
Relationship between Biomass and Biodiversity of Degraded Grassland in the Sanjiangyuan Region of Qinghai–Tibet Plateau

Abstract
:1. Introduction
2. Materials and Methods
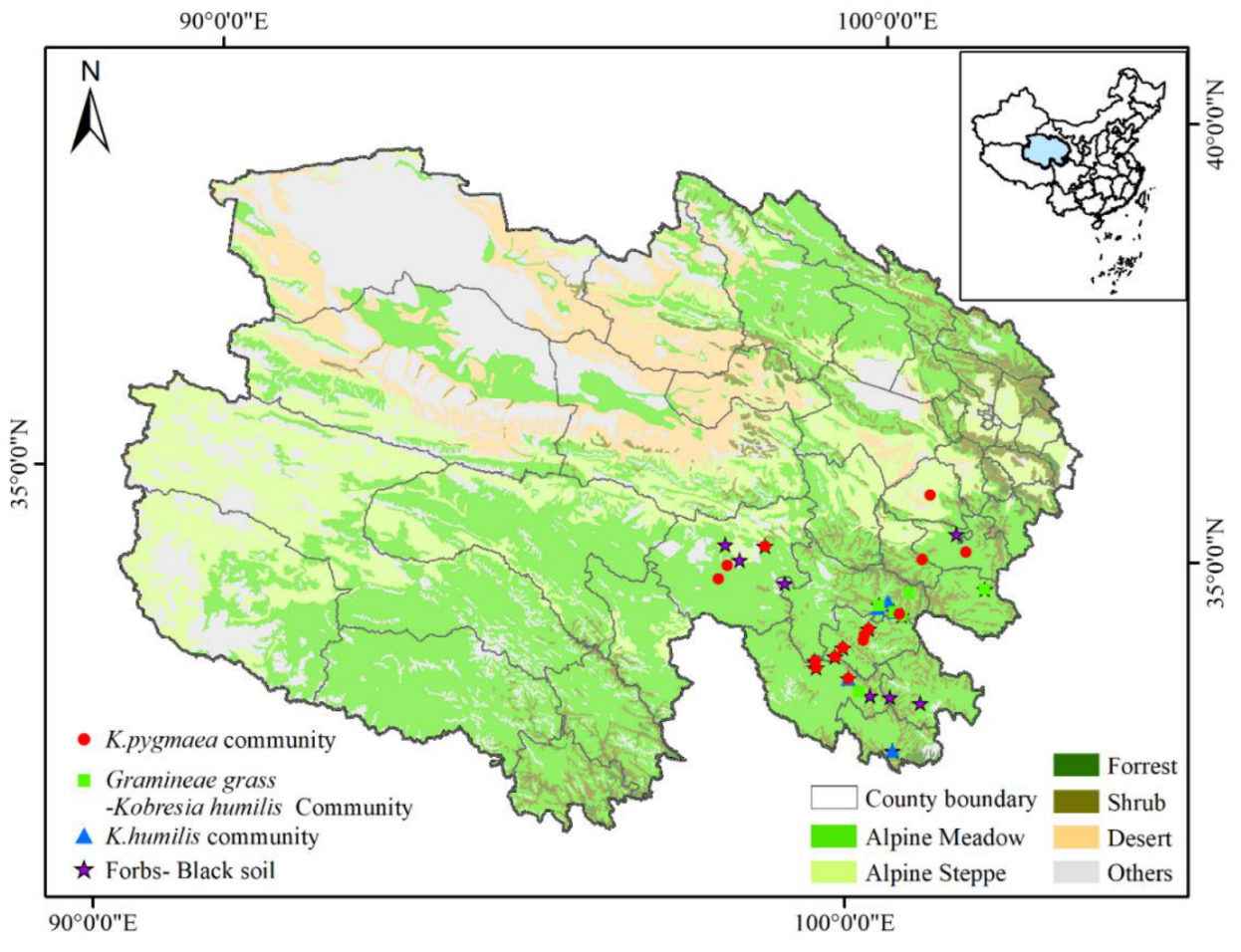
2.1. Study Area
2.2. Experimental Design
2.3. Measures of Biodiversity
- (i).
- Richness index (R), the number of vascular species recorded in each plot;
- (ii).
- Shannon-Wiener Diversity index(H), calculated as:
- (iii).
- Pielou evenness index (J), calculated as:where Pi = Ni/N, Ni is the importance value of species i (which is calculated based on plant coverage), and N is the total importance value of all vascular species present.
2.4. Statistical Analysis
3. Results
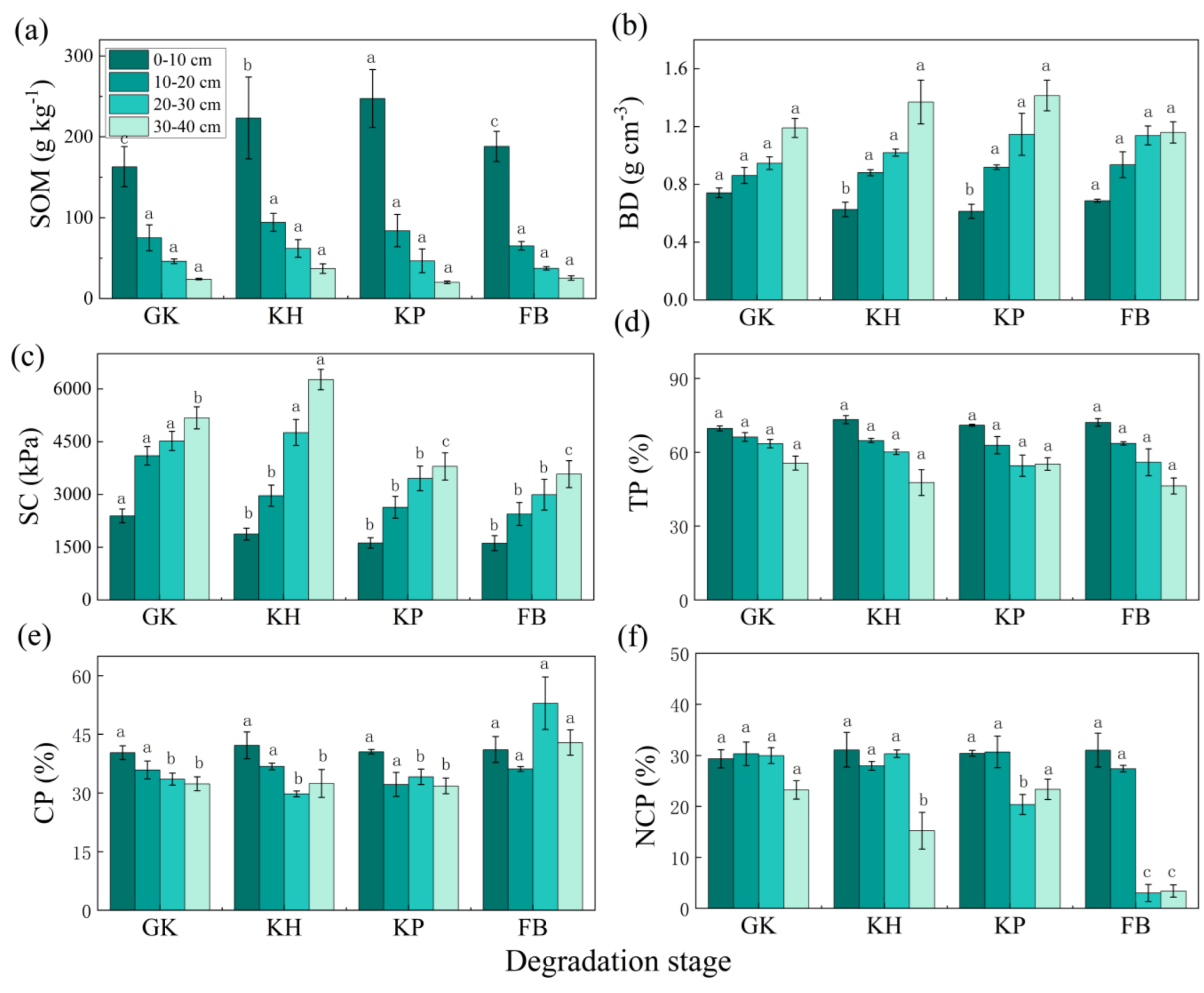
3.1. Soil Properties of the Multi-Stable Alpine Meadows
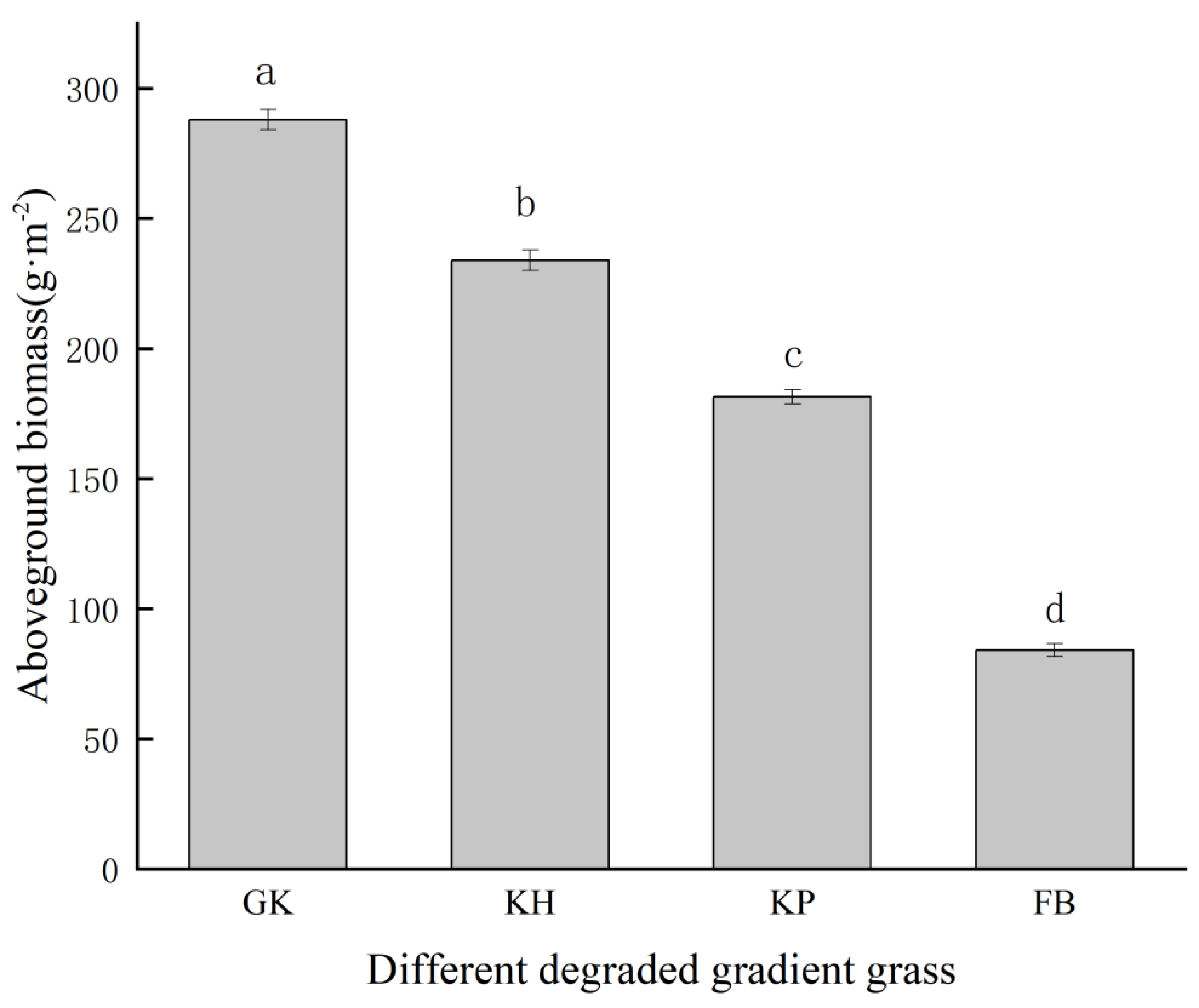
3.2. Aboveground Biomass of the Multi-Stable Alpine Meadows
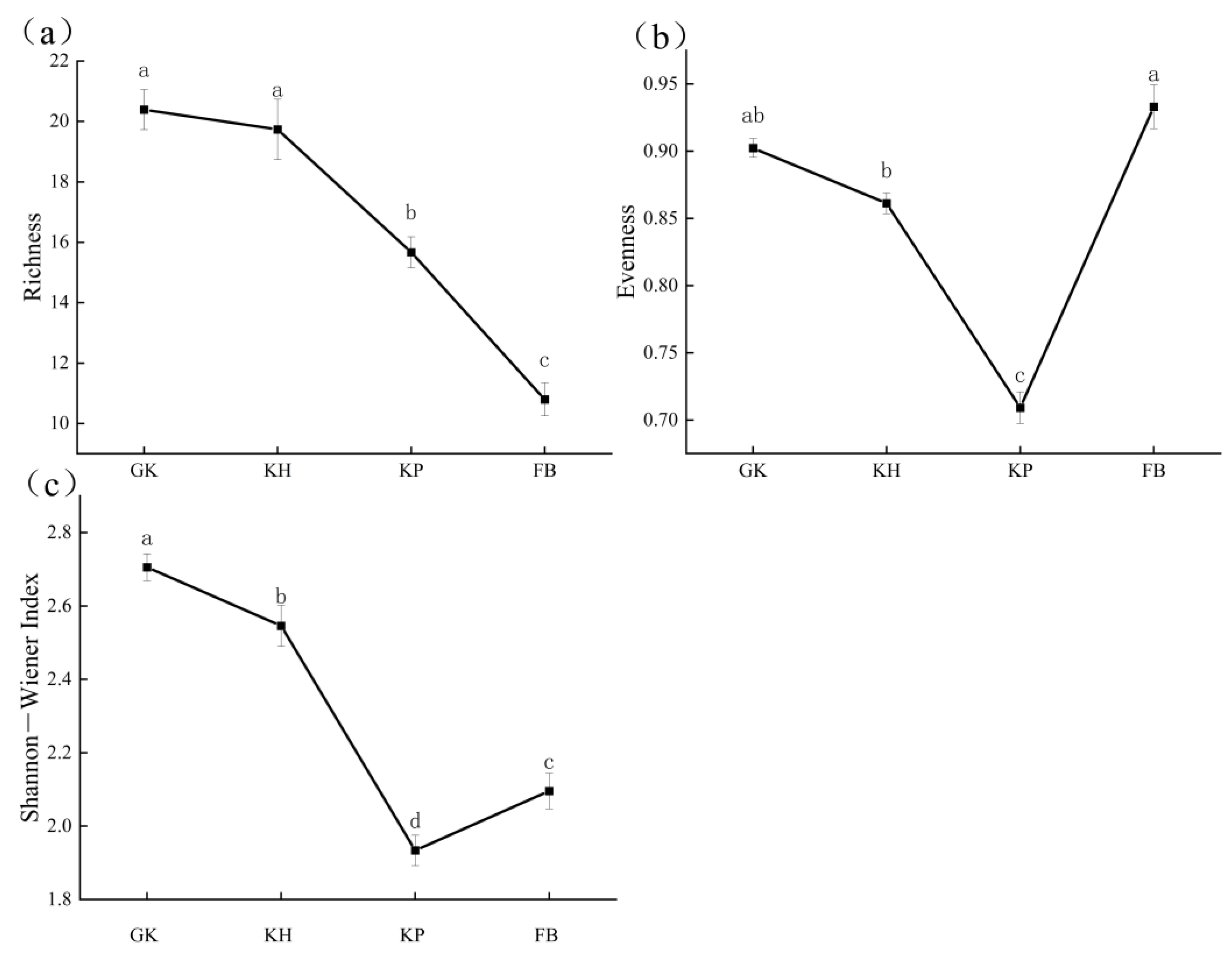
3.3. Biodiversity of the Multi-Stable Alpine Meadows
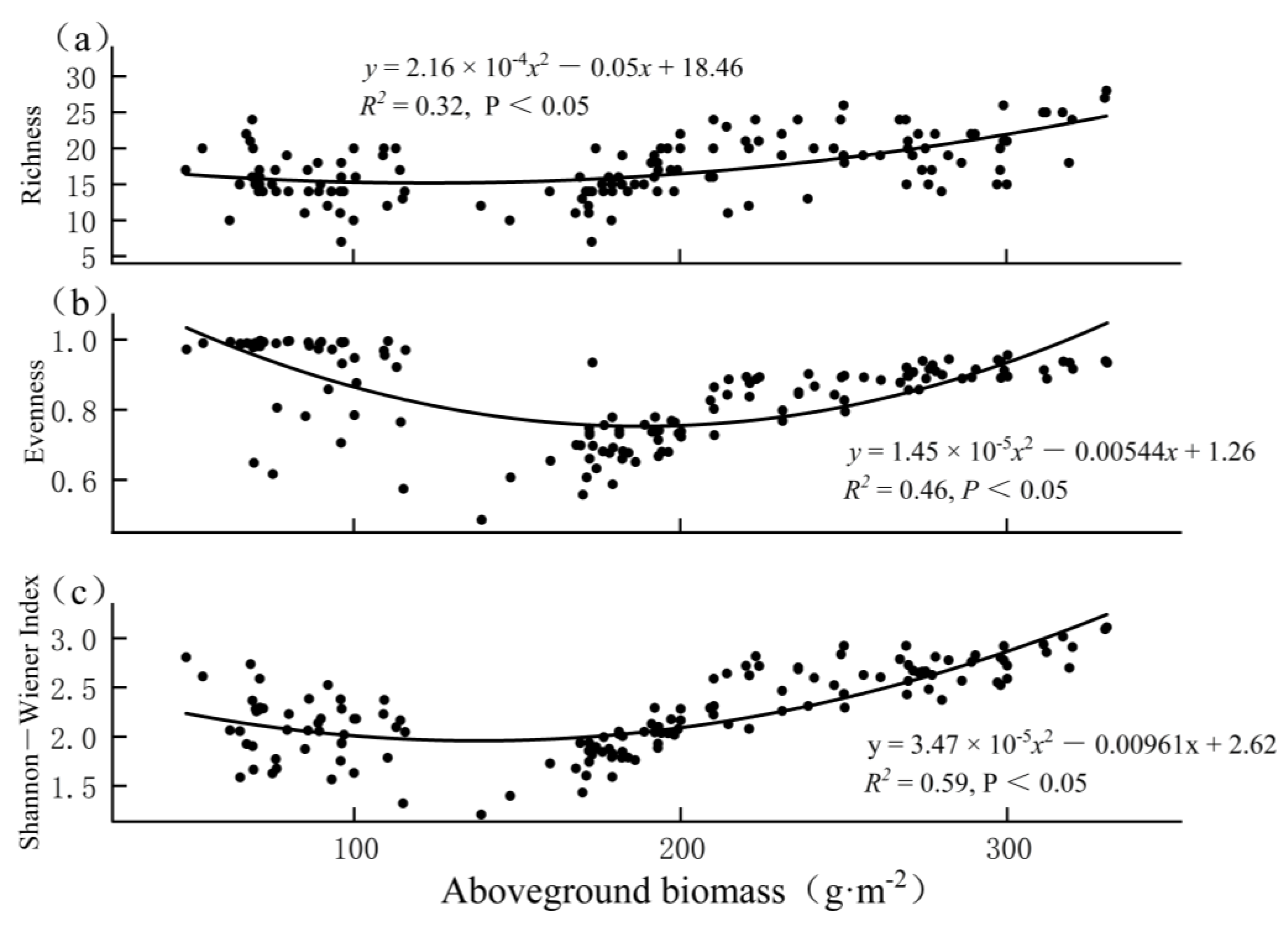
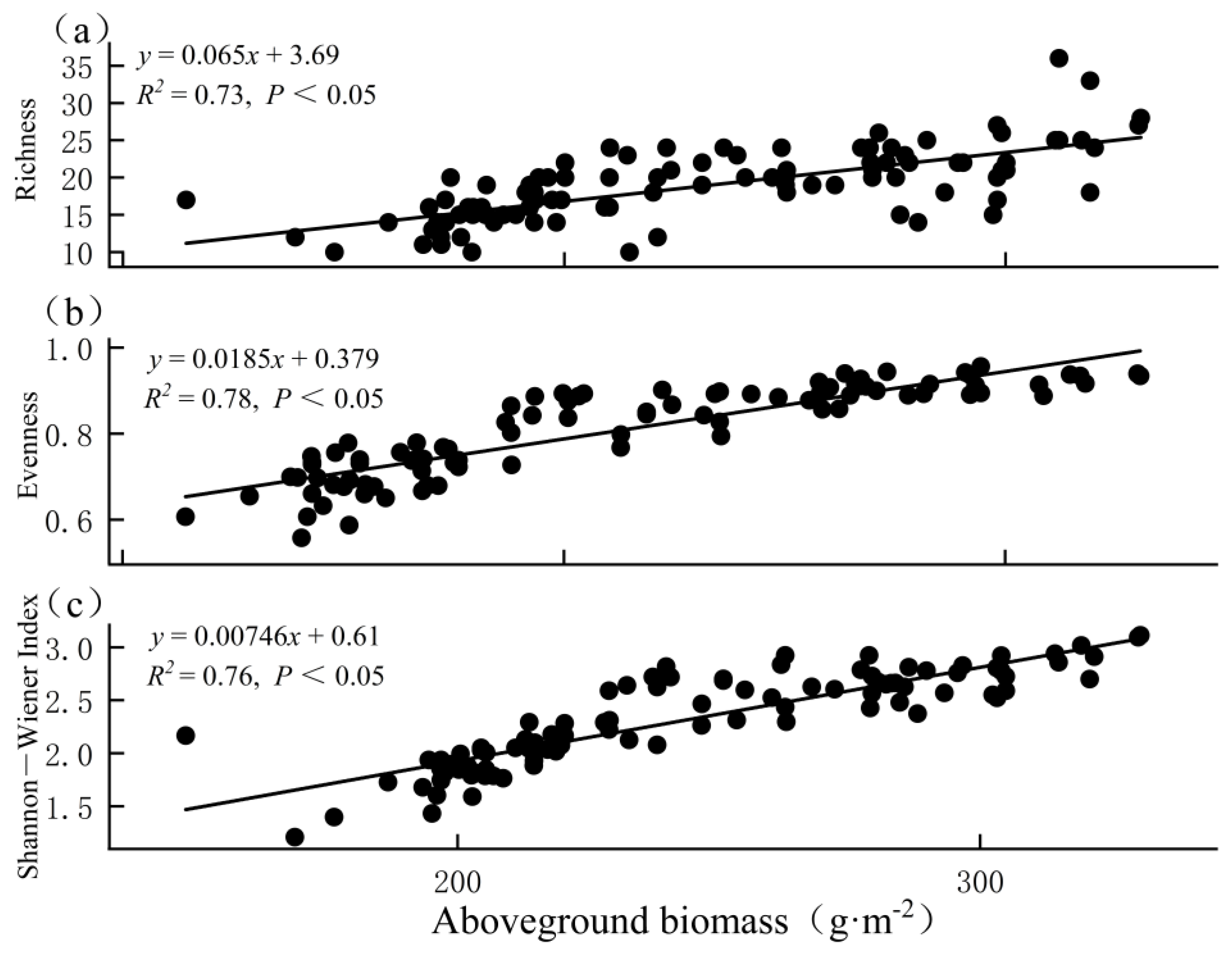
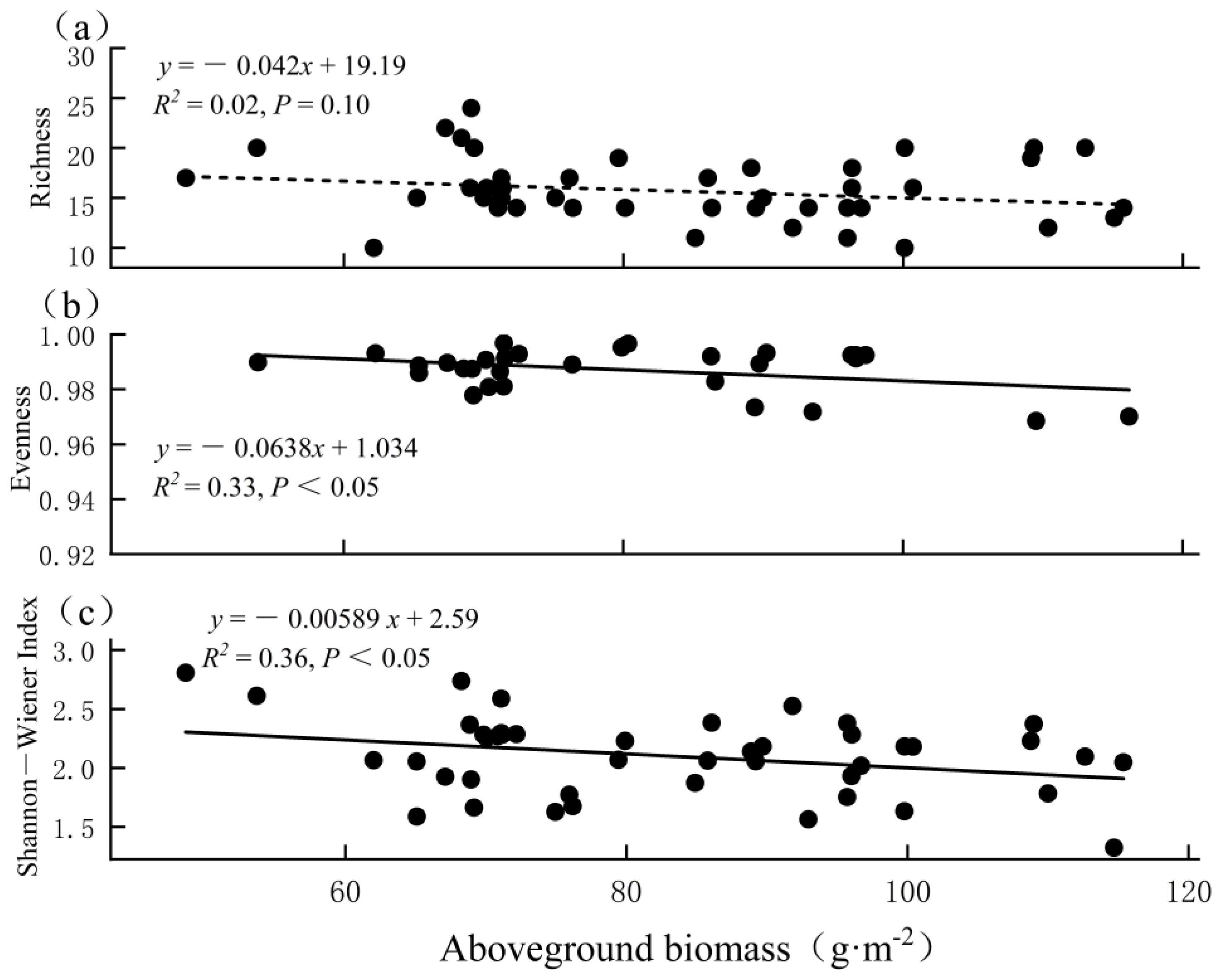
3.4. Relationship between Biodiversity and Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, R.; Long, R.; Shang, Z.; Ma, Y.; Dong, S.; Wang, Y. Establishment of Elymus natans improves soil quality of a heavily degraded alpine meadow in Qinghai-Tibetan Plateau, China. Plant Soil 2010, 327, 403–411. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Ji, C.; Han, W. Above- and belowground biomass allocation in Tibetan grasslands. J. Veg. Sci. 2009, 20, 177–184. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity, and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef] [Green Version]
- Akatov, V.V.; Akatova, T.V.; Chefranov, S.G. Relationship between Degree of Dominance and Species Richness in Grass Communities with Different Productivities. Contemp. Probl. Ecol. 2018, 11, 355–365. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Gross, K.; Fritschie, K.; Flombaum, P.; Fox, J.W.; Rixen, C.; Ruijven, J.V.; Reich, P.B.; Scherer-Lorenzen, M.; Wilsey, B.J. Biodiversity simultaneously enhances the production and stability of community biomass, but the effects are Independent. Ecology 2013, 94, 1697–1707. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; De Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar]
- Liu, Y.X.; Jian, W.H.; Qin, Y.; Xue-Fei, X.U.; Zhang, Y.Z. The relationship between biomass and biodiversity in the forest of Larix principis-rupprechtii. Hebei J. For. Orchard. Res. 2010, 25, 225–227. [Google Scholar]
- Bhattarai, K.R.; Vetaas, O.R.; Grytnes, J.A. Relationship between plant species richness and biomass in an arid sub-alpine grassland of the central Himalayas, Nepal. Folia Geobot. 2004, 39, 57–71. [Google Scholar] [CrossRef]
- Liira, J.; Zobel, K. The Species Richness-Biomass Relationship in Herbaceous Plant Communities: What Difference Does the Incorporation of Root Biomass Data Make? Oikos 2010, 91, 109–114. [Google Scholar] [CrossRef]
- Collins, S.L.; Glenn, J.M.; Briggs, S.M. Effect of local and regional processes on plant species richness in tallgrass prairie. Oikos 2002, 99, 571–579. [Google Scholar] [CrossRef]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary Production of the Central Grassland Region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Cingolani, A.M.; Noy-Meir, I.; Díaz, S. Grazing Effects on Rangeland Diversity: A Synthesis of Contemporary Models. Ecol. Appl. 2005, 15, 757–773. [Google Scholar] [CrossRef]
- Collins, S.L.; Knapp, A.K.; Briggs, J.M.; Blair, J.M.; Steinauer, E.M. Modulation of diversity by grazing and mowing in native tallgrass prairie. Science 1998, 280, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Filazzola, A.; Brown, C.; Dettlaff, M.A.; Batbaatar, A.; Grenke, J.; Bao, T.; Peetoom Heida, I.; Cahill, J.F., Jr. The effects of livestock grazing on biodiversity are multi-trophic: A meta-analysis. Ecol. Lett. 2020, 23, 1298–1309. [Google Scholar] [CrossRef]
- Wang, L.; Delgado-Baquerizo, M.; Wang, D.; Isbell, F.; Liu, J.; Feng, C.; Liu, J.; Zhong, Z.; Zhu, H.; Yuan, X.; et al. Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proc. Natl. Acad. Sci. USA 2019, 116, 6187–6192. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Dong, S.; Li, Y.; Li, X.; Shi, J.; Wang, Y.; Liu, D.; Ma, Y. Effect of Degradation Intensity on Grassland Ecosystem Services in the Alpine Region of Qinghai-Tibetan Plateau, China. PLoS ONE 2013, 8, e58432. [Google Scholar] [CrossRef] [Green Version]
- Miehe, G.; Schleuss, P.M.; Seeber, E.; Babel, W.; Biermann, T.; Braendle, M.; Chen, F.; Coners, H.; Foken, T.; Gerken, T.; et al. The Kobresia pygmaea ecosystem of the Tibetan highlands—Origin, functioning and degradation of the world’s largest pastoral alpine ecosystem: Kobresia pastures of Tibet. Sci. Total Environ. 2019, 648, 754–771. [Google Scholar] [CrossRef]
- Kefi, S.; Rietkerk, M.; van Baalen, M.; Loreau, M. Local facilitation, bistability and transitions in arid ecosystems. Theor. Popul. Biol. 2007, 71, 367–379. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Gornish, E.S.; Hu, G.; Wan, Y.; Li, Y.; Danjiu, L.; Gao, Q. Temperature leads to annual changes of plant community composition in alpine grasslands on the Qinghai-Tibetan Plateau. Environ. Monit. Assess 2018, 190, 585. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Long, R. Formation causes and recovery of the “Black Soil Type” degraded alpine grassland in Qinghai-Tibetan Plateau. Front. Agric. China 2007, 1, 197–202. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, H.; Zhao, X.; Yao, B.; Ma, Z.; Dong, Q.; Zhang, Z.; Wang, W.; Yang, Y. Relationship between biodiversity and ecosystem functioning in alpine meadows of the Qinghai-Tibet Plateau. Biodivers. Sci. 2018, 26, 111–129. [Google Scholar] [CrossRef]
- Zobel, K.J. A scale-independent approach to the richness vs biomass relationship in ground-layer plant communities. Oikos 1997, 80, 325–332. [Google Scholar] [CrossRef]
- Tilman, D. The Influence of Functional Diversity and Composition on Ecosystem Processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.T.; Long, R.J. The effects of differences in functional group diversity and composition on plant community productivity in four types of alpine meadow communities. Chin. Biodivers. 2004, 12, 403–409. [Google Scholar]
- Xu, Q.; Yang, X.; Yan, Y.; Wang, S.; Loreau, M.; Jiang, L. Consistently positive effect of species diversity on ecosystem, but not population, temporal stability. Ecol. Lett. 2021, 24, 2256–2266. [Google Scholar] [CrossRef]
- Abrams, P.A. Resource Productivity-Consumer Species Diversity: Simple Models of Competition in Spatially Heterogeneous Environments. Ecology 1988, 69, 1433. [Google Scholar] [CrossRef]
- Liang, M.; Liang, C.; Hautier, Y.; Wilcox, K.R.; Wang, S. Grazing-induced biodiversity loss impairs grassland ecosystem stability at multiple scales. Ecol. Lett. 2021, 24, 2054–2064. [Google Scholar] [CrossRef] [PubMed]
- Gross, K.; Cardinale, B.J.; Fox, J.W.; Gonzalez, A.; Loreau, M.; Wayne Polley, H.; Reich, P.B.; van Ruijven, J. Species richness and the temporal stability of biomass production: A new analysis of recent biodiversity experiments. Am. Nat. 2014, 183, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Li, Y.; Härdtle, W.; Von Oheimb, G.; Yang, X.; et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, C.P.; Bazeley-White, E.; Dimitrakopoulos, P.G.; Hector, A.; Scherer-Lorenzen, M.; Schmid, B. Species evenness and productivity in experimental plant communities. Oikos 2004, 107, 50–63. [Google Scholar] [CrossRef]
- Briske, D.D.; Fuhlendorf, S.D.; Smeins, F.E. State-and-Transition Models, Thresholds, and Rangeland Health: A Synthesis of Ecological Concepts and Perspectives. Rangel. Ecol. Manag. 2005, 58, 1–10. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime Shifts, Resilience, and Biodiversity in Ecosystem Management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Yu, L.; Zhao, L. Ecological Networks in Agroecosystems: Approaches and Applications. Front. Agric. Sci. Eng. 2022, 9, 523–535. [Google Scholar]
- Ma, Z.; Liu, H.; Mi, Z.; Zhang, Z.; Wang, Y.; Xu, W.; Jiang, L.; He, J.S. Climate warming reduces the temporal stability of plant community biomass production. Nat. Commun. 2017, 8, 15378. [Google Scholar] [CrossRef] [Green Version]
- Schnabel, F.; Liu, X.; Kunz, M.; Barry, K.E.; Bongers, F.J.; Bruelheide, H.; Fichtner, A.; Härdtle, W.; Li, S.; Pfaff, C.T.; et al. Species richness stabilizes productivity via asynchrony and drought-tolerance diversity in a large-scale tree biodiversity experiment. Sci. Adv. 2021, 7, eabk1643. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Succession Stage | Grazing Intensity (Sheep Unit/ha) | Dominant Species | Plot General Situation |
|---|---|---|---|
| Gramineae grass–Kobresia humilis community (GK) | 4 | Elymus nutans, Poa sp. and Festuca rubra | The total coverage of plant communities was 96.7–92.7% and the ground was relatively flat. Almost no rodent activity. The thickness of the sod layer was around 1.66 cm. |
| K. humilis community (KH) | 8 | K. humilis, (subdominant species: E. nutans and F. rubra) | The total coverage of grass was 82.6–88.7%, the ground was flat, the rodent activity was less, and the thickness of the sod layer was around 2.13 cm. |
| K. pygmaea community (KP) | 12 | K. pygmaea | The total coverage of grass was 35.8–75.8%; black spots and cracks were visible on the surface. The bark was uneven, the rodent activity was frequent, and the thickness of the sod layer was about 3.0 cm. |
| Forbs-Black Soil (FB) | 16 | Ligularia sibirica, Clematis florida, and Rheum palmatum | The total coverage of grassland was about 20%. The mucus mound covered about 80%, the density of rat holes was 2/m2, and there were many pebbles on the surface. The rats were submerged and their size was 1–5 cm in length. Grazing yak mainly occupied the plot. There were almost no plants on the surface in winter. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, K.; Gao, X.; Qian, D.; Zhao, L.; Li, Q.; Dai, L. Relationship between Biomass and Biodiversity of Degraded Grassland in the Sanjiangyuan Region of Qinghai–Tibet Plateau. Diversity 2022, 14, 1002. https://doi.org/10.3390/d14111002
Shu K, Gao X, Qian D, Zhao L, Li Q, Dai L. Relationship between Biomass and Biodiversity of Degraded Grassland in the Sanjiangyuan Region of Qinghai–Tibet Plateau. Diversity. 2022; 14(11):1002. https://doi.org/10.3390/d14111002
Chicago/Turabian StyleShu, Kai, Xue Gao, Dawen Qian, Lei Zhao, Qian Li, and Licong Dai. 2022. "Relationship between Biomass and Biodiversity of Degraded Grassland in the Sanjiangyuan Region of Qinghai–Tibet Plateau" Diversity 14, no. 11: 1002. https://doi.org/10.3390/d14111002