

High-Resolution Mapping of Seagrass Biomass Dynamics Suggests Differential Response of Seagrasses to Fluctuating Environments

Abstract
:1. Introduction
2. Materials and Methods
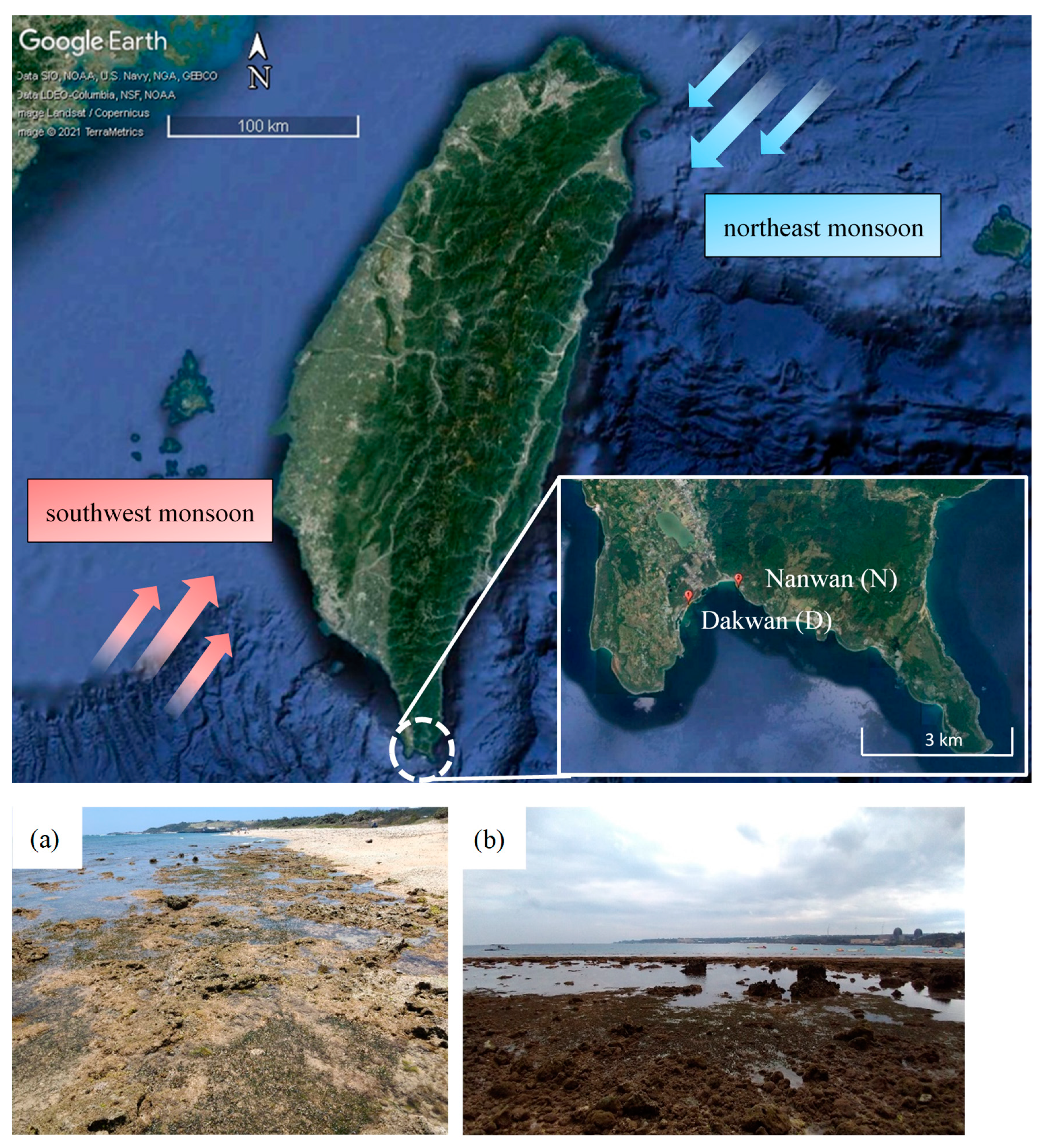
2.1. Study Sites
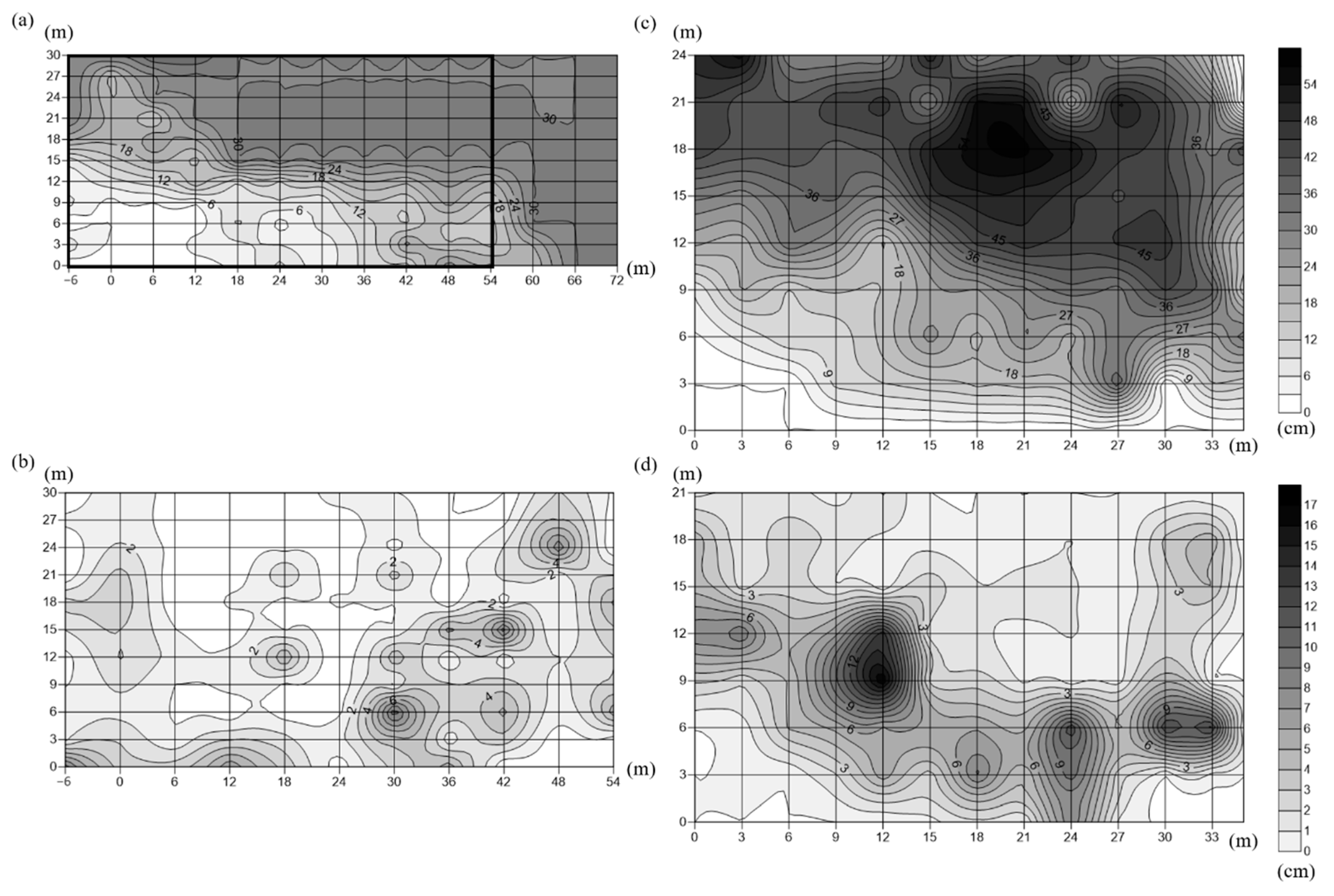
2.2. Sediment Features and Water Quality
2.3. Seagrass Shoot Density and Biomass
2.4. Seagrass Productivity
2.5. High-Resolution Mapping of Seagrass Biomass Dynamics
2.6. Statistical Analyses
3. Results
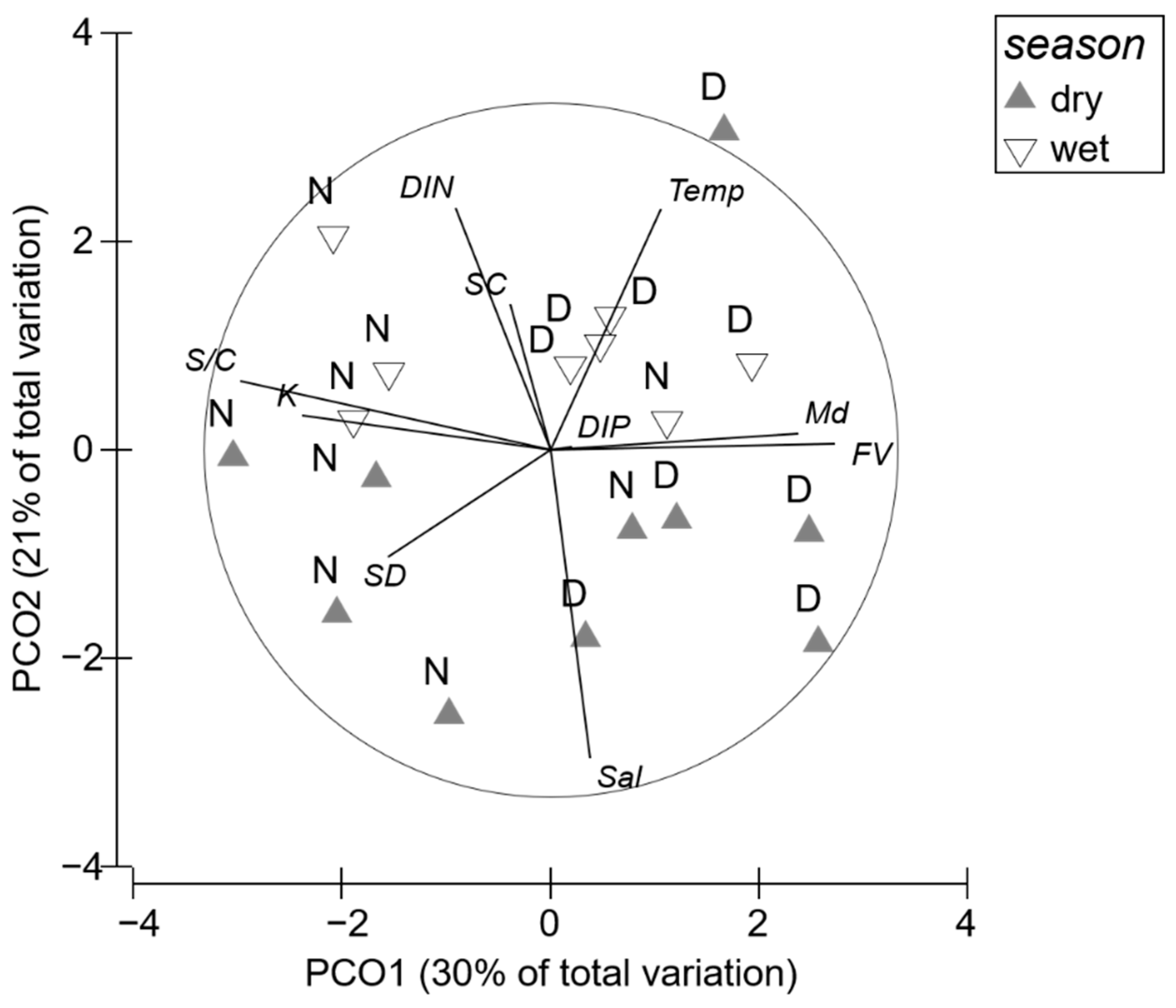
3.1. Environmental Factors
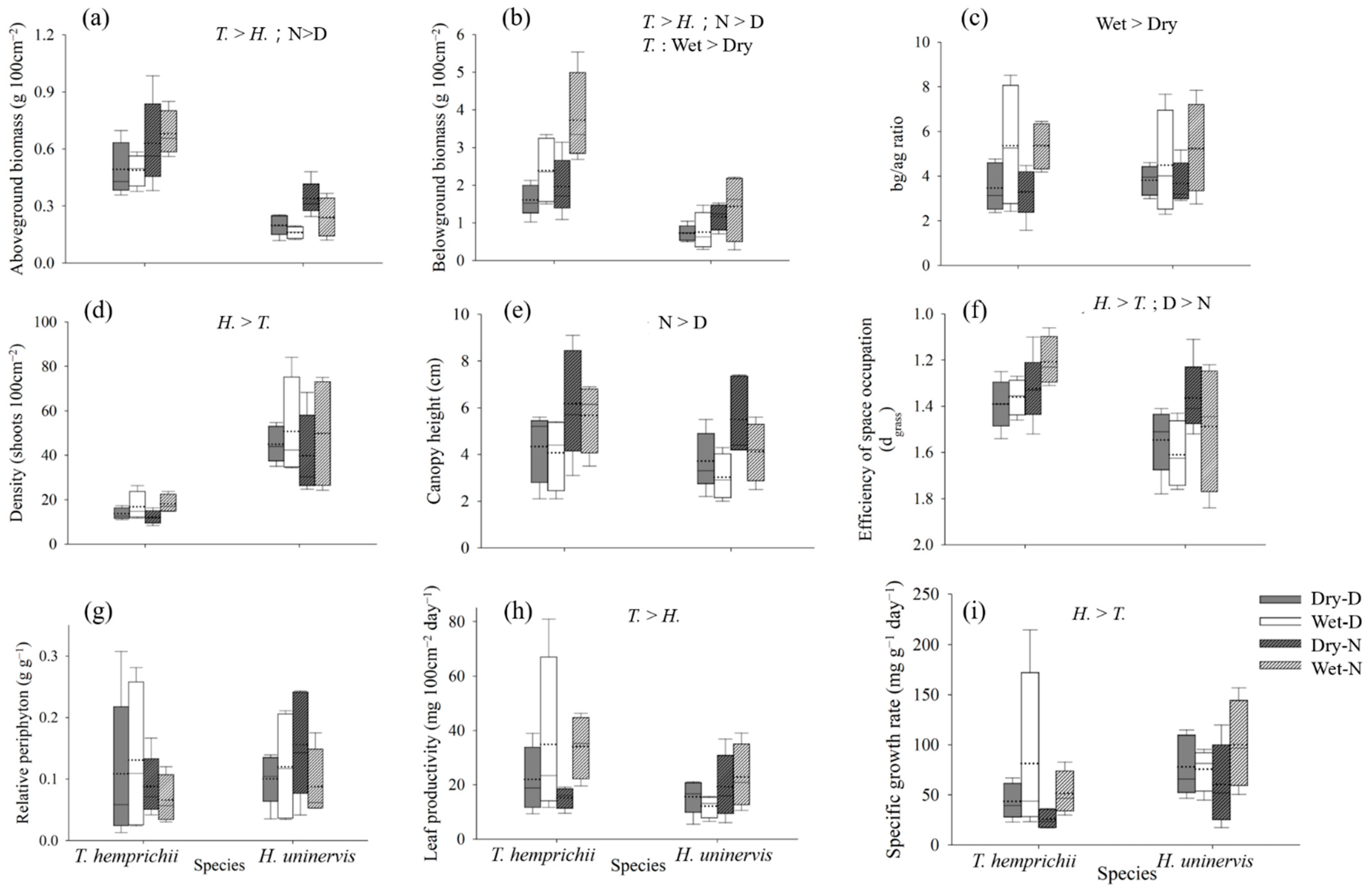
3.2. Seagrass Variables
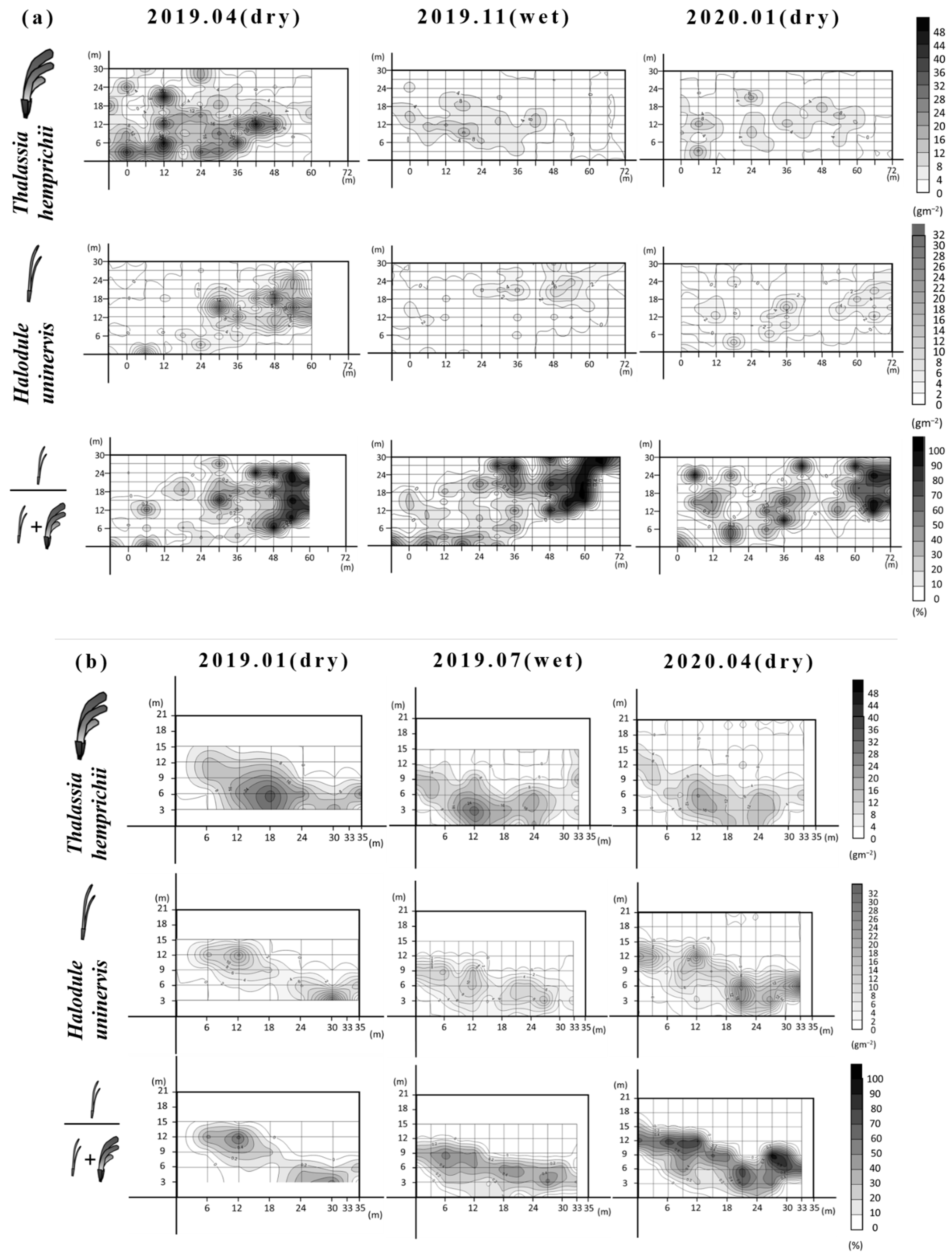
3.3. High-Resolution Mapping of Seagrass Biomass Dynamics
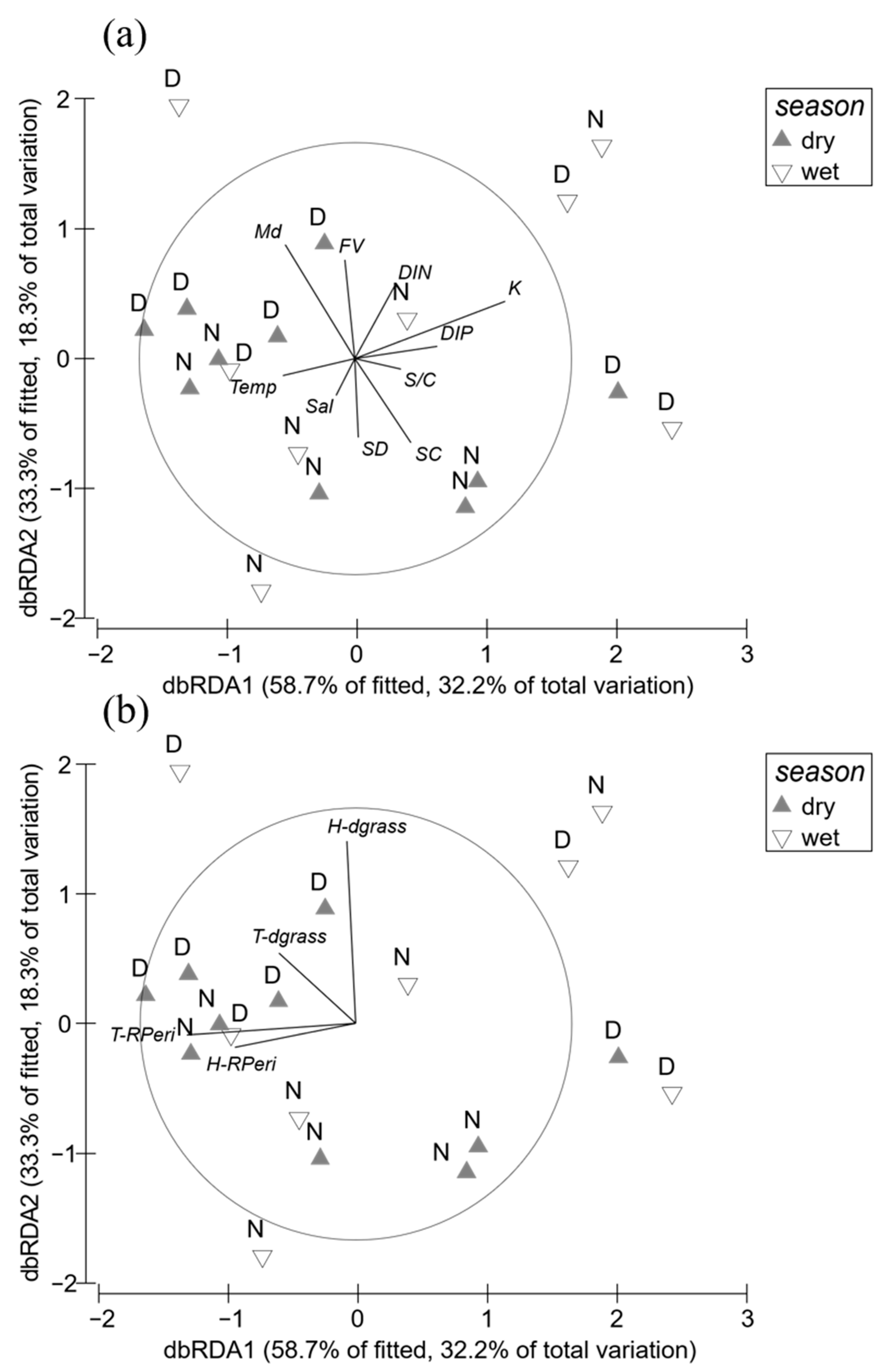
3.4. Relationship between Seagrass Variables and Environmental Factors
4. Discussion
4.1. Competition in Seagrass Community
4.2. Differential Response of Seagrasses to Environments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelleman, C.; Corcoran, E.; Duarte, C.M.; Valdes, L.; DeYoung, C.; Fonseca, L.; Grimsditch, G. Blue Carbon: The Role of Healthy Oceans in Binding Carbon; UNEP/FAO/UNESCO/IUCN/CSIC: Madrid, Spain, 2008. [Google Scholar]
- Laffoley, D.; Grimsditch, G.D. The Management of Natural Coastal Carbon Sinks; IUCN: Gland, Vaud, Switzerland, 2009. [Google Scholar]
- Davidson, N.C.; Finlayson, C.M. Updating global coastal wetland areas presented in Davidson and Finlayson (2018). Mar. Freshw. Res. 2019, 70, 1195–1200. [Google Scholar] [CrossRef]
- Lin, W.-J.; Wu, J.; Lin, H.-J. Contribution of unvegetated tidal flats to coastal carbon flux. Glob. Chang. Biol. 2020, 26, 3443–3454. [Google Scholar] [CrossRef]
- Mukai, H. Biogeography of the tropical seagrasses in the western Pacific. Mar. Freshw. Res. 1993, 44, 1–17. [Google Scholar] [CrossRef]
- Lin, H.-J.; Lee, C.-L.; Peng, S.-E.; Hung, M.-C.; Liu, P.-J.; Mayfield, A.B. The effects of El Niño-Southern Oscillation events on intertidal seagrass beds over a long-term timescale. Glob. Chang. Biol. 2018, 24, 4566–4580. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-H.; Lee, C.-L.; Chung, C.-Y.; Hsiao, S.-C.; Lin, H.-J. Carbon budgets of multispecies seagrass beds at Dongsha Island in the South China Sea. Mar. Environ. Res. 2015, 106, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.-F.; Chen, K.-Y.; Lin, H.-J. Significance of belowground production to the long-term carbon sequestration of intertidal seagrass beds. Sci. Total Environ. 2021, 800, 149579. [Google Scholar] [CrossRef] [PubMed]
- Friedlingstein, P.; Jones, M.W.; O’Sullivan, M.; Andrew, R.M.; Bakker, D.C.; Hauck, J.; Le Quéré, C.; Peters, G.P.; Peters, W.; Pongratz, J. Global carbon budget 2021. Earth Syst. Sci. Data 2022, 14, 1917–2005. [Google Scholar] [CrossRef]
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, R.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.; Plattner, G.-K.; Rogelj, J.; et al. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Technical Summary. In Proceedings of the Intergovernmental Panel on Climate Change AR6; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar]
- Mazarrasa, I.; Samper-Villarreal, J.; Serrano, O.; Lavery, P.S.; Lovelock, C.E.; Marbà, N.; Duarte, C.M.; Cortés, J. Habitat characteristics provide insights of carbon storage in seagrass meadows. Mar. Pollut. Bull. 2018, 134, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Krause-Jensen, D.; Duarte, C.M. Expansion of vegetated coastal ecosystems in the future Arctic. Front. Mar. Sci. 2014, 1, 77. [Google Scholar] [CrossRef] [Green Version]
- Clausen, K.K.; Krause-Jensen, D.; Olesen, B.; Marbà, N. Seasonality of eelgrass biomass across gradients in temperature and latitude. Mar. Ecol. Prog. Ser. 2014, 506, 71–85. [Google Scholar] [CrossRef]
- Collier, C.J.; Langlois, L.; Ow, Y.; Johansson, C.; Giammusso, M.; Adams, M.P.; O’Brien, K.R.; Uthicke, S. Losing a winner: Thermal stress and local pressures outweigh the positive effects of ocean acidification for tropical seagrasses. New Phytol. 2018, 219, 1005–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, R.; Gullström, M.; Mangora, M.M.; Mtolera, M.S.; Björk, M. High midday temperature stress has stronger effects on biomass than on photosynthesis: A mesocosm experiment on four tropical seagrass species. Ecol. Evol. 2018, 8, 4508–4517. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.I.; Leon, J.; Phinn, S.R.; Callaghan, D.P.; O’Brien, K.R.; Roelfsema, C.M.; Lovelock, C.E.; Lyons, M.B.; Mumby, P.J. Coastal retreat and improved water quality mitigate losses of seagrass from sea level rise. Glob. Chang. Biol. 2013, 19, 2569–2583. [Google Scholar] [CrossRef]
- Short, F.T.; Neckles, H.A. The effects of global climate change on seagrasses. Aquat. Bot. 1999, 63, 169–196. [Google Scholar] [CrossRef]
- Congdon, V.M.; Bonsell, C.; Cuddy, M.R.; Dunton, K.H. In the wake of a major hurricane: Differential effects on early vs. late successional seagrass species. Limnol. Oceanogr. Lett. 2019, 4, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.-J.; Chang, H.-F.; Mayfield, A.B.; Lin, H.-J. Assessing the Effects of Ocean Warming and Acidification on the Seagrass Thalassia hemprichii. J. Mar. Sci. Eng. 2022, 10, 714. [Google Scholar] [CrossRef]
- Lin, H.-J.; Shao, K.-T. Temporal changes in the abundance and growth of intertidal Thalassia hemprichii seagrass beds in southern Taiwan. Bot. Bull. Acad. Sin. 1998, 39, 191–198. [Google Scholar]
- Lee, C.-L.; Lin, W.-J.; Liu, P.-J.; Shao, K.-T.; Lin, H.-J. Highly productive tropical seagrass beds support diverse consumers and a large organic carbon pool in the sediments. Diversity 2021, 13, 544. [Google Scholar] [CrossRef]
- Hsieh, H.-L. Spatial and temporal patterns of polychaete communities in a subtropical mangrove swamp: Influences of sediment and microhabitat. Mar. Ecol. Prog. Ser. 1995, 127, 157–167. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Pai, S.-C.; Tsau, Y.-J.; Yang, T.-I. pH and buffering capacity problems involved in the determination of ammonia in saline water using the indophenol blue spectrophotometric method. Anal. Chim. Acta 2001, 434, 209–216. [Google Scholar] [CrossRef]
- Jenkins, D.; Medsker, L.L. Brucine Method for the Determination of Nitrate in Ocean, Estuarine, and Fresh Waters. Anal. Chem. 1964, 36, 610–612. [Google Scholar] [CrossRef]
- Pai, S.-C.; Yang, C.-C.; Riley, J.P. Formation kinetics of the pink azo dye in the determination of nitrite in natural waters. Anal. Chim. Acta 1990, 232, 345–349. [Google Scholar] [CrossRef]
- Doty, M.S. Measurement of water movement in reference to benthic algal growth. Bot. Mar. 1971, 14, 32–35. [Google Scholar] [CrossRef]
- Lin, H.-J.; Wang, T.-C.; Su, H.-M.; Hung, J.-J. Relative importance of phytoplankton and periphyton on oyster-culture pens in a eutrophic tropical lagoon. Aquaculture 2005, 243, 279–290. [Google Scholar] [CrossRef]
- Short, F.T.; Coles, R.G. Global Seagrass Research Methods; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Vieira, V.M.; Lopes, I.E.; Creed, J.C. The biomass–density relationship in seagrasses and its use as an ecological indicator. BMC Ecol. 2018, 18, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R.; Warwick, R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Tilman, D. The resource-ratio hypothesis of plant succession. Am. Nat. 1985, 125, 827–852. [Google Scholar] [CrossRef]
- Rose, C.D.; Dawes, C.J. Effects of community structure on the seagrass Thalassia testudinum. Mar. Ecol. Prog. Ser. 1999, 184, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, M.E.; Merino, M.; Rodriguez, A.; Marbà, N.; Duarte, C.M. Growth patterns and demography of pioneer Caribbean seagrasses Halodule wrightii and Syringodium filiforme. Mar. Ecol. Prog. Ser. 1994, 109, 99–104. [Google Scholar] [CrossRef]
- Zieman, J.C. The Ecology of the Seagrasses of South Florida: A Community Profile; Department of the Interior, US Fish and Wildlife Service: Washington, DC, USA, 1982.
- Iverson, R.L.; Bittaker, H.F. Seagrass distribution and abundance in Eastern Gulf of Mexico coastal waters. Estuar. Coast. Shelf Sci. 1986, 22, 577–602. [Google Scholar] [CrossRef]
- Lan, C.-Y.; Kao, W.-Y.; Lin, H.-J.; Shao, K.-T. Measurement of chlorophyll fluorescence reveals mechanisms for habitat niche separation of the intertidal seagrasses Thalassia hemprichii and Halodule uninervis. Mar. Biol. 2005, 148, 25–34. [Google Scholar] [CrossRef]
- Fourqurean, J.W.; Powell, G.V.; Kenworthy, W.J.; Zieman, J.C. The effects of long-term manipulation of nutrient supply on competition between the seagrasses Thalassia testudinum and Halodule wrightii in Florida Bay. Oikos 1995, 72, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Furman, B.T.; Merello, M.; Shea, C.P.; Kenworthy, W.J.; Hall, M.O. Monitoring of physically restored seagrass meadows reveals a slow rate of recovery for Thalassia testudinum. Restor. Ecol. 2019, 27, 421–430. [Google Scholar] [CrossRef]
- Gaubert-Boussarie, J.; Altieri, A.H.; Duffy, J.E.; Campbell, J.E. Seagrass structural and elemental indicators reveal high nutrient availability within a tropical lagoon in Panama. PeerJ 2021, 9, e11308. [Google Scholar] [CrossRef]
- Robbins, B.D.; Bell, S.S. Dynamics of a subtidal seagrass landscape: Seasonal and annual change in relation to water depth. Ecology 2000, 81, 1193–1205. [Google Scholar] [CrossRef]
- Jupp, B.; Durako, M.J.; Kenworthy, W.; Thayer, G.; Schillak, L. Distribution, abundance, and species composition of seagrasses at several sites in Oman. Aquat. Bot. 1996, 53, 199–213. [Google Scholar] [CrossRef]
- La Nafie, Y.A.; De Los Santos, C.B.; Brun, F.G.; Van Katwijk, M.M.; Bouma, T.J. Waves and high nutrient loads jointly decrease survival and separately affect morphological and biomechanical properties in the seagrass Zostera noltii. Limnol. Oceanogr. 2012, 57, 1664–1672. [Google Scholar] [CrossRef] [Green Version]
- Chiu, S.-H.; Huang, Y.-H.; Lin, H.-J. Carbon budget of leaves of the tropical intertidal seagrass Thalassia hemprichii. Estuar. Coast. Shelf Sci. 2013, 125, 27–35. [Google Scholar] [CrossRef]
- Burkholder, J.M.; Tomasko, D.A.; Touchette, B.W. Seagrasses and eutrophication. J. Exp. Mar. Biol. Ecol. 2007, 350, 46–72. [Google Scholar] [CrossRef]
- Camacho, R.; Houk, P. Decoupling seasonal and temporal dynamics of macroalgal canopy cover in seagrass beds. J. Exp. Mar. Biol. Ecol. 2020, 525, 151310. [Google Scholar] [CrossRef]
- Lin, H.-J.; Wu, C.-Y.; Kao, S.-J.; Kao, W.-Y.; Meng, P.-J. Mapping anthropogenic nitrogen through point sources in coral reefs using δ15N in macroalgae. Mar. Ecol. Prog. Ser. 2007, 335, 95–109. [Google Scholar] [CrossRef]
- Strand, J.A.; Weisner, S.E. Wave exposure related growth of epiphyton: Implications for the distribution of submerged macrophytes in eutrophic lakes. Hydrobiologia 1996, 325, 113–119. [Google Scholar] [CrossRef]
- Christianen, M.J.; Govers, L.L.; Bouma, T.J.; Kiswara, W.; Roelofs, J.G.; Lamers, L.P.; van Katwijk, M.M. Marine megaherbivore grazing may increase seagrass tolerance to high nutrient loads. J. Ecol. 2012, 100, 546–560. [Google Scholar] [CrossRef]
- Lee, C.-L.; Huang, Y.-H.; Chung, C.-Y.; Lin, H.-J. Tidal variation in fish assemblages and trophic structures in tropical Indo-Pacific seagrass beds. Zool. Stud. 2014, 53, 56. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.G. Development of an epiphyte indicator of nutrient enrichment: Threshold values for seagrass epiphyte load. Ecol. Indic. 2017, 74, 343–356. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Dakwan (D) | Nanwan (N) | |||
|---|---|---|---|---|---|
| (21.95038, 120.74888) | (21.95646, 120.76826) | ||||
| Season | Dry | Wet | Dry | Wet | |
| Seawater temperature (°C) | 28.12 ± 3.30 | 32.89 ± 2.39 | 27.96 ± 2.83 | 33.15 ± 1.51 | |
| Salinity | 34.81 ± 0.17 | 33.21 ± 0.23 | 34.66 ± 0.20 | 32.99 ± 0.70 | |
| Light extinction coefficient (m−1) | 0.97 ± 0.30 | 1.54 ± 0.77 | 1.88 ± 0.62 | 1.63 ± 0.51 | |
| Seawater DIN (μM) | 4.02 ± 2.00 | 9.45 ± 5.59 | 3.12 ± 1.45 | 9.89 ± 8.68 | |
| Seawater DIP (μM) | 0.14 ± 0.09 | 0.21 ± 0.09 | 0.13 ± 0.10 | 0.16 ± 0.09 | |
| Flow velocity (%) | 49.96 ± 3.52 | 43.78 ± 3.25 | 36.59 ± 4.54 | 40.42 ± 6.08 | |
| Sediment depth (cm) | 7.79 ± 0.87 | 6.14 ± 2.41 | 12.68 ± 4.43 | 14.61 ± 4.01 | |
| Medium grain size (mm) | 0.59 ± 0.28 | 0.53 ± 0.23 | 0.36 ± 0.14 | 0.36 ± 0.11 | |
| Silt/clay content (%) | 1.69 ± 0.88 | 2.15 ± 0.62 | 5.00 ± 2.17 | 4.67 ± 1.63 | |
| Sorting coefficient | 1.70 ± 0.4 | 1.72 ± 0.39 | 1.75 ± 0.32 | 1.77 ± 0.17 | |
| T. hemprichii | Aboveground biomass (g 100 cm−2) | 0.49 ± 0.12 | 0.49 ± 0.07 | 0.63 ± 0.20 | 0.68 ± 0.11 |
| Belowground biomass (g 100 cm−2) | 1.61 ± 0.37 | 2.39 ± 0.78 | 1.97 ± 0.68 | 3.73 ± 1.08 | |
| bg/ag ratio | 3.47 ± 0.96 | 5.37 ± 2.39 | 3.29 ± 0.97 | 5.35 ± 0.93 | |
| Density (shoots 100 cm−2) | 13.73 ± 2.28 | 16.83 ± 5.66 | 12.13 ± 2.71 | 18.08 ± 3.56 | |
| Canopy height (cm) | 4.35 ± 1.32 | 4.09 ± 1.35 | 6.17 ± 2.10 | 5.68 ± 1.30 | |
| Periphyton biomass (g 100 cm−2) | 0.05 ± 0.06 | 0.06 ± 0.06 | 0.06 ± 0.02 | 0.04 ± 0.02 | |
| Relative periphyton biomass (g g−1) | 0.11 ± 0.11 | 0.13 ± 0.11 | 0.09 ± 0.04 | 0.07 ± 0.03 | |
| Leaf productivity (mg 100 cm−2 day−1) | 21.98 ± 10.64 | 34.86 ± 27.02 | 15.18 ± 3.47 | 34.07 ± 10.15 | |
| Specific growth rate (mg g−1 day−1) | 43.6 ± 15.85 | 81.34 ± 77.46 | 25.86 ± 8.16 | 51.44 ± 19.28 | |
| Efficiency of space occupation (dgrass) | 1.39 ± 0.10 | 1.36 ± 0.07 | 1.32 ± 0.13 | 1.21 ± 0.09 | |
| H. uninervis | Aboveground biomass (g 100 cm−2) | 0.20 ± 0.05 | 0.16 ± 0.03 | 0.34 ± 0.08 | 0.24 ± 0.09 |
| Belowground biomass (g 100 cm−2) | 0.73 ± 0.19 | 0.76 ± 0.44 | 1.16 ± 0.31 | 1.44 ± 0.78 | |
| bg/ag ratio | 3.81 ± 0.60 | 4.49 ± 2.04 | 3.67 ± 0.84 | 5.26 ± 1.80 | |
| Density (shoots 100 cm−2) | 45.00 ± 7.20 | 50.75 ± 19.98 | 39.80 ± 16.33 | 49.83 ± 21.70 | |
| Canopy height (cm) | 3.70 ± 1.12 | 3.02 ± 0.83 | 5.49 ± 1.52 | 4.11 ± 1.10 | |
| Periphyton biomass (g 100 cm−2) | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.06 ± 0.04 | 0.03 ± 0.03 | |
| Relative periphyton biomass (g g−1) | 0.10 ± 0.04 | 0.12 ± 0.08 | 0.16 ± 0.08 | 0.09 ± 0.05 | |
| Leaf productivity (mg 100 cm−2 day−1) | 15.58 ± 5.62 | 12.15 ± 3.50 | 19.29 ± 10.64 | 22.84 ± 10.36 | |
| Specific growth rate (mg g−1 day−1) | 77.98 ± 26.73 | 75.68 ± 18.78 | 60.41 ± 36.22 | 100.01 ± 38.49 | |
| Efficiency of space occupation (dgrass) | 1.55 ± 0.13 | 1.61 ± 0.13 | 1.36 ± 0.14 | 1.49 ± 0.24 | |
| Location | Species | Time (Season) | 2019.04 (Dry) | 2019.11 (Wet) | 2020.01 (Dry) |
| Dakwan | T. hemprichii | Cover area (m2) | 1693 | 1862 | 1756 |
| Average aboveground biomass density (range) (g/m2) | 10.7 (0.1–49.5) | 3.4 (0.1–18.5) | 3.5 (0.1–18.6) | ||
| Total aboveground biomass (g) | 18236 | 6491 | 6683 | ||
| Dominant area (m2) (%) | 1228.0 (83.1%) | 1268.2 (78.0%) | 1305.7 (84.8%) | ||
| H. uninervis | Average aboveground biomass density (range) (g/m2) | 3.9 (0.0–29.1) | 1.6 (0.1–7.9) | 1.8 (0.0–10.8) | |
| Total aboveground biomass (g) | 6646 | 3022 | 3177 | ||
| Dominant area (m2) (%) | 250.0 (16.9%) | 358.1 (22.0%) | 234.0 (15.2%) | ||
| Location | Species | Time (Season) | 2019.01 (Dry) | 2019.07 (Wet) | 2020.04 (Dry) |
| Nanwan | T. hemprichii | Cover area (m2) | 331 | 355 | 414 |
| Average aboveground biomass density (range) (g/m2) | 11.4 (5.7–34.4) | 8.6 (0.7–31.6) | 6.3 (0.2–20.8) | ||
| Total aboveground biomass (g) | 4034 | 3635 | 3067 | ||
| Dominant area (m2) (%) | 313.1 (98.3%) | 305.1 (99.1%) | 259.2 (72.1%) | ||
| H. uninervis | Average aboveground biomass density (range) (g/m2) | 4.3 (0.4–26.2) | 4.5 (0.3–12.6) | 6.5 (0.3–30.6) | |
| Total aboveground biomass (g) | 1425 | 1586 | 2685 | ||
| Dominant area (m2) (%) | 5.3 (1.7%) | 2.8 (0.9%) | 100.2 (27.9%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.-Y.; Lin, H.-J. High-Resolution Mapping of Seagrass Biomass Dynamics Suggests Differential Response of Seagrasses to Fluctuating Environments. Diversity 2022, 14, 999. https://doi.org/10.3390/d14110999
Chen K-Y, Lin H-J. High-Resolution Mapping of Seagrass Biomass Dynamics Suggests Differential Response of Seagrasses to Fluctuating Environments. Diversity. 2022; 14(11):999. https://doi.org/10.3390/d14110999
Chicago/Turabian StyleChen, Kuan-Yu, and Hsing-Juh Lin. 2022. "High-Resolution Mapping of Seagrass Biomass Dynamics Suggests Differential Response of Seagrasses to Fluctuating Environments" Diversity 14, no. 11: 999. https://doi.org/10.3390/d14110999