
Resilience and Species Accumulation across Seafloor Habitat Transitions in a Northern New Zealand Harbour
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
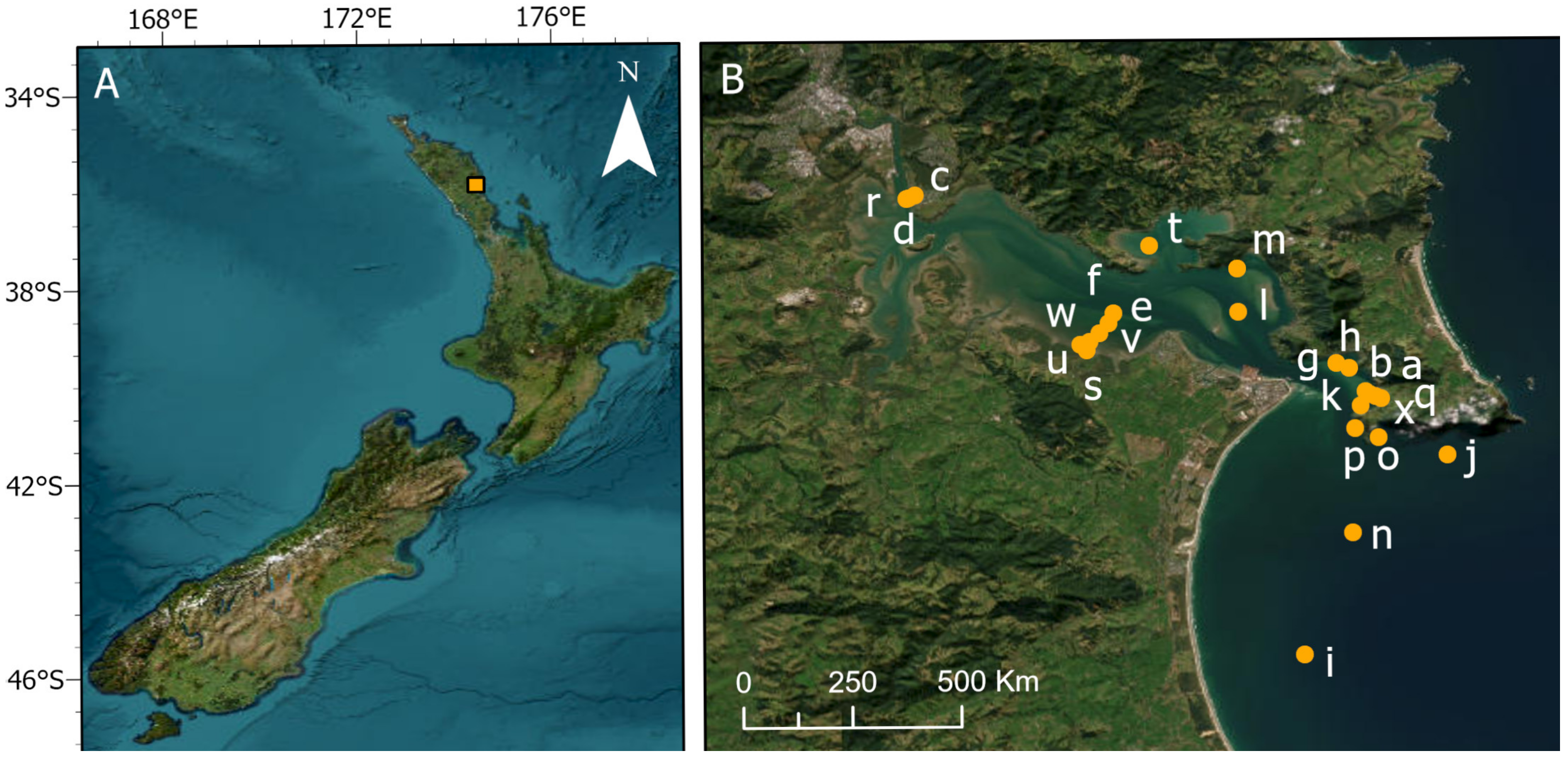
2.1. Study Area and Sampling
2.2. Data Analysis
3. Results
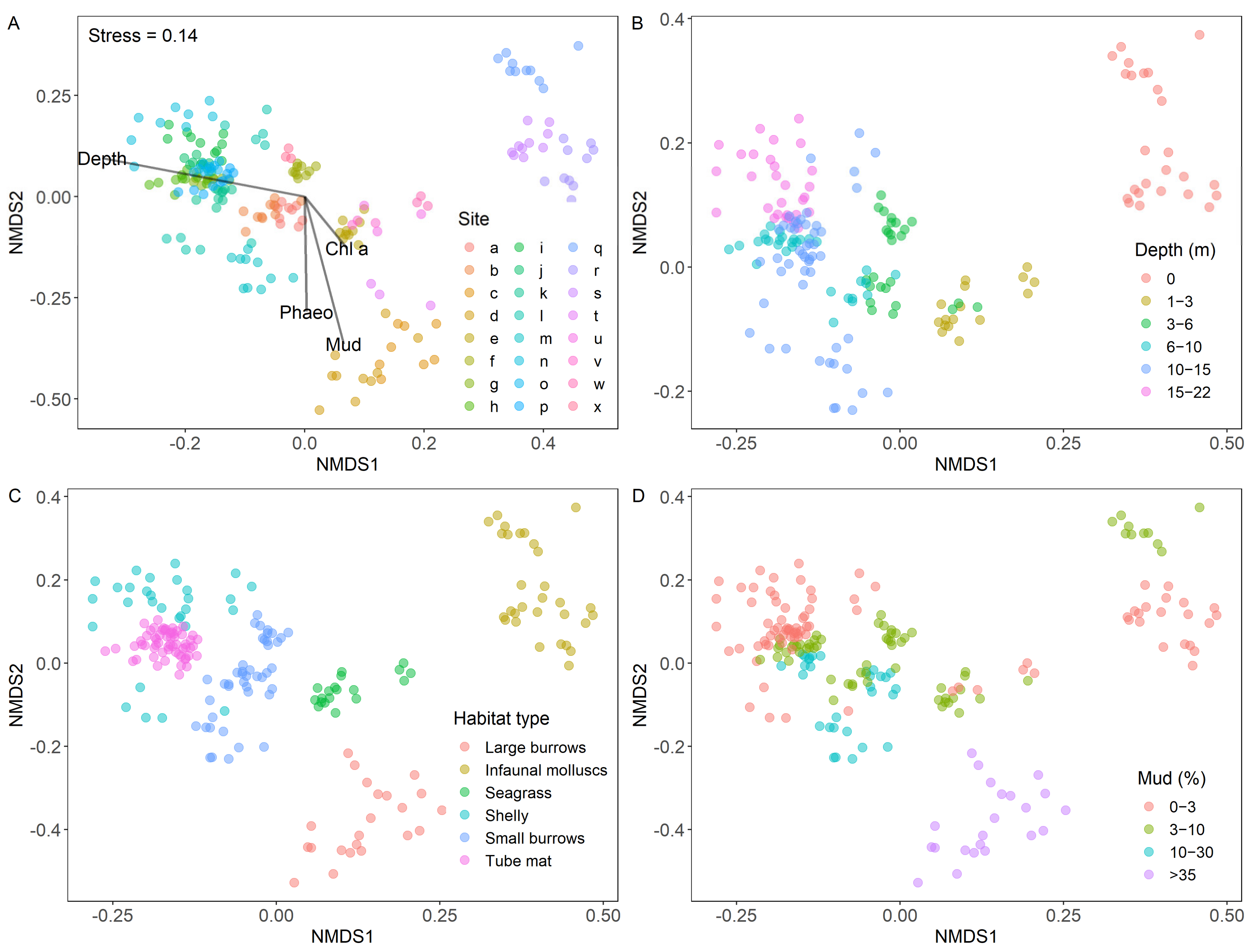
3.1. Estuary Characterisation and Group Selection
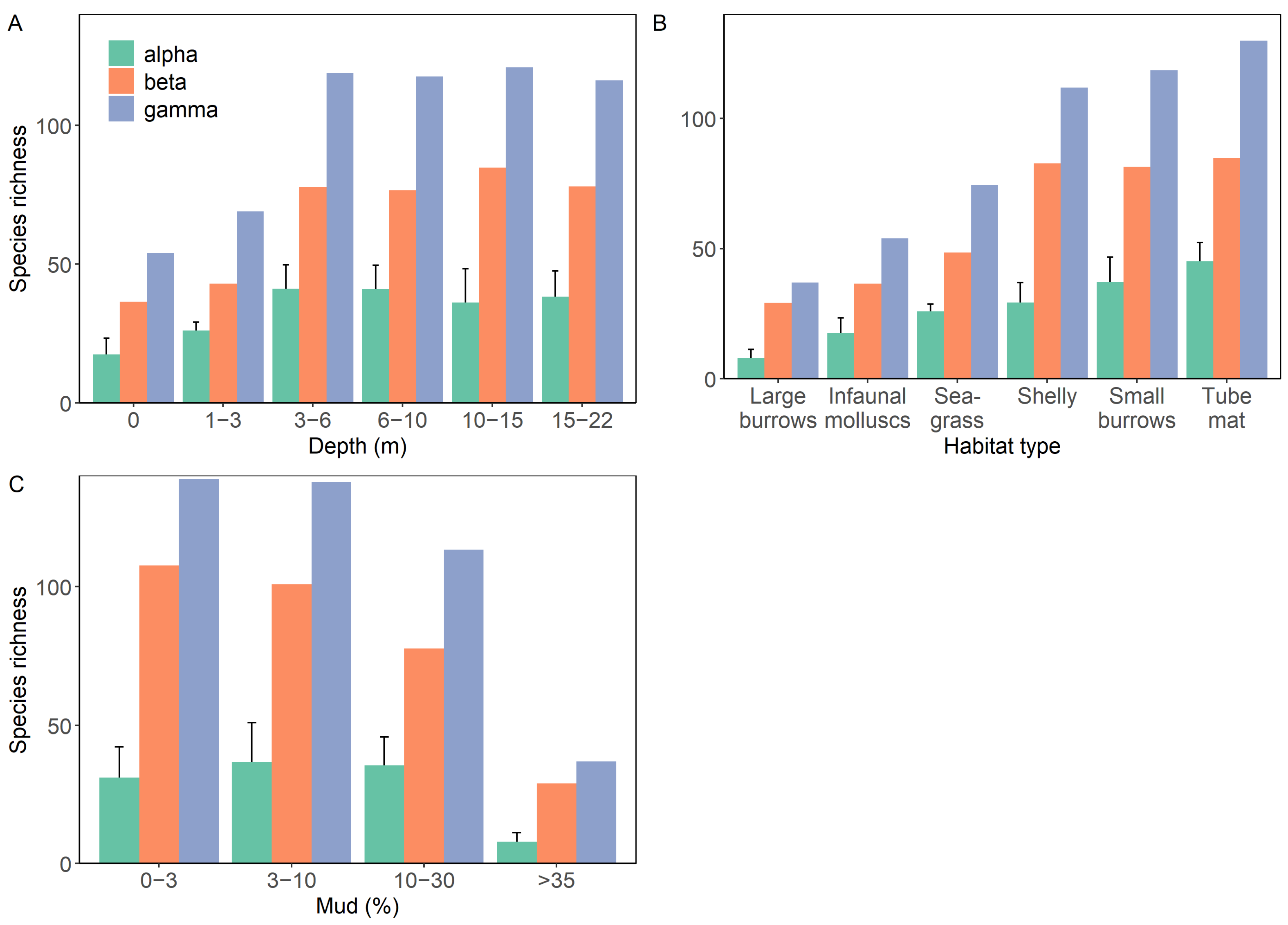
3.2. Patterns of Diversity within Groups
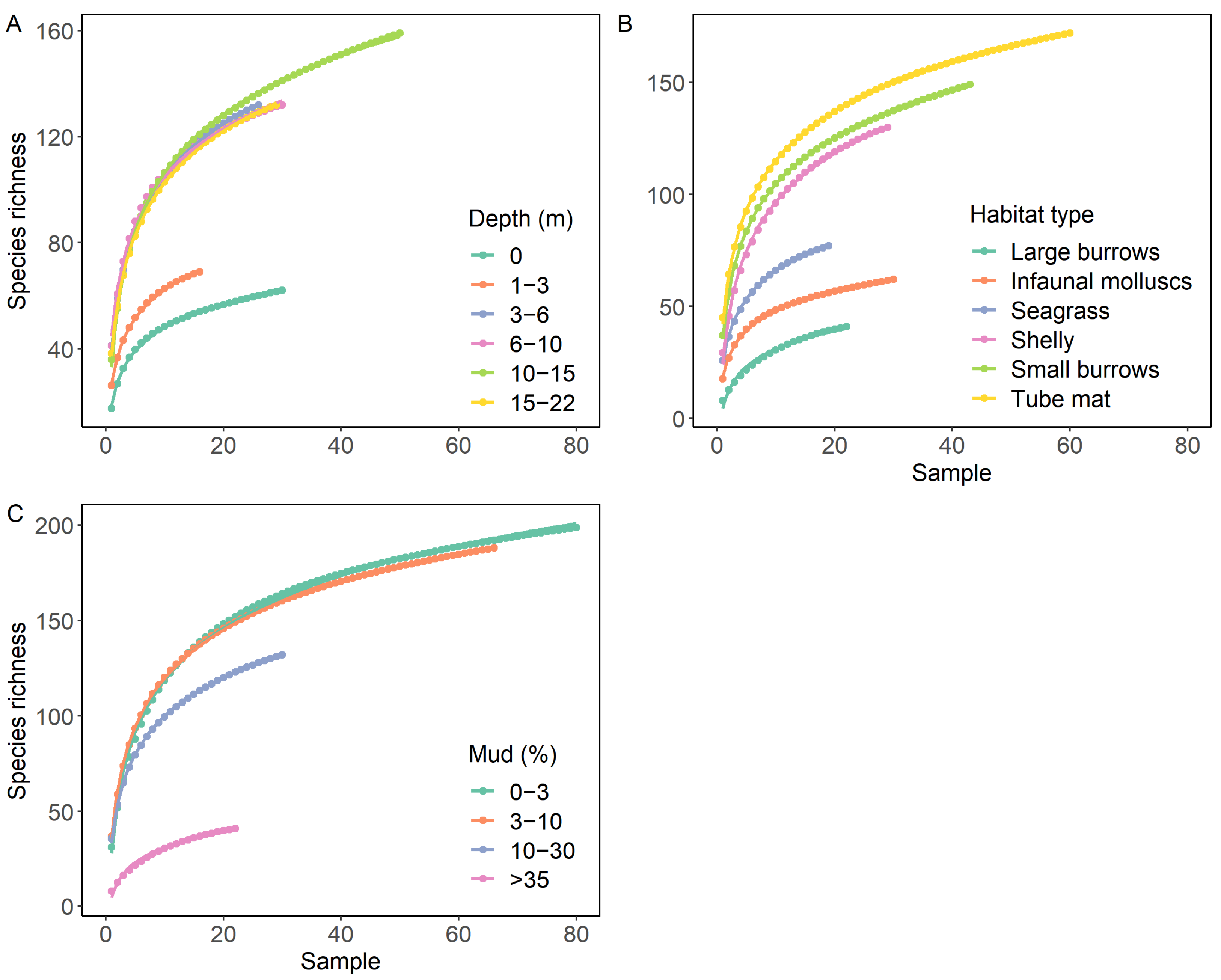
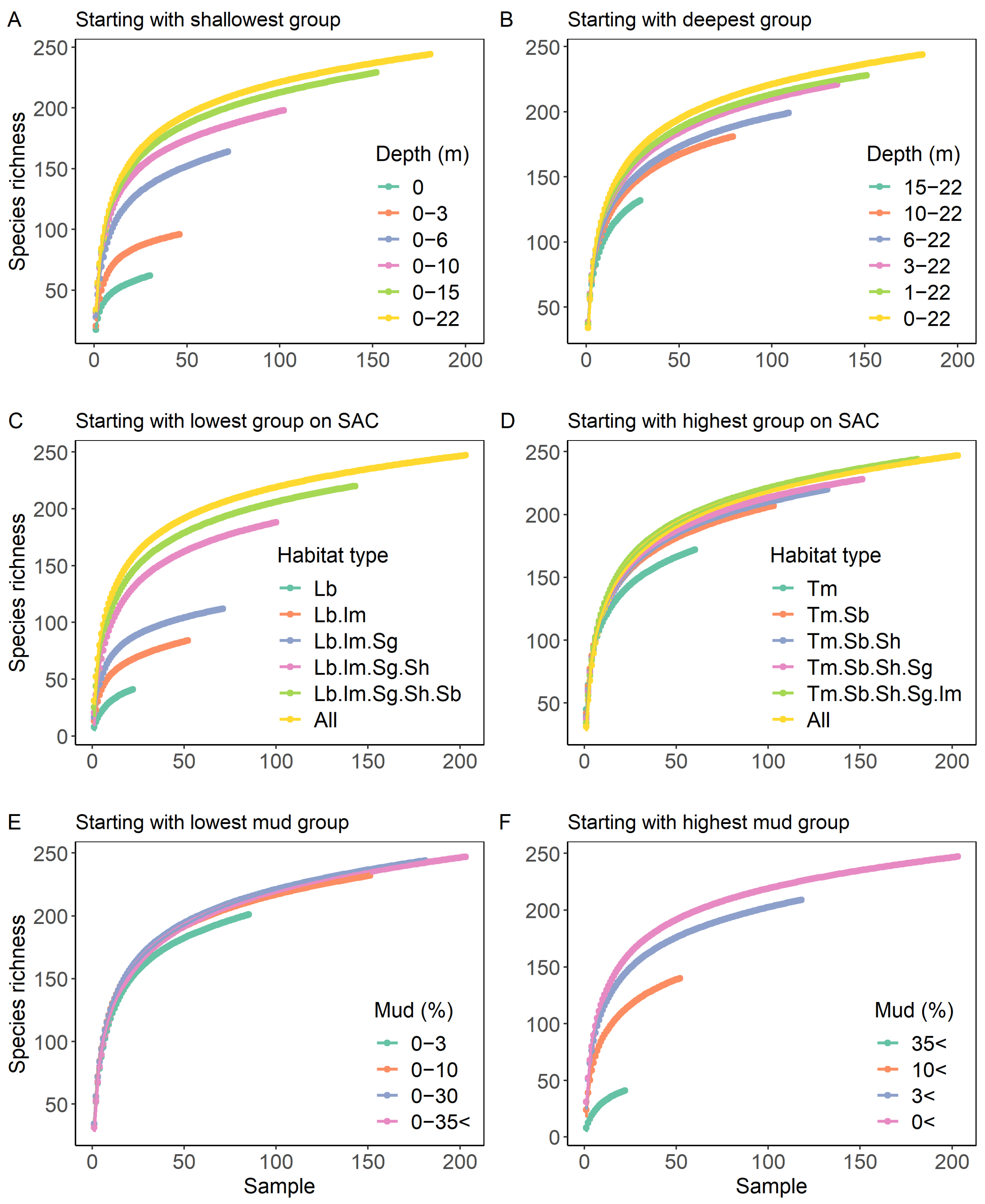
3.3. Species Accumulation in Relation to Sediment Mud Content, Water Column Depth and Habitat Type
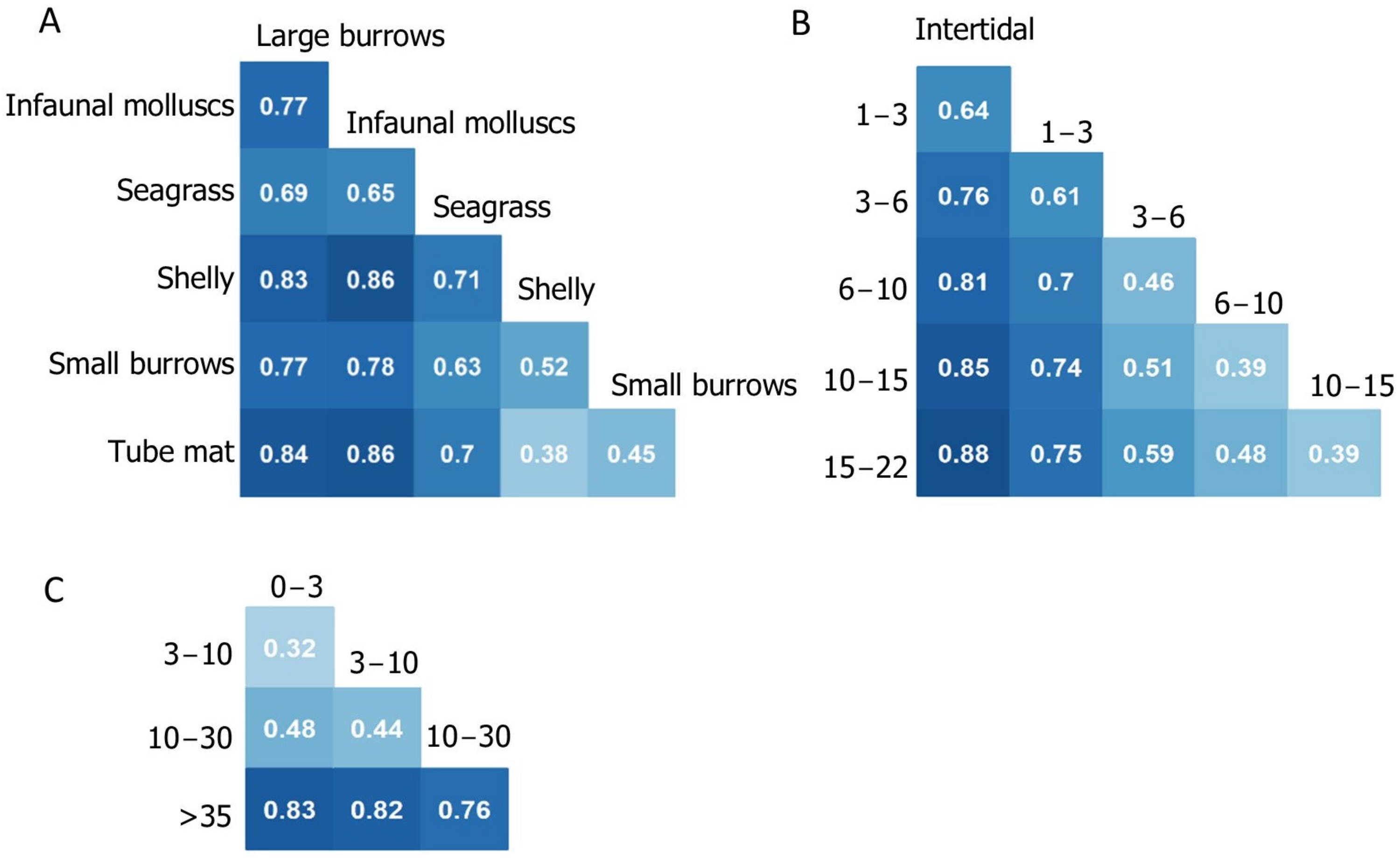
3.4. Dissimilarity Patterns within Groups
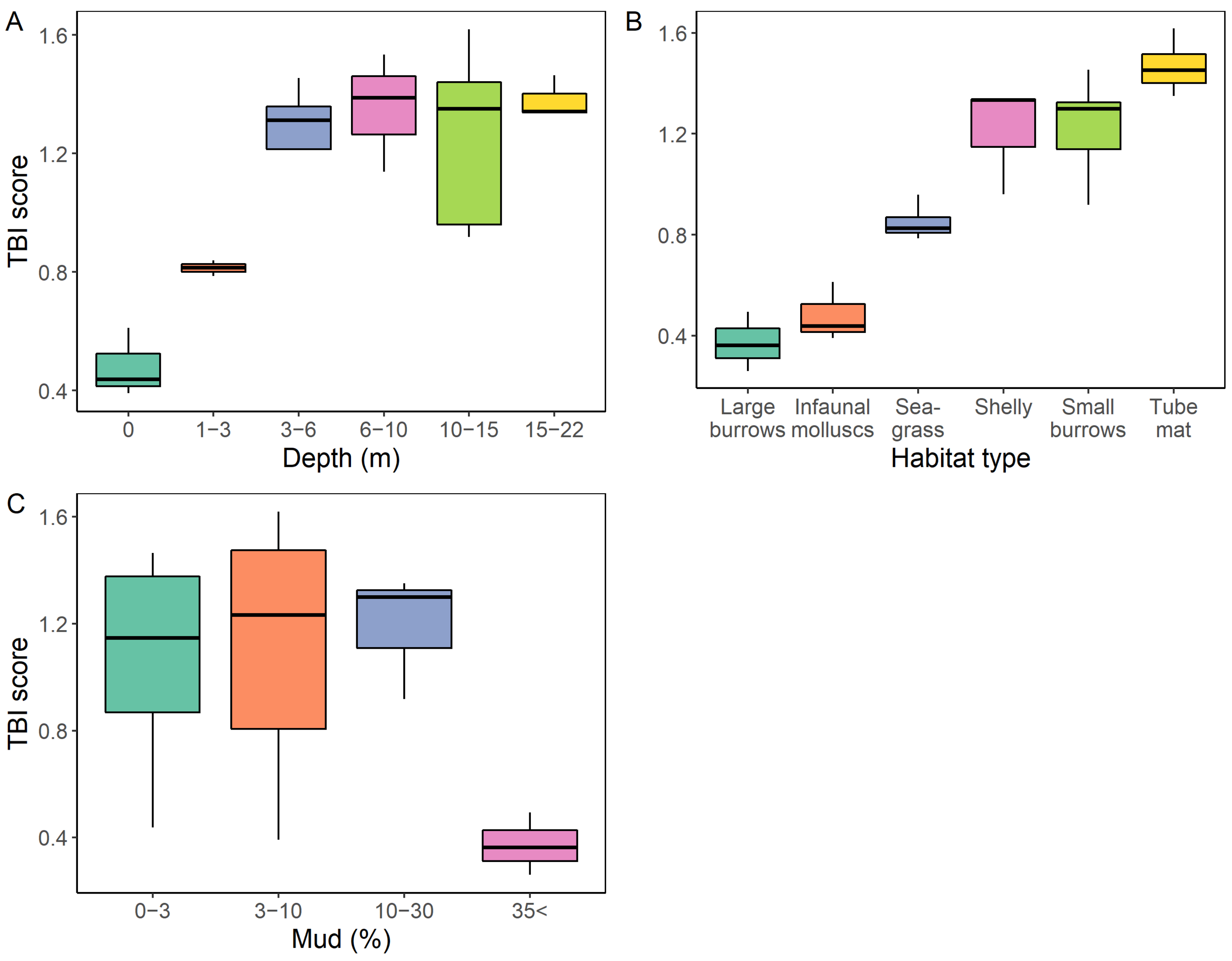
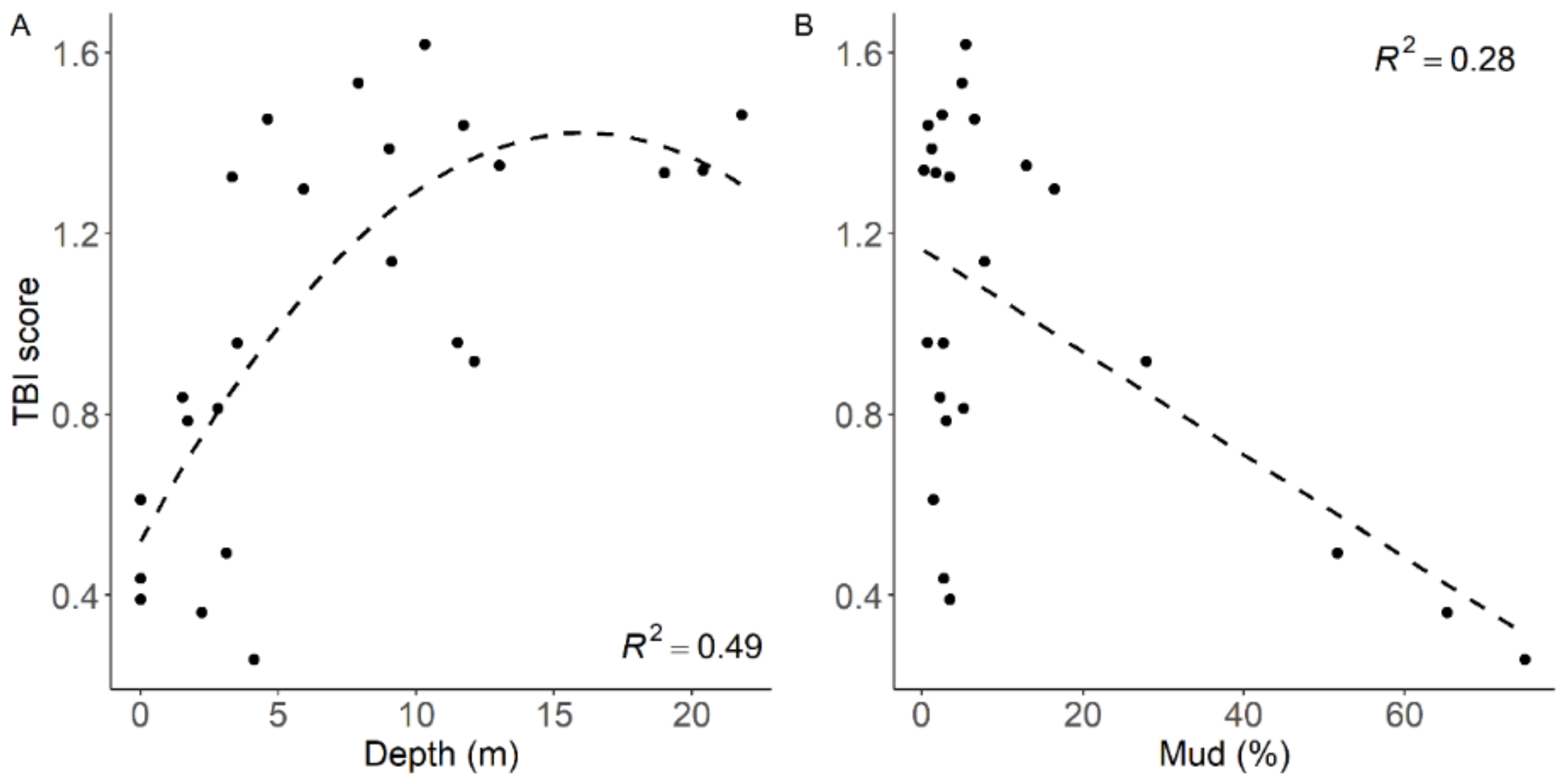
3.4.1. Water Column Depth
3.4.2. Habitat Type
3.4.3. Sediment Mud Content
3.5. Changes in Functional Redundancy between Groups
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, T.P.; Bellwood, D.R.; Folke, C.; Steneck, R.S.; Wilson, J. New paradigms for supporting the resilience of marine ecosystems. Trends Ecol. Evol. 2005, 20, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R.; McLeod, K.L.; Grünbaum, D. Ecosystems in Action: Lessons from Marine Ecology about Recovery, Resistance, and Reversibility. BioScience 2008, 58, 33–42. [Google Scholar] [CrossRef]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. A Synthesis is Emerging between Biodiversity-Ecosystem Function and Ecological Resilience Research: Reply to Mori. Trends Ecol. Evol. 2016, 31, 89–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.L.; Petchey, O.L.; et al. Biodiversity and Resilience of Ecosystem Functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, B.H. Biodiversity and Ecological Redundancy. Conserv. Biol. 1992, 6, 18–23. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Lefcheck, J.S.; Byrnes, J.E.K.; Cardinale, B.J.; Duffy, J.E.; Griffin, J.N. Marine biodiversity and ecosystem functioning: What’s known and what’s next? Oikos 2015, 124, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Albright, R.; Mason, B. Projected Near-Future Levels of Temperature and pCO2 Reduce Coral Fertilization Success. PLoS ONE 2013, 8, e56468. [Google Scholar] [CrossRef] [Green Version]
- Lenihan, H.S.; Micheli, F.; Shelton, S.W.; Peterson, C.H. The influence of multiple environmental stressors on susceptibility to parasites: An experimental determination with oysters. Limnol. Oceanogr. 1999, 44, 910–924. [Google Scholar] [CrossRef] [Green Version]
- Maltby, L. Studying stress: The importance of organism-level responses. Ecol. Appl. 1999, 9, 431–440. [Google Scholar] [CrossRef]
- Elliott, M.; Quintino, V. The Estuarine Quality Paradox, Environmental Homeostasis and the difficulty of detecting anthropogenic stress in naturally stressed areas. Mar. Pollut. Bull. 2007, 54, 640–645. [Google Scholar] [CrossRef]
- Rautenberger, R.; Bischof, K. Impact of temperature on UV-susceptibility of two Ulva (Chlorophyta) species from Antarctic and Subantarctic regions. Polar Biol. 2006, 29, 988–996. [Google Scholar] [CrossRef] [Green Version]
- Gladstone-Gallagher, R.V.; Pilditch, C.A.; Stephenson, F.; Thrush, S.F. Linking Traits across Ecological Scales Determines Functional Resilience. Trends Ecol. Evol. 2019, 34, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, B.L.; Kraan, C.; Pilditch, C.A.; Thrush, S.F. Mapping functional groups can provide insight into ecosystem functioning and potential resilience of intertidal sandflats. Mar. Ecol. Prog. Ser. 2016, 548, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Loreau, M.; Mouquet, N.; Gonzalez, A. Biodiversity as spatial insurance in heterogeneous landscapes. Proc. Natl. Acad. Sci. USA 2003, 100, 12765–12770. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes—Eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- De Juan, S.; Thrush, S.F.; Hewitt, J.E. Counting on β-Diversity to Safeguard the Resilience of Estuaries. PLoS ONE 2013, 8, e65575. [Google Scholar] [CrossRef]
- Hewitt, J.E.; Thrush, S.F.; Lohrer, A.M.; Townsend, M.J. A latent threat to biodiversity: Consequences of small-scale heterogeneity loss. Biodivers. Conserv. 2010, 19, 1315–1323. [Google Scholar] [CrossRef]
- Thrush, S.F.; Hewitt, J.E.; Cummings, V.J.; Norkko, A.; Chiantore, M. β-Diversity and Species Accumulation in Antarctic Coastal Benthos: Influence of Habitat, Distance and Productivity on Ecological Connectivity. PLoS ONE 2010, 5, e11899. [Google Scholar] [CrossRef]
- Telesh, I.V.; Khlebovich, V.V. Principal processes within the estuarine salinity gradient: A review. Mar. Pollut. Bull. 2010, 61, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Van der Wal, D.; Lambert, G.I.; Ysebaert, T.; Plancke, Y.M.G.; Herman, P.M.J. Hydrodynamic conditioning of diversity and functional traits in subtidal estuarine macrozoobenthic communities. Estuar. Coast. Shelf Sci. 2017, 197, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; van Maren, D.S.; Guo, L.; Lin, J.; He, Q.; Wang, Z.B. Effects of Sediment-Induced Density Gradients on the Estuarine Turbidity Maximum in the Yangtze Estuary. J. Geophys. Res. Ocean. 2021, 126, e2020JC016927. [Google Scholar] [CrossRef]
- De Juan, S.; Hewitt, J. Relative importance of local biotic and environmental factors versus regional factors in driving macrobenthic species richness in intertidal areas. Mar. Ecol. Prog. Ser. 2011, 423, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Peterson, C.H.; Jackson, J.B.C. Depletion, Degradation, and Recovery Potential of Estuaries and Coastal Seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- McWethy, D.B.; Whitlock, C.; Wilmshurst, J.M.; McGlone, M.S.; Fromont, M.; Li, X.; Dieffenbacher-Krall, A.; Hobbs, W.O.; Fritz, S.C.; Cook, E.R. Rapid landscape transformation in South Island, New Zealand, following initial Polynesian settlement. Proc. Natl. Acad. Sci. USA 2010, 107, 21343–21348. [Google Scholar] [CrossRef] [Green Version]
- Abrahim, G.; Parker, R. Heavy-metal contaminants in Tamaki Estuary: Impact of city development and growth, Auckland, New Zealand. Environ. Geol. 2002, 42, 883–890. [Google Scholar] [CrossRef]
- Moller, H.; MacLeod, C.J.; Haggerty, J.; Rosin, C.; Blackwell, G.; Perley, C.; Meadows, S.; Weller, F.; Gradwohl, M. Intensification of New Zealand agriculture: Implications for biodiversity. N. Z. J. Agric. Res. 2008, 51, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Swales, A.; Williamson, R.B.; Van Dam, L.F.; Stroud, M.J.; McGlone, M.S. Reconstruction of urban stormwater contamination of an estuary using catchment history and sediment profile dating. Estuaries 2002, 25, 43–56. [Google Scholar] [CrossRef]
- Hicks, D.M.; Shankar, U.; McKerchar, A.I.; Basher, L.; Lynn, I.; Page, M.; Jessen, M. Suspended sediment yields from New Zealand rivers. J. Hydrol. 2011, 50, 81–142. [Google Scholar]
- Thrush, S.F.; Hewitt, J.E.; Cummings, V.J.; Ellis, J.I.; Hatton, C.; Lohrer, A.; Norkko, A. Muddy Waters: Elevating Sediment Input to Coastal and Estuarine Habitats. Front. Ecol. Environ. 2004, 2, 299–306. [Google Scholar] [CrossRef]
- Cummings, V.; Vopel, K.; Thrush, S. Terrigenous deposits in coastal marine habitats: Influences on sediment geochemistry and behaviour of post-settlement bivalves. Mar. Ecol. Prog. Ser. 2009, 383, 173–185. [Google Scholar] [CrossRef]
- Lohrer, A.M.; Thrush, S.F.; Hewitt, J.E.; Berkenbusch, K.; Ahrens, M.; Cummings, V.J. Terrestrially derived sediment: Response of marine macrobenthic communities to thin terrigenous deposits. Mar. Ecol. Prog. Ser. 2004, 273, 121–138. [Google Scholar] [CrossRef] [Green Version]
- Pratt, D.R.; Lohrer, A.M.; Pilditch, C.A.; Thrush, S.F. Changes in Ecosystem Function Across Sedimentary Gradients in Estuaries. Ecosystems 2014, 17, 182–194. [Google Scholar] [CrossRef]
- Robertson, B.P.; Gardner, J.P.A.; Savage, C. Macrobenthic–mud relations strengthen the foundation for benthic index development: A case study from shallow, temperate New Zealand estuaries. Ecol. Indic. 2015, 58, 161–174. [Google Scholar] [CrossRef]
- Rodil, I.F.; Lohrer, A.M.; Chiaroni, L.D.; Hewitt, J.E.; Thrush, S.F. Disturbance of sandflats by thin terrigenous sediment deposits: Consequences for primary production and nutrient cycling. Ecol Appl 2011, 21, 416–426. [Google Scholar] [CrossRef]
- Swales, A.; Gibbs, M.; Pritchard, M.; Budd, R.; Olsen, G.; Ovenden, R.; Costley, K.; Hermanspahn, N.; Griffiths, R. Whangarei Harbour Sedimentation: Sediment Accumulation Rates and Present-Day Sediment Sources; National Institute of Water and Atmospheric Research: Auckland, New Zealand, 2013; p. 40. [Google Scholar]
- Bulmer, R.H.; Schwendenmann, L.; Lohrer, A.M.; Lundquist, C.J. Sediment carbon and nutrient fluxes from cleared and intact temperate mangrove ecosystems and adjacent sandflats. Sci. Total Environ. 2017, 599–600, 1874–1884. [Google Scholar] [CrossRef]
- Cummings, V.J.; Thrush, S.F. Behavioural response of juvenile bivalves to terrestrial sediment deposits: Implications for post-disturbance recolonisation. Mar. Ecol. Prog. Ser. 2004, 278, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Hohaia, A.; Vopel, K.; Pilditch, C.A. Thin terrestrial sediment deposits on intertidal sandflats: Effects on pore-water solutes and juvenile bivalve burial behaviour. Biogeosciences 2014, 11, 2225–2235. [Google Scholar] [CrossRef] [Green Version]
- Norkko, A.; Thrush, S.F.; Hewitt, J.E.; Cummings, V.J.; Norkko, J.; Ellis, J.I.; Funnell, G.A.; Schultz, D.; MacDonald, I. Smothering of estuarine sandflats by terrigenous clay: The role of wind-wave disturbance and bioturbation in site-dependent macrofaunal recovery. Mar. Ecol. Prog. Ser. 2002, 234, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Thrush, S.F.; Hewitt, J.; Lohrer, A. Interaction networks in coastal soft-sediments highlight the potential for change in ecological resilience. Ecol. Appl. A Publ. Ecol. Soc. Am. 2012, 22, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Thrush, S.F.; Hewitt, J.E.; Norkko, A.; Cummings, V.J.; Funnell, G.A. Macrobenthic Recovery Processes Following Catastrophic Sedimentation on Estuarine Sandflats. Ecol. Appl. 2003, 13, 1433–1455. [Google Scholar] [CrossRef]
- Cummings, V.; Thrush, S.F.; Hewitt, J.; Norkko, A.; Pickmere, S. Terrestrial deposits on intertidal sandflats: Sediment characteristics as indicators of habitat suitability for recolonising macrofauna. Mar. Ecol. Prog. Ser. 2003, 253, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.; Cummings, V.; Hewitt, J.; Thrush, S.; Norkko, A. Determining effects of suspended sediment on condition of a suspension feeding bivalve (Atrina zelandica): Results of a survey, a laboratory experiment and a field transplant experiment. J. Exp. Mar. Biol. Ecol. 2002, 267, 147–174. [Google Scholar] [CrossRef]
- Ugland, K.I.; Gray, J.S.; Ellingsen, K.E. The species–accumulation curve and estimation of species richness. J. Anim. Ecol. 2003, 72, 888–897. [Google Scholar] [CrossRef] [Green Version]
- Crist, T.O.; Veech, J.A.; Gering, J.C.; Summerville, K.S. Partitioning species diversity across landscapes and regions: A hierarchical analysis of alpha, beta, and gamma diversity. Am. Nat. 2003, 162, 734–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gering, J.C.; Crist, T.O.; Veech, J.A. Additive Partitioning of Species Diversity across Multiple Spatial Scales: Implications for Regional Conservation of Biodiversity. Conserv. Biol. 2003, 17, 488–499. [Google Scholar] [CrossRef]
- Ricotta, C. Computing additive beta-diversity from presence and absence scores: A critique and alternative parameters. Popul. Biol. 2008, 73, 244–249. [Google Scholar] [CrossRef]
- Rodil, I.F.; Lohrer, A.M.; Hewitt, J.E.; Townsend, M.; Thrush, S.F.; Carbines, M. Tracking environmental stress gradients using three biotic integrity indices: Advantages of a locally-developed traits-based approach. Ecol. Indic. 2013, 34, 560–570. [Google Scholar] [CrossRef]
- Hewitt, J.E.; Anderson, M.J.; Hickey, C.W.; Kelly, S.; Thrush, S.F. Enhancing the Ecological Significance of Sediment Contamination Guidelines through Integration with Community Analysis. Environ. Sci. Technol. 2009, 43, 2118–2123. [Google Scholar] [CrossRef]
- Hewitt, J.E.; Thrush, S.F.; Halliday, J.; Duffy, C. The importance of small-scale habitat structure for maintaining beta diversity. Ecology 2005, 86, 1619–1626. [Google Scholar] [CrossRef]
- Thrush, S.F.; Hewitt, J.E.; Hickey, C.W.; Kelly, S. Multiple stressor effects identified from species abundance distributions: Interactions between urban contaminants and species habitat relationships. J. Exp. Mar. Biol. Ecol. 2008, 366, 160–168. [Google Scholar] [CrossRef]
- Ministry for the Environment; Stats NZ. New Zealand’s Environmental Reporting Series: Our Land 2018; Ministry for the Environment and Stats NZ: Auckland, New Zealand, 2018.
- Marinelli, R.L.; Woodin, S.A. Experimental evidence for linkages between infaunal recruitment, disturbance, and sediment surface chemistry. Limnol. Oceanogr. 2002, 47, 221–229. [Google Scholar] [CrossRef]
- Douglas, E.J.; Lohrer, A.M.; Pilditch, C.A. Biodiversity breakpoints along stress gradients in estuaries and associated shifts in ecosystem interactions. Sci. Rep. 2019, 9, 17567. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S. Species richness of marine soft sediments. Mar. Ecol. Prog. Ser. 2002, 244, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Aller, J.Y. Quantifying sediment disturbance by bottom currents and its effect on benthic communities in a deep-sea western boundary zone. Deep Sea Res. Part A Oceanogr. Res. Pap. 1989, 36, 901–934. [Google Scholar] [CrossRef]
- Turner, S.J.; Grant, J.; Pridmore, R.D.; Hewitt, J.E.; Wilkinson, M.R.; Hume, T.M.; Morrisey, D.J. Bedload and water-column transport and colonization processes by post-settlement benthic macrofauna: Does infaunal density matter? J. Exp. Mar. Biol. Ecol. 1997, 216, 51–75. [Google Scholar] [CrossRef]
- Thrush, S.F.; Hewitt, J.E.; Gibbs, M.; Lundquist, C.; Norkko, A. Functional Role of Large Organisms in Intertidal Communities: Community Effects and Ecosystem Function. Ecosystems 2006, 9, 1029–1040. [Google Scholar] [CrossRef]
- Berkenbusch, K.; Rowden, A.A. An examination of the spatial and temporal generality of the influence of ecosystem engineers on the composition of associated assemblages. Aquat. Ecol. 2007, 41, 129–147. [Google Scholar] [CrossRef]
- Hewitt, J.E.; Norkko, J. Incorporating temporal variability of stressors into studies: An example using suspension-feeding bivalves and elevated suspended sediment concentrations. J. Exp. Mar. Biol. Ecol. 2007, 341, 131–141. [Google Scholar] [CrossRef]
- Widdows, J.; Brinsley, M.D.; Salkeld, P.N.; Elliott, M. Use of annular flumes to determine the influence of current velocity and bivalves on material flux at the sediment-water interface. Estuaries 1998, 21, 552–559. [Google Scholar] [CrossRef]
- Turner, S.J.; Thrush, S.F.; Hewitt, J.E.; Cummings, V.J.; Funnell, G. Fishing impacts and the degradation or loss of habitat structure. Fish. Manag. Ecol. 1999, 6, 401–420. [Google Scholar] [CrossRef]
- Bremner, J.; Rogers, S.I.; Frid, C.L.J. Assessing functional diversity in marine benthic ecosystems: A comparison of approaches. Mar. Ecol. Prog. Ser. 2003, 254, 11–25. [Google Scholar] [CrossRef]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The Influence of Functional Diversity and Composition on Ecosystem Processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, R.J.; Cazenave, A. Sea-Level Rise and Its Impact on Coastal Zones. Science 2010, 328, 1517–1520. [Google Scholar] [CrossRef]
- Seneviratne, S.; Nicholls, N.; Easterling, D.; Goodess, C.; Kanae, S.; Kossin, J.; Luo, Y.; Marengo, J.; McInnes, K.; Rahimi, M.; et al. Changes in climate extremes and their impacts on the natural physical environment. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; Field, C.B., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Plattner, G.-K., Allen, S.K., et al., Eds.; A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC); Cambridge University Press: Cambridge, UK; New York, NY, USA, 2012; pp. 109–230. [Google Scholar]
- Bulmer, R.H.; Kelly, S.; Jeffs, A.G. Light requirements of the seagrass, Zostera muelleri, determined by observations at the maximum depth limit in a temperate estuary, New Zealand. N. Z. J. Mar. Freshw. Res. 2016, 50, 183–194. [Google Scholar] [CrossRef]
- Drylie, T.P.; Lohrer, A.M.; Needham, H.R.; Bulmer, R.H.; Pilditch, C.A. Benthic primary production in emerged intertidal habitats provides resilience to high water column turbidity. J. Sea Res. 2018, 142, 101–112. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. Sites (No. Cores) | Habitat (No. Sites) | Mud (%) (No. Sites) | Depth (m) (No. Sites) | |
|---|---|---|---|---|
| Habitat | ||||
| Large burrows | 3 (22) | >35 (3) | 1–3 (1), 3–6 (2) | |
| Infaunal molluscs | 3 (30) | 0–3 (2), 3–10 (1) | Intertidal (3) | |
| Seagrass | 4 (19) | 0–3 (2), 3–10 (2) | 1–3 (3), 3–6 (1) | |
| Shelly | 3 (29) | 0–3 (3) | 10–15 (1), 15–22 (2) | |
| Small burrows | 5 (43) | 3–10 (3), 10–30 (2) | 3–6 (3), 6–10 (1), 10–15 (1) | |
| Tube mat | 6 (60) | 0–3 (3), 3–10 (2), 10–30 (1) | 6–10 (2), 10–15 (3), 15–22 (1) | |
| Sediment mud content (%) | ||||
| 0–3 | 10 (85) | Infaunal molluscs (2), seagrass (2), shelly (3), tube mat (3) | Intertidal (2), 1–3 (1), 3–6 (1), 6–10 (1), 10–15 (2), 15–22 (3) | |
| 3–10 | 8 (66) | Infaunal molluscs (1), seagrass (2), small burrows (3), tube mat (2) | Intertidal (1), 1–3 (2), 3–6 (2), 6–10 (2), 10–15 (1) | |
| 10–30 | 3 (30) | Small burrows (2), tube mat (1) | 3–6 (1), 10–15 (2) | |
| >35 | 3 (22) | Large burrows (3) | 1–3 (1), 3–6 (2) | |
| Depth (m) | ||||
| Intertidal | 3 (30) | Infaunal molluscs (3) | 0–3 (2), 3–10 (1) | |
| 1–3 | 3 (16) | Seagrass (3) | 0–3 (1), 3–10 (2) | |
| 3–6 | 4 (26) | Small burrows (3), seagrass (1) | 0–3 (1), 3–10 (2), 10–30 (1) | |
| 6–10 | 3 (30) | Small burrows (1), tube mat (2) | 0–3 (1), 3–10 (2) | |
| 10–15 | 5 (50) | Small burrows (1), shelly (1), tube mat (3) | 0–3 (2), 3–10 (1), 10–30 (2) | |
| 15–22 | 3 (29) | Shelly (2), tube mat (1) | 0–3 (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mangan, S.; Bulmer, R.H.; Greenfield, B.L.; Hailes, S.F.; Carter, K.; Hewitt, J.E.; Lohrer, A.M. Resilience and Species Accumulation across Seafloor Habitat Transitions in a Northern New Zealand Harbour. Diversity 2022, 14, 998. https://doi.org/10.3390/d14110998
Mangan S, Bulmer RH, Greenfield BL, Hailes SF, Carter K, Hewitt JE, Lohrer AM. Resilience and Species Accumulation across Seafloor Habitat Transitions in a Northern New Zealand Harbour. Diversity. 2022; 14(11):998. https://doi.org/10.3390/d14110998
Chicago/Turabian StyleMangan, Stephanie, Richard H. Bulmer, Barry L. Greenfield, Sarah F. Hailes, Kelly Carter, Judi E. Hewitt, and Andrew M. Lohrer. 2022. "Resilience and Species Accumulation across Seafloor Habitat Transitions in a Northern New Zealand Harbour" Diversity 14, no. 11: 998. https://doi.org/10.3390/d14110998