | −0.389 | 0.903 | 4 |
| 2018-V | 1.667 | 4.736 | −0.183 | −0.198 | 4.108 | 1 |
| 2018-VI | −0.513 | −1.456 | 1.149 | 1.247 | −1.113 | 6 |
| 2016-I | −0.888 | 2.523 | −0.187 | −0.203 | −2.228 | 12 |
| 2016-II | −0.600 | −1.704 | −0.129 | −0.140 | −1.505 | 7 |
| 2016-III | −0.725 | −2.061 | 0.097 | 0.105 | −1.785 | 8 |
| 2016-IV | −0.809 | −2.298 | −0.104 | −0.113 | −2.020 | 9 |
| 2016-V | −0.864 | −2.455 | −0.192 | −0.208 | −2.169 | 11 |
| 2016-VI | −0.832 | −2.366 | −0.092 | −0.100 | −2.077 | 10 |
| pH | TN | TP | AP | BD | TOP | MC | Ure | Suc | Cat | |
|---|---|---|---|---|---|---|---|---|---|---|
| pH | 1 | |||||||||
| TN | −0.176 | 1 | ||||||||
| TP | −0.080 | 0.658 ** | 1 | |||||||
| AP | −0.162 | 0.845 ** | 0.690 ** | 1 | ||||||
| BD | 0.174 | −0.659 ** | −0.699 ** | −0.597 ** | 1 | |||||
| TOP | −0.086 | 0.624 ** | 0.723 ** | −0.670 ** | −0.704 ** | 1 | ||||
| MC | −0.094 | 0.773 ** | 0.755 ** | 0.727 ** | −0.763 ** | 0.881 ** | 1 | |||
| Ure | −0.166 | 0.872 ** | 0.618 ** | 0.828 ** | −0.579 ** | 0.625 ** | 0.768 ** | 1 | ||
| Suc | −0.163 | 0.859 ** | 0.690 ** | 0.745 ** | −0.681 ** | 0.642 ** | 0.775 ** | 0.814 ** | 1 | |
| Cat | −0.011 | 0.667 ** | 0.467 ** | 0.671 ** | −0.471 ** | 0.496 ** | 0.576 ** | 0.711 ** | 0.617 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Liu, L.; Zhang, Z.; Ze, S.; Ji, M.; Li, Z.; Yu, J.; Yang, B.; Zhao, N. Do Mixed Pinus yunnanensis Plantations Improve Soil’s Physicochemical Properties and Enzyme Activities? Diversity 2022, 14, 214. https://doi.org/10.3390/d14030214
Liang C, Liu L, Zhang Z, Ze S, Ji M, Li Z, Yu J, Yang B, Zhao N. Do Mixed Pinus yunnanensis Plantations Improve Soil’s Physicochemical Properties and Enzyme Activities? Diversity. 2022; 14(3):214. https://doi.org/10.3390/d14030214
Chicago/Turabian StyleLiang, Chen, Ling Liu, Zhixiao Zhang, Sangzi Ze, Mei Ji, Zongbo Li, Jinde Yu, Bin Yang, and Ning Zhao. 2022. "Do Mixed Pinus yunnanensis Plantations Improve Soil’s Physicochemical Properties and Enzyme Activities?" Diversity 14, no. 3: 214. https://doi.org/10.3390/d14030214